





Exploring Texture Diversity of Beech-Spruce-Fir Stands through Development Phase Analysis in the Frakto Virgin Forest of Greece


Abstract
:1. Introduction
- (a)
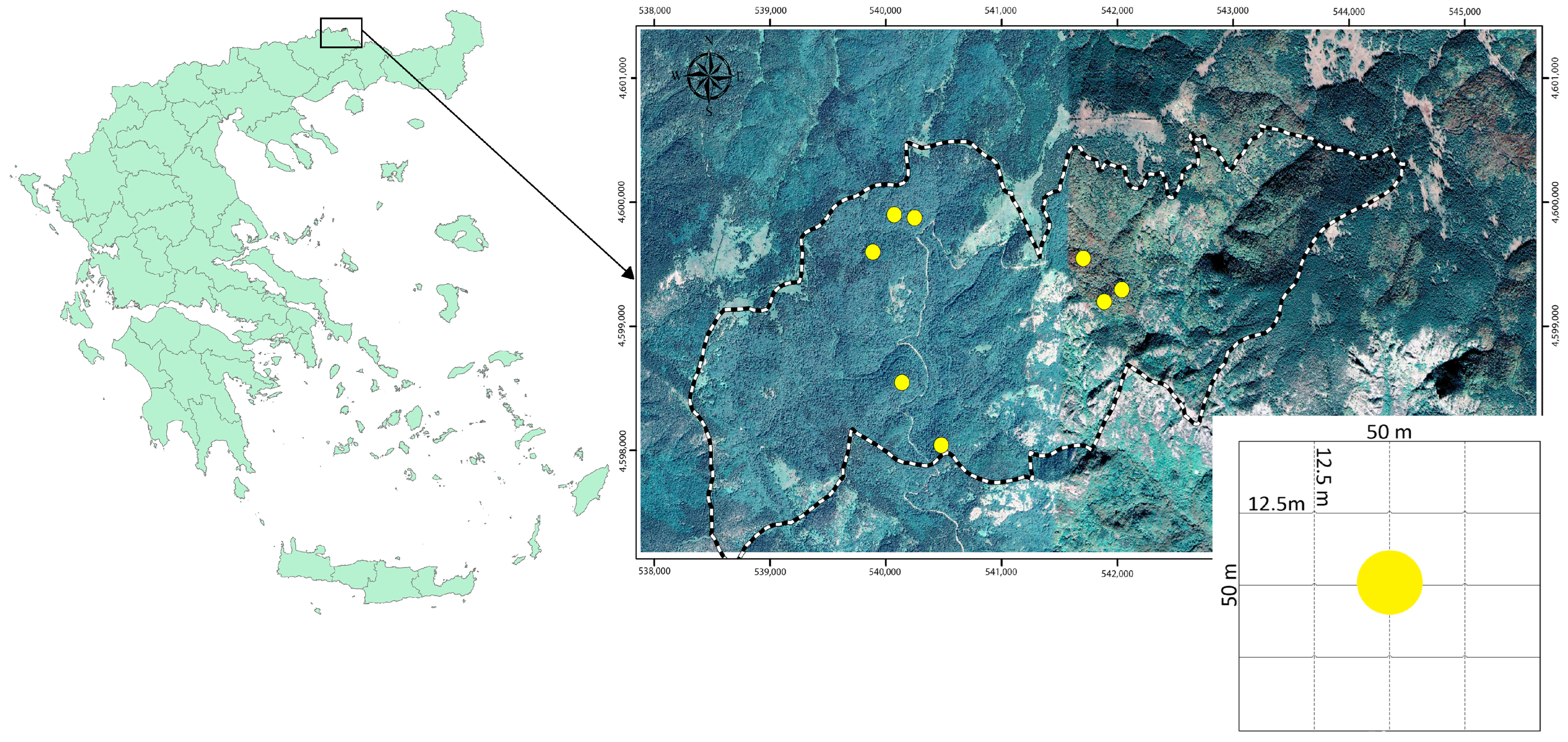
- To document the texture diversity of the unique old-growth virgin Frakto forest in northeastern Greece through identification, frequency spatial distribution, and arrangement of the forest development phases;
- (b)
- To investigate the specific stand structural attributes that may differ significantly among the three major developmental stages.
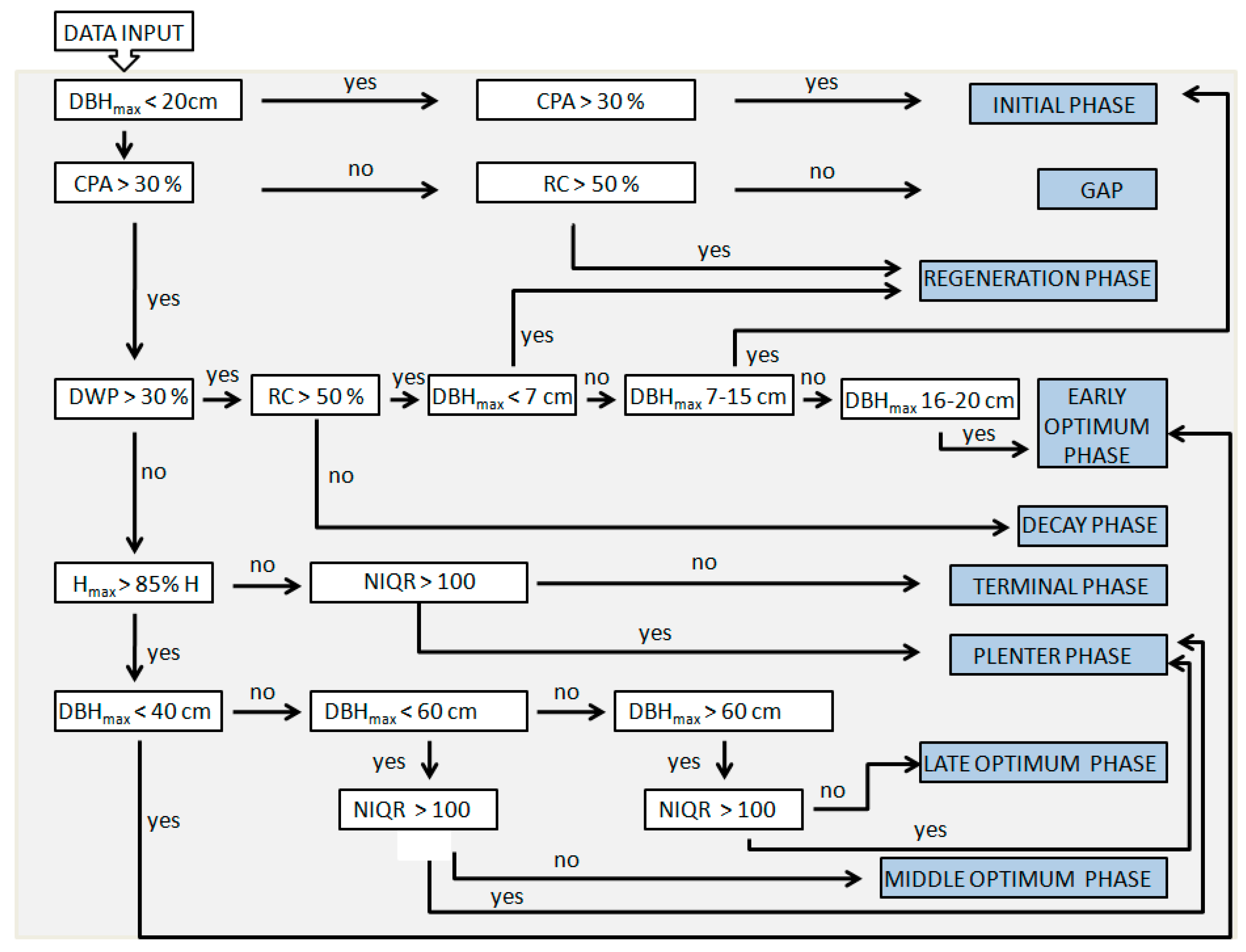
2. Materials and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glatthorn, J.; Feldmann, E.; Tabaku, V.; Leuschner, C.; Meyer, P. Classifying development stages of primeval European beech forests: Is clustering a useful tool? BMC Ecol. 2018, 18, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, P.; Schmidt, M. Aspekte der Biodiversität von Buchenwäldern—Konsequenzen für eine naturnahe Bewirtschaftung. In Ergebnisse Angewandter Forschung Zur Buche. Beiträge Aus Der Nordwestdeutschen Forstlichen Versuchsanstalt; NW-FVA: Göttingen, Germany, 2008; Volume 3, pp. 159–192. [Google Scholar]
- Král, K.; Vrška, T.; Hort, L.; Adam, D.; Šamonil, P. Developmental phases in a temperate natural spruce-fir-beech forest: Determination by a supervised classification method. Eur. J. For. Res. 2009, 129, 339–351. [Google Scholar] [CrossRef]
- Winter, S. Ermittlung Von Struktur-Indikatoren Zur Abschätzung Des Einflusses Forstlicher Bewirtschaftung Auf Die Biozönosen Von Tiefland-Buchenwäldern. Ph.D. Dissertation, TU Dresden, Dresden, Germany, 2005. [Google Scholar]
- Giurgiu, V. (Ed.) Pădurile Virgine Si Cvasivirgine Ale României [Virgin and Quasi-Virgin Romanian Forests]; Editura AcademieiRomâne: București, Romania, 2013; p. 390. [Google Scholar]
- Boncina, A. Comparison of structure and biodiversity in the Rajhenav virgin forest remnant and managed forest in the Dinaric region of Slovenia. Glob. Ecol. Biogeogr. 2000, 9, 201–211. [Google Scholar] [CrossRef]
- Grgic, T.; Kos, I. Centipede diversity in patches of different development phases in an unevenly-aged beech forest stand in Slovenia. Afr. Invertebr. 2003, 44, 237–252. [Google Scholar]
- Král, K.; McMahon, S.M.; Janík, D.; Adam, D.; Vrška, T. Patch mosaic of developmental stages in central European natural forests along vegetation gradient. For. Ecol. Manag. 2014, 330, 17–28. [Google Scholar] [CrossRef]
- Mokrzycki, T. Effect of pine forest stand age on bark beetles communities (Coleoptera: Scolytidae) in Puszcza Białowieska. Fragm. Faun. 1995, 38, 411–417. [Google Scholar] [CrossRef]
- Niemelä, J.; Haila, Y.; Punttila, P. The importance of small-scale heterogeneity in boreal forests: Variation in diversity in forest-floor invertebrates across the succession gradient. Ecography 1996, 19, 352–368. [Google Scholar] [CrossRef]
- Begehold, H.; Rzanny, M.; Flade, M. Forest development phases as an integrating tool to describe habitat preferences of breeding birds in lowland beech forests. J. Ornithol. 2015, 156, 19–29. [Google Scholar] [CrossRef]
- Sattout, E.; Caligari, P.D. Forest biodiversity assessment in relic ecosystem: Monitoring and management practice implications. Diversity 2011, 3, 531–546. [Google Scholar] [CrossRef]
- Watt, A.S. Pattern and process in the plant community. J. Ecol. 1947, 35, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Rehush, N.; Waser, L. Assessing the structure of primeval and managed beech forests in the Ukrainian Carpathians using remote sensing. Can. J. For. Res. 2017, 47, 63–72. [Google Scholar] [CrossRef]
- Parobeková, Z.; Pittner, J.; Kucbel, S.; Saniga, M.; Filípek, M.; Sedmáková, D.; Vencurik, J.; Jaloviar, P. Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians. Forests 2018, 9, 379. [Google Scholar] [CrossRef] [Green Version]
- Drößer, L.; Von Lüpke, B. Canopy gaps in two virgin beech forest reserves in Slovakia. J. For. Sci. 2005, 51, 446–457. [Google Scholar] [CrossRef] [Green Version]
- Kenderes, K.; Mihók, B.; Standovár, T. Thirty years of gap dynamics in a central european beech forest reserve. For. Int. J. For. Res. 2008, 81, 111–123. [Google Scholar] [CrossRef]
- Zenner, E.K.; Peck, J.E.; Hobi, M.L.; Commarmot, B. The dynamics of structure across scale in a primaeval European beech stand. For. Int. J. For. Res. 2014, 88, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Paluch, J.G. The spatial pattern of a natural European beech (Fagus sylvatica L.)–silver fir (Abies alba Mill.) forest: A patch-mosaic perspective. For. Ecol. Manag. 2007, 253, 161–170. [Google Scholar] [CrossRef]
- Leibundgut, H. Über Zweck und Methodik der Struktur- und Zuwachsanalyse von Urwäldern. Schweiz. Ztg. Für Forstwes. 1959, 110, 111–124. [Google Scholar]
- Leibundgut, H. Über Dynamik europäischer Urwälder. Allg. Forstztg. 1978, 33/24, 686–690. [Google Scholar]
- Leibundgut, H. Europäische Urwälder der Bergstufe, dargestellt für Forstleute, Naturwissenschaftler und Freunde des Waldes; Haupt: Bern, Germany; Stuttgart, Germany, 1982; 306p. [Google Scholar]
- Oldeman, R.A.A. Forests: Elements of Silvology; Springer: Berlin/Heidelberg, Germany, 1990. [Google Scholar] [CrossRef] [Green Version]
- Mayer, H.; Ott, E. Gebirgswaldbau—Schutzwaldpflege, 2nd ed.; Gustav Fischer: Stuttgart, Germany; New York, NY, USA, 1991; 587p. [Google Scholar]
- Körpel, S. Ergebnisse der Urwaldforschung für die Waldwirtschaft im Buchen-Ökosysteme. Allg. Forst-Und Jagdztg. 1992, 21, 1148–1152. [Google Scholar]
- Körpel, S. Vorkommen, Charakteristik und Folge der Enwicklungsstadien-phasen in den europäischen Urwäldern. In Symposium Über Die Urwälder; Saniga, M., Körpel, S., Eds.; Forstliche Fakultät der technischen Universität Zvolen: Zvolen, Slovakia, 1993; pp. 3–10. [Google Scholar]
- Körpel, S. Die Urwälder Der Westkarpaten; Gustav Fischer: Stuttgart, Germany; Jena, Germany; New York, NY, USA, 1995; 310p. [Google Scholar]
- Meyer, P. Bestimmung der Waldentwicklungsphasen und der Texturdiversität in Naturwäldern. Allg. Forst-Und Jagdztg. 1999, 170, 203–211. [Google Scholar]
- Tabaku, V. Struktur von Buchen-Urwäldern in Albanien im Vergleich mit deutschen Buchen-Naturwaldreservaten und-Wirtschaftwäldern. Ph.D. Dissertation, Univ. Göttingen, Göttingen, Germany, 1999. [Google Scholar]
- Commarmot, B.; Bachofen, H.; Bundziak, Y.; Bürgi, A.; Ramp, B.; Shparyk, Y.; Sukhariuk, D.; Viter, R.; Zingg, A. Structure of virgin and managed beech forests in Uholka (Ukraine) and Sihlwald (Switzerland): A comparative study. For. Snow Lands. Res. 2005, 79, 45–56. [Google Scholar]
- Drößler, L.; Meyer, P. Waldentwicklungsphasen in zwei Buchen-Urwaldreservaten in der Slowakei. Forstarchiv 2006, 77, 155–161. [Google Scholar]
- Winter, S.; Brambach, F. Determination of a Common Forest Life Cycle Assessment Method for Biodiversity Evaluation. For. Ecol. Manag. 2011, 262, 2120–2132. [Google Scholar] [CrossRef]
- Král, K.; Daněk, P.; Janík, D.; Krůček, M.; Vrška, T. How cyclical and predictable are Central European temperate forest dynamics in terms of development phases? J. Veg. Sci. 2018, 29, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Government Gazette 121/issue D’/21-02-1980. Available online: https://www.et.gr/ (accessed on 1 September 2022).
- Papadopoulou, D. Dynamics of Structure and Natural Regeneration in the Undisturbed Forest Ecosystem of Virgin Forest Frakto. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2017. [Google Scholar]
- Kazana, V.; Kazaklis, A.; Raptis, D.; Stamatiou, C. A combined multi-criteria approach to assess forest management sustainability: An application to the forests of Eastern Macedonia & Thrace Region in Greece. Ann. Oper. Res. 2020, 294, 321–343. [Google Scholar] [CrossRef]
- Kechagioglou, S.; Papadopoulou, D.; Tsitsoni, T. Determining structure and volume of the European beech (Fagus Sylvatica L.) deadwood in managed stands in the Rodopi Mountain Range National Park, Greece. Folia Oecologica 2022, 49, 122–129. [Google Scholar] [CrossRef]
- Mansourian, S.; Rossi, M.; Vallauri, D. Ancient Forests in the Northern Mediterranean: Neglected High Conservation Value Areas; WWF: Marseille, France, 2013; 80p. [Google Scholar]
- Eleftheriadou, E.; Raus, T. The vascular flora of the nature reserve Frakto Virgin Forest of Nomos Dramas (E Makedonia, Greece). Willdenowia 1996, 25, 455–485. [Google Scholar]
- Tsiftsis, S.; Karagiannakidou, V.; Tsiripidis, I. The orchid flora of east Macedonia (NE Greece). J. Eur. Orch. 2007, 39, 489–526. [Google Scholar]
- Blockeel, T.L. Bryophytes of Frakto Forest (Rhodope Mountains, Greece), Including Syzygiella autumnalis and the Genus Ulota New to Greece. Cryptogam. Bryol. 2018, 39, 129–146. [Google Scholar] [CrossRef]
- Dafis, S. The Forests of Greece; Goulandris Natural History Museum: Athens, Greece, 2010; 192p. (In Greek) [Google Scholar]
- Kershaw, J.A.; Ducey, M.J.; Beers, T.W.; Husch, B. Forest Mensuration, 5th ed.; Wiley: New York, NY, USA, 2016; 632p. [Google Scholar]
- Raptis, D.I.; Kazana, V.; Kechagioglou, S.; Kazaklis, A.; Stamatiou, C.; Papadopoulou, D.; Tsitsoni, T. Nonlinear Quantile Mixed-Effects Models for Prediction of the Maximum Crown Width of Fagus sylvatica L., Pinus nigra Arn. and Pinus brutia Ten. Forests 2022, 13, 499. [Google Scholar] [CrossRef]
- Lemon, P. A Spherical Densiometer For Estimating Forest Overstory Density. For. Sci. 1956, 2, 314–320. [Google Scholar]
- Raptis, D.I.; Kazana, V.; Kazaklis, A.; Stamatiou, C. Development and testing of volume models for Pinus nigra Arn., Fagus sylvatica L., and Quercus pubescen Willd. South. For. A J. For. Sci. 2020, 82, 331–341. [Google Scholar] [CrossRef]
- Merganičová, K.; Merganič, J. Coarse woody debris carbon stocks in natural spruce forests of Babia hora. J. For. Sci. 2010, 56, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Mataji, A.; Sagheb-Talebi, K.; Eshaghi-Rad, J. Deadwood assessment in different developmental stages of beech (Fagus orientalis Lipsky) stands in Caspian forest ecosystems. Int. J. Environ. Sci. Technol. 2014, 11, 1215–1222. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 10 September 2022).
- Meyer, P.; Tabaku, V.; von Lüpke, Β. Die Struktur albanischer Rotbuchen-Urwälder–Ableitungen für eine naturnahe Buchenwirtschaft: Structural Characteristics of Albanian Beech (Fagus sylvatica L.) Virgin Forests–Deductions for Semi-Natural Forestry. Forstwiss. Cent. 2003, 122, 47–58. [Google Scholar] [CrossRef]
- Leibundgut, H. Europäische Urwälder. Wegweiser zur naturnahen Waldwirtschaft; Haupt: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 1993; 260p. [Google Scholar]
- Anic, I.; Mikac, S. Structure, texture and regeneration of dinaric beech–fir virgin forest of Corkova uvala. Sumarski List 2008, 132, 505–515. [Google Scholar]
- Višnjić, C.; Balić, B.; Vojniković, S.; Mekić, F.; Uzunović, A. Structural characteristics, dynamics and texture development of virgin forest Ravna vala on Bjelašnica. Work. Fac. For. Univ. Sarajevo 2015, 45, 100–112. [Google Scholar] [CrossRef]
- Mayer, H.; Neumann, M. Struktureller und entwicklungsdynamischer Vergleich der Fichten-Tannen-Buchen-Urwälder Rothwald/Niederösterreich und CorkovaUvala/Kroatien. Forstwiss. Cent.bl. 1981, 100, 111–132. [Google Scholar] [CrossRef]
- Drößler, L. Struktur und Dynamik von zwei Buchenurwäldern in der Slowakei. Ph.D. Dissertation, Georg-August-Universität, Göttingen, Germany, 2006. [Google Scholar]
- Bobiec, A. Living stands and dead wood in the Białowieża forest: Suggestions for restoration management. For. Ecol. Manag. 2002, 165, 125–140. [Google Scholar] [CrossRef]
- Larrieu, L.; Cabanettes, A.; Gonin, P.; Lachat, T.; Paillet, Y.; Winter, S.; Bouget, C.; Deconchat, M. Deadwood and tree microhabitat dynamics in unharvested temperate mountain mixed forests: A life-cycle approach to biodiversity monitoring. For. Ecol. Manag. 2014, 334, 163–173. [Google Scholar] [CrossRef]
- Öder, V.; Petritan, A.M.; Schellenberg, J.; Bergmeier, E.; Walentowski, H. Patterns and drivers of deadwood quantity and variation in mid-latitude deciduous forests. For. Ecol. Manag. 2021, 487, 118977. [Google Scholar] [CrossRef]
- Bujoczek, L.; Szewczyk, J.; Bujoczek, M. Deadwood volume in strictly protected, natural, and primeval forests in Poland. Eur. J. For. Res. 2018, 137, 401–418. [Google Scholar] [CrossRef]
- Schuck, A.; Meyer, P.; Menke, N.; Lier, M.; Lindner, M. Forest biodiversity indicator: Dead wood—A proposed approach towards operationalising the MCPFE indicator. Monit. Indic. For. Biodivers. Eur. Ideas Oper. 2004, 51, 49–77. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
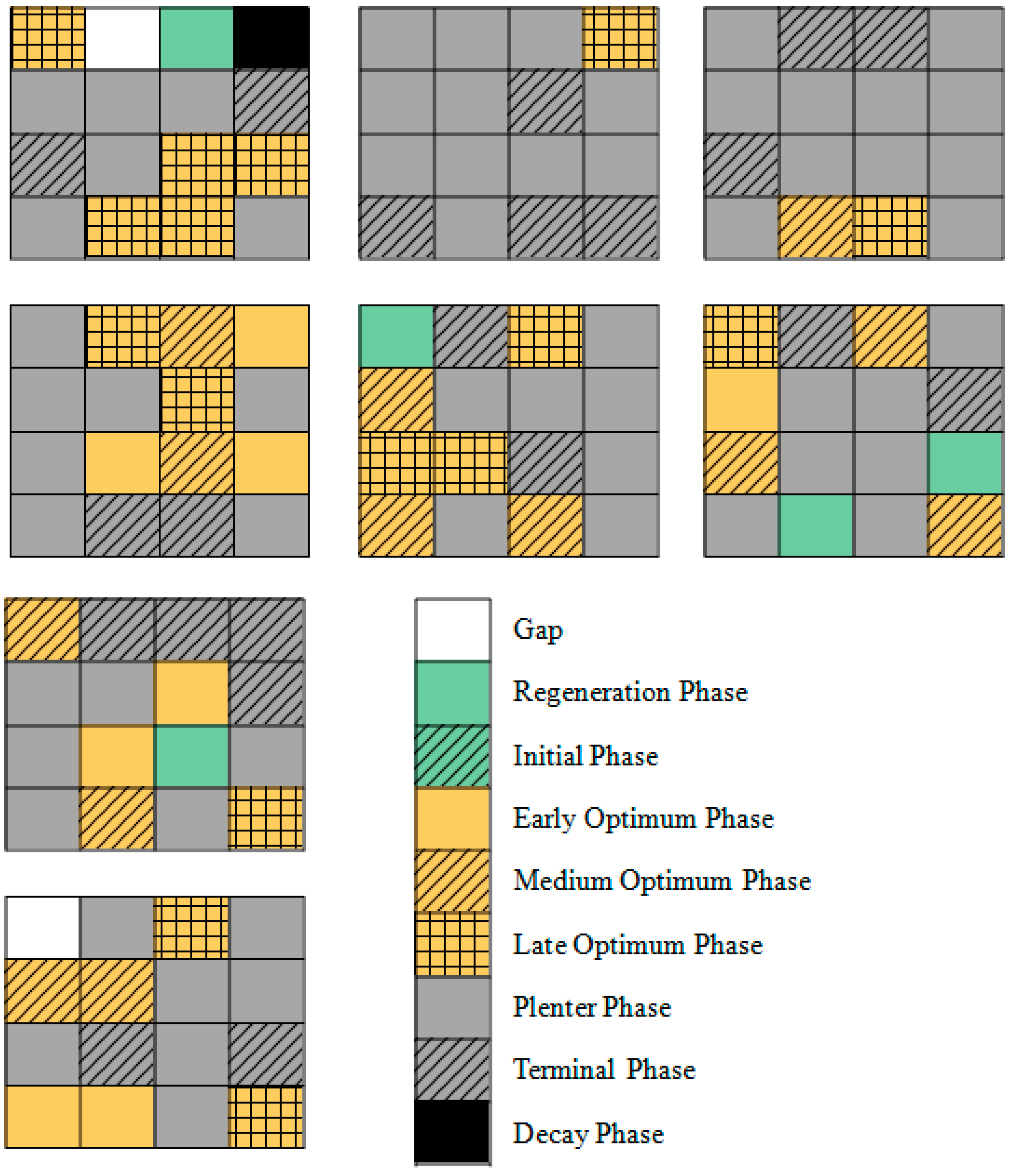
| Development Phases | Plot 1 (%) | Plot 2 (%) | Plot 3 (%) | Plot 4 (%) | Plot 5 (%) | Plot 6 (%) | Plot 7 (%) | Plot 8 (%) | Total Spatial Proportion (%) |
|---|---|---|---|---|---|---|---|---|---|
| Gap | 0 | 6.25 | 6.25 | 0 | 0 | 0 | 0 | 0 | 1.56 |
| Regeneration Phase | 6.25 | 0 | 6.25 | 0 | 6.25 | 0 | 0 | 6.25 | 3.12 |
| Initial Phase | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.00 |
| Early Optimum Phase | 6.25 | 12.5 | 0 | 0 | 0 | 0 | 18.75 | 12.5 | 6.25 |
| Middle Optimum Phase | 18.75 | 12.5 | 0 | 0 | 18.75 | 6.25 | 12.5 | 12.5 | 10.16 |
| Late Optimum Phase | 6.25 | 12.5 | 31.25 | 6.25 | 18.75 | 6.25 | 12.5 | 6.25 | 12.50 |
| Plenter Phase | 43.75 | 43.75 | 37.5 | 68.75 | 43.75 | 68.75 | 43.75 | 37.5 | 48.44 |
| Terminal Phase | 12.5 | 12.5 | 12.5 | 25 | 12.5 | 18.75 | 12.5 | 25 | 16.41 |
| Decay Phase | 6.25 | 0 | 6.25 | 0 | 0 | 0 | 0 | 0 | 1.56 |
| Species | Density (%) | Basal Area | Volume of Living Trees | Deadwood Volume |
|---|---|---|---|---|
| Beech | 84 | 67 | 63 | 26 |
| Silver fir | 10 | 13 | 16 | 69 1 |
| Spruce | 6 | 20 | 21 | |
| Unidentified | - | - | - | 5 |
| Development Phase | Species | Density (N/ha) | Basal Area (m2/ha) | Volume of Living Trees (m3/ha) |
|---|---|---|---|---|
| Gap | Beech | 100 | 100 | 100 |
| Silver fir | - | - | - | |
| Spruce | - | - | - | |
| Regeneration Phase | Beech | 74.1 | 78.1 | 77.8 |
| Silver fir | 6.3 | 1 | 0.4 | |
| Spruce | 19.6 | 21 | 21.7 | |
| Initial Phase | Beech | - | - | - |
| Silver fir | - | - | - | |
| Spruce | - | - | - | |
| Early Optimum Phase | Beech | 89.3 | 96 | 91.1 |
| Silver fir | 9.4 | 2.3 | 0.8 | |
| Spruce | 1.3 | 1.7 | 2.2 | |
| Middle Optimum Phase | Beech | 89.1 | 79.2 | 79.5 |
| Silver fir | 6.4 | 8 | 9.8 | |
| Spruce | 4.5 | 10.6 | 10.6 | |
| Late Optimum Phase | Beech | 84.4 | 75.6 | 73.3 |
| Silver fir | 8.7 | 12.8 | 14.5 | |
| Spruce | 6.9 | 11.6 | 12.3 | |
| Plenter Phase | Beech | 82.5 | 63.6 | 60.8 |
| Silver fir | 11.3 | 14.3 | 16.5 | |
| Spruce | 6.2 | 22.1 | 22.7 | |
| Terminal Phase | Beech | 81.9 | 56.6 | 54.5 |
| Silver fir | 6.3 | 12.2 | 13.5 | |
| Spruce | 11.8 | 31.2 | 32 | |
| Decay Phase | Beech | 46.6 | 64.9 | 68.6 |
| Silver fir | 49.5 | 20.8 | 20.2 | |
| Spruce | 3.8 | 14.3 | 11.1 |
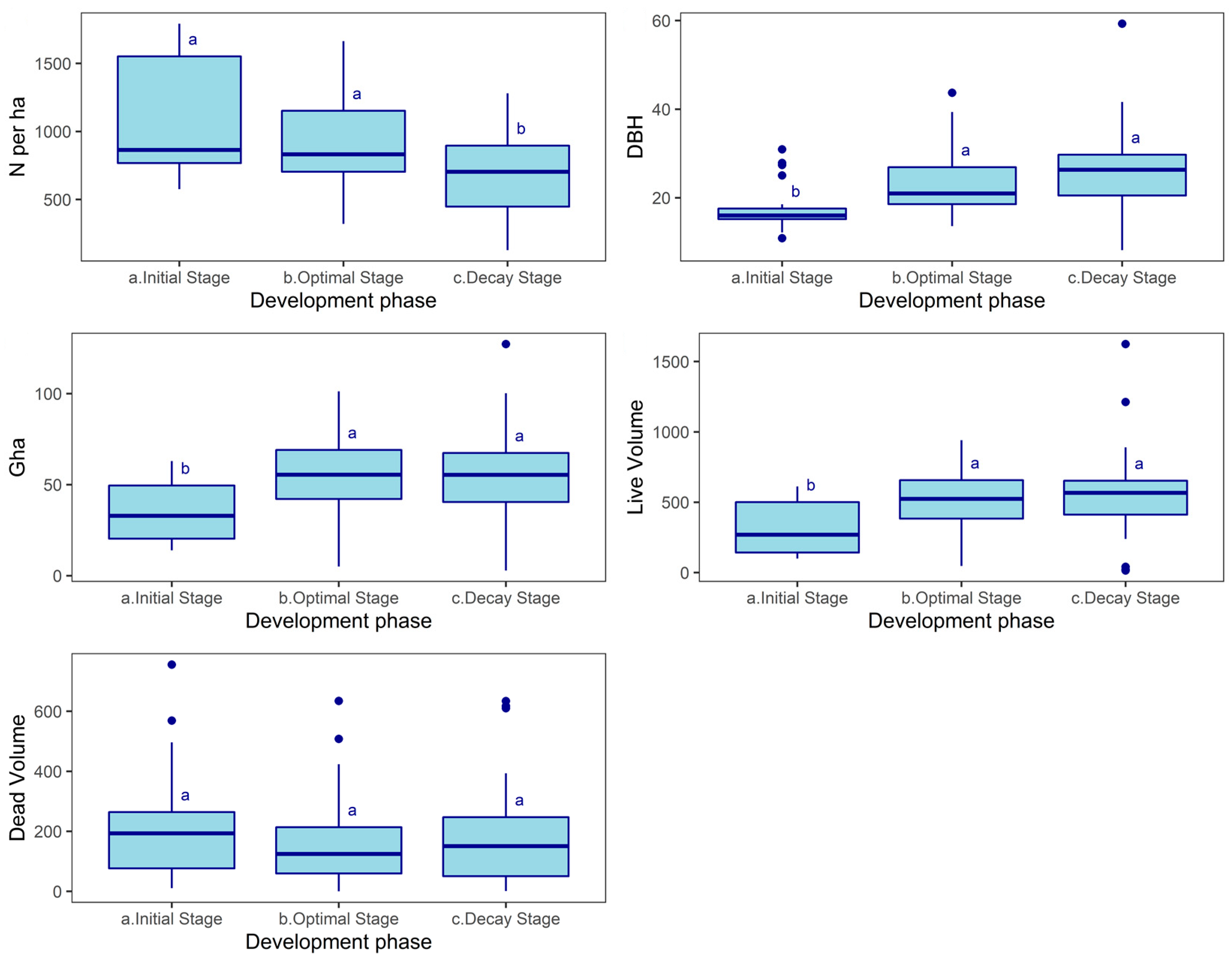
| Development Phase | Number of Subplots | Density (N/ha) | DBH (cm) | Basal Area (m2/ha) | Volume of Living Trees (m3/ha) | Deadwood Volume (m3/ha) |
|---|---|---|---|---|---|---|
| Gap | 2 | 192 ± 91 | 12.30 ± 5.80 | 3.09 ± 0.36 | 18.46 ± 5.17 | 146.24 ± 38.92 |
| Regeneration Phase | 4 | 224 ± 45 | 23.97 ± 6.41 | 14.91 ± 12.31 | 140.46 ± 139.86 | 465.12 ± 215.87 |
| Initial Phase | - | - | - | - | - | - |
| Early Optimum Phase | 8 | 1152 ± 445 | 14.67 ± 3.21 | 25.37 ± 12.44 | 196.69 ± 120.47 | 150.18 ± 189.04 |
| Middle Optimum Phase | 13 | 1014 ± 414 | 19.87 ± 5.73 | 44.55 ± 15.65 | 401.01 ± 157.06 | 255.82 ± 196.87 |
| Late Optimum Phase | 16 | 908 ± 402 | 23.78 ± 8.62 | 53.64 ± 13.31 | 505.65 ± 138.29 | 112.54 ± 92.97 |
| Plenter Phase | 62 | 905 ± 290 | 22.85 ± 5.97 | 56.29 ± 18.55 | 537.32 ± 199.44 | 167.54 ± 134.25 |
| Terminal Phase | 21 | 776 ± 292 | 27.86 ± 10.18 | 60.89 ± 20.65 | 614.05 ± 267.47 | 160.19 ± 152.83 |
| Decay Phase | 2 | 672 ± 226 | 27.85 ± 1.65 | 76.91 ± 32.92 | 868.88 ± 485.89 | 433.84 ± 283.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadopoulou, D.; Raptis, D.; Kazana, V.; Tsitsoni, T. Exploring Texture Diversity of Beech-Spruce-Fir Stands through Development Phase Analysis in the Frakto Virgin Forest of Greece. Diversity 2023, 15, 278. https://doi.org/10.3390/d15020278
Papadopoulou D, Raptis D, Kazana V, Tsitsoni T. Exploring Texture Diversity of Beech-Spruce-Fir Stands through Development Phase Analysis in the Frakto Virgin Forest of Greece. Diversity. 2023; 15(2):278. https://doi.org/10.3390/d15020278
Chicago/Turabian StylePapadopoulou, Dimitra, Dimitrios Raptis, Vassiliki Kazana, and Thekla Tsitsoni. 2023. "Exploring Texture Diversity of Beech-Spruce-Fir Stands through Development Phase Analysis in the Frakto Virgin Forest of Greece" Diversity 15, no. 2: 278. https://doi.org/10.3390/d15020278
APA StylePapadopoulou, D., Raptis, D., Kazana, V., & Tsitsoni, T. (2023). Exploring Texture Diversity of Beech-Spruce-Fir Stands through Development Phase Analysis in the Frakto Virgin Forest of Greece. Diversity, 15(2), 278. https://doi.org/10.3390/d15020278