
First Short-Term Study of the Relationship between Native and Invasive Earthworms in the Zone of Soil Freezing in Western Siberia—Experiments in Mesocosms
 , , , and
, , , and
Abstract
:1. Introduction
- What is the reproductive potential of the invasive species A. caliginosa and L. rubellus compared to the native E. nordenskioldi?
- Do species interactions affect each other’s reproductive performance, survival, and vertical distribution?
- Are the quantity and quality of litter characteristics of natural habitats a limitation for invasive species?
- Can invasive species tolerate the freezing soils in the absence of additional heat sources in the form of accumulations of organic matter?
2. Materials and Methods
2.1. Experimental Site
2.2. Preparation of Soils, Litter, and Worms
2.3. Mesocosms
2.4. Experiment Design during the Vegetation Period
2.5. Experiment Design during the Winter Period
2.6. Statistical Analysis
3. Results
3.1. Vegetation Experiment
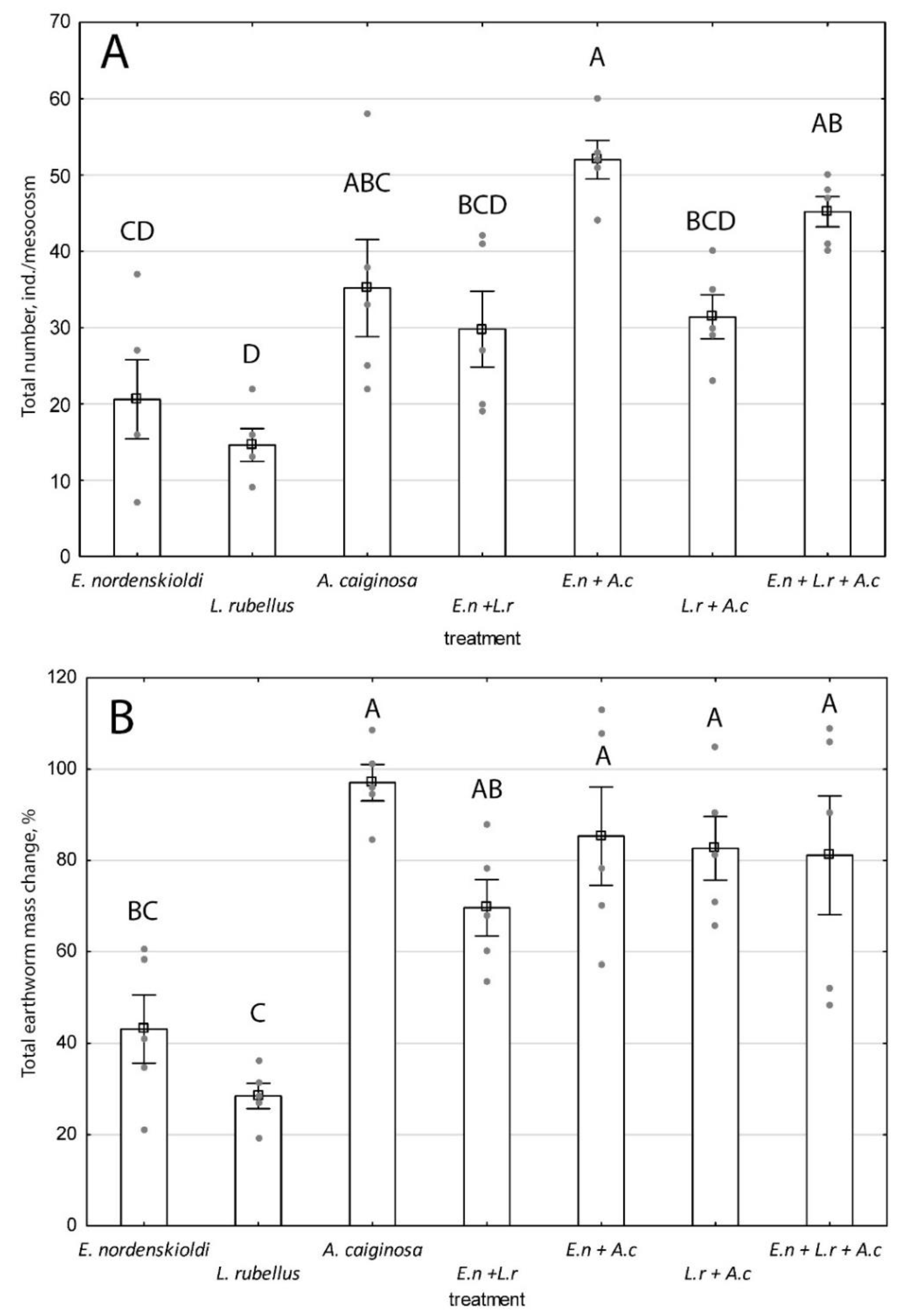
3.1.1. Survival and Reproductive Potential of Species in a Monospecies Experiment
3.1.2. Interaction of Species
3.1.3. Vertical Distribution
3.1.4. Litter Consumption
3.2. Winter Experiment
4. Discussion
4.1. Survival and Reproductive Potential of Native and Invasive Species
4.2. Interaction of Species
4.3. Vertical Distribution
4.4. Litter Consumption
4.5. Soil Freezing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roura-Pascual, N.; Leung, B.; Rabitsch, W.; Rutting, L.; Vervoort, J.; Bacher, S.; Dullinger, S.; Erb, K.-H.; Jeschke, J.M.; Katsanevakis, S.; et al. Alternative Futures for Global Biological Invasions. Sustain. Sci. 2021, 16, 1637–1650. [Google Scholar] [CrossRef]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussaard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Peres, G.; Tondoh, J.E.; et al. A Review of Earthworm Impact on Soil Function and Ecosystem Services: Earthworm Impact on Ecosystem Services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Crooks, J.A. Characterizing Ecosystem-Level Consequences of Biological Invasions: The Role of Ecosystem Engineers. Oikos 2002, 97, 153–166. [Google Scholar] [CrossRef]
- Vilà, M.; Hulme, P.E. (Eds.) Impact of Biological Invasions on Ecosystem Services; Springer International Publishing: Cham, Switzerland, 2017; ISBN 9783319451190. [Google Scholar]
- Blakemore, R.J. Cosmopolitan Earthworms—A Global and Historical Perspective. In Annelids in Modern Biology; Shain, D.H., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 257–283. ISBN 9780470455203. [Google Scholar]
- Hendrix, P.F.; Callaham, M.A.; Drake, J.M.; Huang, C.-Y.; James, S.W.; Snyder, B.A.; Zhang, W. Pandora’s Box Contained Bait: The Global Problem of Introduced Earthworms. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 593–613. [Google Scholar] [CrossRef]
- Stebbings, J.H. Endemic-Exotic Earthworm Competition in the American Midwest. Nature 1962, 196, 905–906. [Google Scholar] [CrossRef]
- Winsome, T.; Epstein, L.; Hendrix, P.F.; Horwath, W.R. Competitive Interactions between Native and Exotic Earthworm Species as Influenced by Habitat Quality in a California Grassland. Appl. Soil Ecol. 2006, 32, 38–53. [Google Scholar] [CrossRef]
- Loss, S.R.; Paudel, S.; Laughlin, C.M.; Zou, C. Local-Scale Correlates of Native and Non-Native Earthworm Distributions in Juniper-Encroached Tallgrass Prairie. Biol. Invasions 2017, 19, 1621–1635. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil Fauna Responses to Natural Disturbances, Invasive Species, and Global Climate Change: Current State of the Science and a Call to Action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Uvarov, A.V. Inter- and Intraspecific Interactions in Lumbricid Earthworms: Their Role for Earthworm Performance and Ecosystem Functioning. Pedobiologia 2009, 53, 1–27. [Google Scholar] [CrossRef]
- Uvarov, A.V.; Ilieva-Makulec, K.; Karaban, K.; Yakovenko, N.S.; Uchmański, J. Effects of Intra- and Interspecific Interactions in Earthworm Assemblages: A Comparative Study. Biol. Bull. Russ. Acad. Sci. 2019, 46, 475–482. [Google Scholar] [CrossRef]
- Kniazev, S.Y.; Kislyi, A.A.; Bogomolova, I.N.; Golovanova, E.V. Territorial Heterogeneity of the Earthworm Population (Opisthopora, Lumbricidae) of Omsk Oblast and Environmental Factors: A Quantitative Assessment of the Relationship. Contemp. Probl. Ecol. 2022, 15, 484–493. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Golovanova, E.V.; Peltek, S.E. Different Dispersal Histories of Lineages of the Earthworm Aporrectodea caliginosa (Lumbricidae, Annelida) in the Palearctic. Biol. Invasions 2016, 18, 751–761. [Google Scholar] [CrossRef]
- Golovanova, E.V.; Kniazev, S.Y.; Babiy, K.A.; Tsvirko, E.I. Distribution of Alien Species of Earthworms Aporrectodea caliginosa in Natural Habitats of the Omsk Region. In Knowledge and Activity: From the Past to the Present; Materials of the II Russian Interdisciplinary Scientific Conference; Federal State Budgetary Educational Institution of Higher Professional Education, Omsk State Pedagogical University: Omsk, Russia, 2020; pp. 299–302. [Google Scholar]
- Ermolov, S.A. Earthworm Communities (Oligochaeta, Lumbricidae) of Pine Forests and Small Foliage Forests in the Forest-Steppe Ob Region. FSI 2020, 3, 1–24. [Google Scholar] [CrossRef]
- Golovanova, E.V. Earthworms of the Omsk region. Proc. Tomsk. State Univ. 2010, 275, 111–114. [Google Scholar]
- Striganova, B.R.; Poryadina, N.M. Soil Animal Population in Boreal Forests of West-Siberian Plain; KMK: Moscow, Russia, 2005; ISBN 9785873172382. [Google Scholar]
- Vsevolodova-Perel, T.S. The Earthworm of the Fauna of Russia: Cadaster and Key; Nauka: Moscow, Russia, 1997. [Google Scholar]
- Blakemore, R.J. Checklist of USSR/Russian Federation Taxa Updated from Perel (1979, 1997) in a Series of Searchable Texts on Earthworm Biodiversity, Ecology and Systematics from Various Regions of the World, 3rd ed.; 2008. Available online: http://www.annelida.net/earthworm/ (accessed on 17 January 2023).
- Shekhovtsov, S.V.; Golovanova, E.V.; Peltek, S.E. Cryptic Diversity within the Nordenskiold’s Earthworm, Eisenia nordenskioldi subsp. nordenskioldi (Lumbricidae, Annelida). Eur. J. Soil Biol. 2013, 58, 13–18. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Golovanova, E.V.; Ershov, N.I.; Poluboyarova, T.V.; Berman, D.I.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Phylogeny of the Eisenia nordenskioldi Complex Based on Mitochondrial Genomes. Eur. J. Soil Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Shipova, A.A.; Poluboyarova, T.V.; Vasiliev, G.V.; Golovanova, E.V.; Geraskina, A.P.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Species Delimitation of the Eisenia nordenskioldi Complex (Oligochaeta, Lumbricidae) Using Transcriptomic Data. Front. Genet. 2020, 11, 598196. [Google Scholar] [CrossRef]
- Perel, T.S. Distribution and Patterns of Distribution of Earthworms in the Fauna of the USSR (with Key Tables); Nauka: Moscow, Russia, 1979. [Google Scholar]
- Felten, D.; Emmerling, C. Earthworm Burrowing Behaviour in 2D Terraria with Single- and Multi-Species Assemblages. Biol. Fertil Soils 2009, 45, 789–797. [Google Scholar] [CrossRef]
- Neilson, R.; Boag, B. Feeding Preferences of Some Earthworm Species Common to Upland Pastures in Scotland. Pedobiologia 2003, 47, 1–8. [Google Scholar] [CrossRef]
- Potapov, A.M.; Tiunov, A.V.; Scheu, S.; Larsen, T.; Pollierer, M.M. Combining Bulk and Amino Acid Stable Isotope Analyses to Quantify Trophic Level and Basal Resources of Detritivores: A Case Study on Earthworms. Oecologia 2019, 189, 447–460. [Google Scholar] [CrossRef]
- Bottinelli, N.; Hedde, M.; Jouquet, P.; Capowiez, Y. An Explicit Definition of Earthworm Ecological Categories—Marcel Bouché’s Triangle Revisited. Geoderma 2020, 372, 114361. [Google Scholar] [CrossRef]
- Hsu, G.-C.; Szlavecz, K.; Csuzdi, C.; Bernard, M.; Chang, C.-H. Ecological Groups and Isotopic Niches of Earthworms. Appl. Soil Ecol. 2023, 181, 104655. [Google Scholar] [CrossRef]
- Capowiez, Y.; Gilbert, F.; Vallat, A.; Poggiale, J.-C.; Bonzom, J.-M. Depth Distribution of Soil Organic Matter and Burrowing Activity of Earthworms—Mesocosm Study Using X-ray Tomography and Luminophores. Biol. Fertil. Soils 2021, 57, 337–346. [Google Scholar] [CrossRef]
- Klok, C. Effects of Earthworm Density on Growth, Development, and Reproduction in Lumbricus rubellus (Hoffm.) and Possible Consequences for the Intrinsic Rate of Population Increase. Soil Biol. Biochem. 2007, 39, 2401–2407. [Google Scholar] [CrossRef]
- Uvarov, A.V. Density-Dependent Responses in Some Common Lumbricid Species. Pedobiologia 2017, 61, 1–8. [Google Scholar] [CrossRef]
- Lowe, C.; Butt, K. Growth of Hatchling Earthworms in the Presence of Adults: Interactions in Laboratory Culture. Biol. Fertil. Soils 2002, 35, 204–209. [Google Scholar] [CrossRef]
- Lowe, C.N.; Butt, K.R. Influence of Food Particle Size on Inter- and Intra-Specific Interactions of Allolobophora chlorotica (Savigny) and Lumbricus terrestris. Pedobiologia 2003, 47, 574–577. [Google Scholar] [CrossRef]
- Karaban, K.; Uvarov, A.V. Non-Trophic Effects of Earthworms on Enchytraeids: An Experimental Investigation. Soil Biol. Biochem. 2014, 73, 84–92. [Google Scholar] [CrossRef]
- Uvarov, A.V.; Karaban, K. Do Alterations in Mesofauna Community Affect Earthworms? Oecologia 2015, 179, 877–887. [Google Scholar] [CrossRef]
- Räty, M.; Huhta, V. Earthworms and PH Affect Communities of Nematodes and Enchytraeids in Forest Soil. Biol. Fertil. Soils 2003, 38, 52–58. [Google Scholar] [CrossRef]
- Aira, M.; Monroy, F.; Domínguez, J. Effects of Two Species of Earthworms (Allolobophora Spp.) on Soil Systems: A Microfaunal and Biochemical Analysis. Pedobiologia 2003, 47, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Clause, J.; Barot, S.; Richard, B.; Decaëns, T.; Forey, E. The Interactions between Soil Type and Earthworm Species Determine the Properties of Earthworm Casts. Appl. Soil Ecol. 2014, 83, 149–158. [Google Scholar] [CrossRef]
- Babiy, K.A.; Kniazev, S.Y.; Golovanova, E.V.; Solomatin, D.V.; Bel’skaya, L.V. What Determines Ion Content of Lumbricid Casts: Soil Type, Species, or Ecological Group? Pol. J. Ecol. 2021, 69, 96–110. [Google Scholar] [CrossRef]
- Bezel, V.S. Ecological Toxicology: Population and Biocenotic Aspects; Goshchitsky: Ekaterinburg, Russia, 2006; ISBN 9785988290070. [Google Scholar]
- Hoeffner, K.; Monard, C.; Cluzeau, D.; Santonja, M. Response of Temperate Anecic Earthworm Individual Biomass to Species Interactions. Appl. Soil Ecol. 2019, 144, 8–11. [Google Scholar] [CrossRef]
- Lowe, C.N.; Butt, K.R. Interspecific interactions between earthworms: A laboratory-based investigation. Pedobiologia 1999, 43, 808–817. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-0033399117&partnerID=40&md5=4d3fe3d64237bb211f6e1d7839183862 (accessed on 1 December 2022).
- Butt, K.R. Interactions between Selected Earthworm Species: A Preliminary, Laboratory-Based Study. Appl. Soil Ecol. 1998, 9, 75–79. [Google Scholar] [CrossRef]
- Suárez, E.R.; Fahey, T.J.; Yavitt, J.B.; Groffman, P.M.; Bohlen, P.J. Patterns of Litter Disappearance in a Northern Hardwood Forest Invaded by Exotic Earthworms. Ecol. Appl. 2006, 16, 154–165. [Google Scholar] [CrossRef]
- Shipitalo, M.; Protz, R.; Tomlin, A. Effect of Diet on the Feeding and Casting Activity of Lumbricus terrestris and L. rubellus in Laboratory Culture. Soil Biol. Biochem. 1988, 20, 233–237. [Google Scholar] [CrossRef]
- Meshcheryakova, E.N.; Berman, D.I. Resistance to negative temperatures and geographical distribution of earthworms (Oligochaeta, Lumbricidae, Moniligastridae). Zool. J. 2014, 93, 53–64. [Google Scholar] [CrossRef]
- Golovanova, E.V. Alien species of earthworms in Western Siberia. In Ecology and Evolution: New Horizons: Materials of Intern. Symposium Dedicated 100th Anniversary of Academician S.S. Schwartz; Ushakova, K.I., Veselkin, D.V., Vasiliev, A.G., Eds.; Humanities University: Ekaterinburg, Russia, 2019; pp. 494–495. [Google Scholar]
- Makarova, O.L.; Kolesnikova, A.A. Earthworms (Oligochaeta, Lumbricidae) in the Tundra of Eastern Europe. Biol. Bull. Russ. Acad. Sci. 2019, 46, 438–449. [Google Scholar] [CrossRef]
- Kolesnikova, A.; Dolgin, M.; Akulova, L. Earthworms (Oligochaeta, Lumdricidae) in the Komi Republic. Proc. KarRC RAS 2022, 1–18. [Google Scholar] [CrossRef]
- Sherstyukov, A.B. Array of daily data on the temperature of soils at depths up to 320 cm according to the meteorological station of the Russian Federation. Proc. All-Russ. Res. Inst. Hydrometeorol. Inf.-World Data Cent. 2012, 176, 233–256. [Google Scholar]
- Nuutinen, V.; Butt, K.R. Worms from the Cold: Lumbricid Life Stages in Boreal Clay during Frost. Soil Biol. Biochem. 2009, 41, 1580–1582. [Google Scholar] [CrossRef]
- Blakemore, R. Earthworms Newly from Mongolia (Oligochaeta, Lumbricidae, Eisenia). ZooKeys 2013, 285, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil layer | October | November | December | January | February | Mart | April | |
|---|---|---|---|---|---|---|---|---|
| 0–5 cm | max t | 3.4 | −0.1 | 0.0 | −0.9 | −1.0 | 0.2 | 10.7 |
| min t | −1.8 | −5.6 | −2.9 | −2.5 | −2.5 | −1.7 | 0.1 | |
| 50 cm | max t | 5.8 | 3.2 | 0.8 | 0.2 | −0.1 | 0.1 | 7.3 |
| min t | 3.2 | 0.8 | 0.2 | −0.1 | −0.4 | −0.3 | 0.1 | |
| 100 cm | max t | 8.3 | 5.4 | 3.0 | 2.0 | 1.1 | 1.1 | 4.8 |
| min t | 5.4 | 3.0 | 2.0 | 1.5 | 1.5 | 0.9 | 0.8 | |
| 150 cm | max t | 8.6 | 6.2 | 4.6 | 3.4 | 2.7 | 2.2 | 3.9 |
| min t | 2.0 | 4.6 | 3.4 | 2.7 | 2.2 | 2.0 | 1.7 | |
| 200 cm | max t | 9.5 | 7.8 | 5.6 | 4.1 | 3.4 | 2.8 | 3.5 |
| min t | 7.8 | 5.6 | 4.2 | 3.4 | 2.8 | 2.5 | 2.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golovanova, E.V.; Kniazev, S.Y.; Karaban, K.; Babiy, K.A.; Shekhovtsov, S.V. First Short-Term Study of the Relationship between Native and Invasive Earthworms in the Zone of Soil Freezing in Western Siberia—Experiments in Mesocosms. Diversity 2023, 15, 248. https://doi.org/10.3390/d15020248
Golovanova EV, Kniazev SY, Karaban K, Babiy KA, Shekhovtsov SV. First Short-Term Study of the Relationship between Native and Invasive Earthworms in the Zone of Soil Freezing in Western Siberia—Experiments in Mesocosms. Diversity. 2023; 15(2):248. https://doi.org/10.3390/d15020248
Chicago/Turabian StyleGolovanova, Elena V., Stanislav Yu. Kniazev, Kamil Karaban, Kirill A. Babiy, and Sergei V. Shekhovtsov. 2023. "First Short-Term Study of the Relationship between Native and Invasive Earthworms in the Zone of Soil Freezing in Western Siberia—Experiments in Mesocosms" Diversity 15, no. 2: 248. https://doi.org/10.3390/d15020248