
Revealing the Differences in Ulnaria acus and Fragilaria radians Distribution in Lake Baikal via Analysis of Existing Metabarcoding Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selecting Amplicons for Distinguishing F. radians and U. acus
2.2. Analysis of Published Metabarcoding Data
3. Results
3.1. Amplicon Selection and Testing
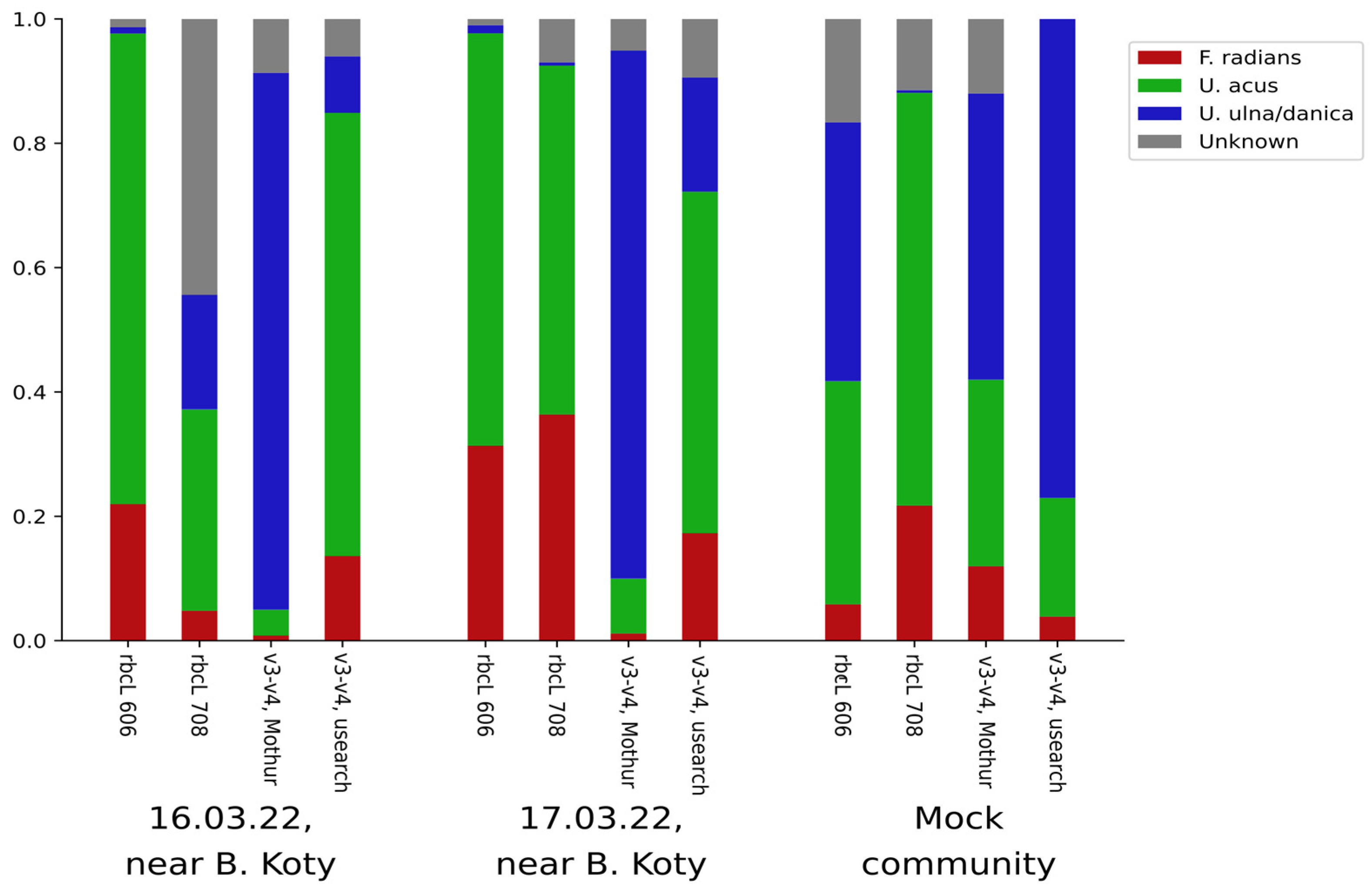
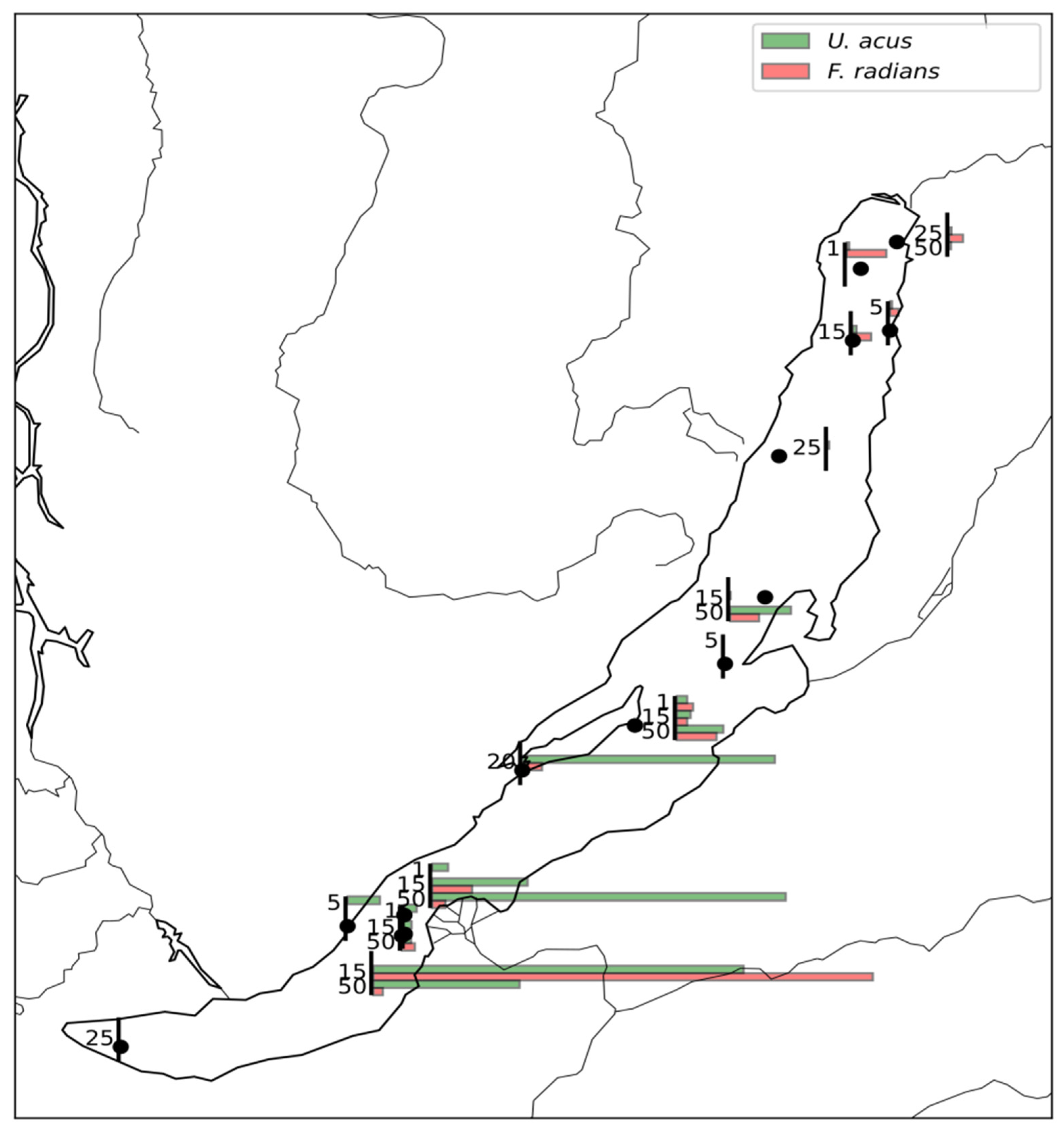
3.2. Analysis of Existing V4 Datasets
4. Discussion
4.1. Identifying the Target Species in Existing Data: Methodological Discussion
4.2. Autecology of U. acus and F. radians
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, D.M.; Round, F.E. Fragilariforma nom. nov., a new generic name for Neofragilaria Williams and Round. Diatom Res. 1988, 3, 265–267. [Google Scholar] [CrossRef]
- Shishlyannikov, S.M.; Zakharova, Y.R.; Volokitina, N.A.; Mikhailov, I.S.; Petrova, D.P.; Likhoshway, Y.V. A procedure for establishing an axenic culture of the diatom Synedra acus subsp. radians (Kütz.) Skabibitsch. from Lake Baikal. Limnol. Oceanogr. Meth. 2011, 9, 478–484. [Google Scholar] [CrossRef]
- Galachyants, Y.P.; Zakharova Yu, R.; Petrova, D.P.; Morozov, A.A.; Sidorov, I.A.; Marchenkov, A.M.; Logacheva, M.D.; Markelov, M.L.; Khabudaev, K.V.; Likhoshway, Y.V.; et al. Sequencing of the complete genome of an araphid pennate diatom Synedra acus subsp. radians from Lake Baikal. Dokl. Biochem. 2015, 461, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Popovskaya, G.I.; Likhoshway, Y.V.; Genkal, S.I.; Firsova, A.D. The role of endemic diatom algae in the phytoplankton of Lake Baikal. Hydrobiologia 2006, 568, 87–94. [Google Scholar] [CrossRef]
- Bashenkhaeva, M.V.; Zakharova, Y.R.; Petrova, D.P.; Khanaev, I.V.; Galachyants, Y.P.; Likhoshway, Y.V. Sub-ice microalgal and bacterial communities in freshwater Lake Baikal, Russia. Microb. Ecol. 2020, 70, 751–765. [Google Scholar] [CrossRef]
- Izmest’eva, L.R.; Moore, M.V.; Hampton, S.E.; Silow, E.A. Seasonal dynamics of common phytoplankton in Lake Baikal. Proc. Russ. Acad. Sci. Sci. Cent. 2006, 8, 191–196. [Google Scholar]
- Bondarenko, N.A.; Logacheva, N.F. Structural changes in phytoplankton of the littoral zone of Lake Baikal. Hydrobiol. J. 2017, 53, 16–24. [Google Scholar] [CrossRef]
- Mikhailov, I.S.; Galachyants, Y.P.; Bukin, Y.S.; Petrova, D.P.; Bashenkhaeva, M.V.; Sakirko, M.V.; Blinov, V.V.; Titova, L.A.; Zhakharova, Y.R.; Likhoshway, Y.V. Seasonal Succession and Coherence Among Bacteria and Microeukaryotes in Lake Baikal. Microb. Ecol. 2022, 84, 404–422. [Google Scholar] [CrossRef]
- Mikhailov, I.S.; Bukin, Y.S.; Zakharova, Y.R.; Usoltseva, M.V.; Galachyants, Y.P.; Sakirko, M.V.; Blinov, V.V.; Likhoshway, Y.V. Co-occurrence patterns between phytoplankton and bacterioplankton across the pelagic zone of Lake Baikal during spring. J. Microbiol. 2019, 57, 252–262. [Google Scholar] [CrossRef]
- Mikhailov, I.S.; Zakharova, Y.R.; Bukin, Y.S.; Galachyants, Y.P.; Petrova, D.P.; Sakirko, M.V.; Likhoshway, Y.V. Co-occurrence networks among bacteria and microbial eukaryotes of Lake Baikal during a spring phytoplankton bloom. Microb. Ecol. 2019, 77, 96–109. [Google Scholar] [CrossRef]
- Grachev, M.; Bukin, Y.; Blinov, V.; Khlystov, O.; Firsova, A.; Bashenkhaeva, M.; Kamshilo, O.; Titova, L.; Bairamova, E.; Bedoshvili, Y.; et al. Is a High Abundance of Spring Diatoms in the Photic Zone of Lake Baikal in July 2019 Due to an Upwelling Event? Diversity 2021, 13, 504. [Google Scholar] [CrossRef]
- Zakharova, Y.; Marchenkov, A.; Petrova, D.; Bukin, Y.; Morozov, A.; Bedoshvili, Y.; Podunay, Y.; Davidovich, O.; Davidovich, N.; Bondar, A.; et al. Delimitation of some taxa of Ulnaria and Fragilaria (Bacillariophyceae) based on genetic, morphological data and mating compatibility. Diversity, 2022; submitted. [Google Scholar]
- Kahlert, M.; Karjalainen, S.M.; Keck, F.; Kelly, M.; Ramon, M.; Rimet, F.; Schneider, S.; Tapolczai, K.; Zimmermann, J. Co-occurrence, ecological profiles and geographical distribution based on unique molecular identifiers of the common freshwater diatoms Fragilaria and Ulnaria. Ecol. Indic. 2022, 141, 109114. [Google Scholar] [CrossRef]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling expedition: Northwest Atlantic through eastern tropical Pacifc. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.; Breiner, H.W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef]
- Bradley, I.M.; Pinto, A.J.; Guest, J.S. Design and evaluation of Illumina MiSeq-compatible, 18S rRNA gene-specific primers for improved characterization of mixed phototrophic communities. Appl. Environ. Microbiol. 2016, 82, 5878–5891. [Google Scholar] [CrossRef] [Green Version]
- Stoof-Leichsenring, K.R.; Epp, L.S.; Trauth, M.H.; Tiedemann, R. Hidden diversity in diatoms of Kenyan Lake Naivasha: A genetic approach detects temporal variation. Mol. Ecol. 2012, 21, 1918–1930. [Google Scholar] [CrossRef]
- Bruder, K.; Medlin, L. Molecular assessment of phylogenetic relationships in selected species/genera in the naviculoid diatoms (Bacillariophyta). I. The genus Placoneis. Nova Hedwigia 2007, 85, 331. [Google Scholar] [CrossRef]
- Kermarrec, L.; Franc, A.; Rimet, F.; Chaumeil, P.; Humbert, J.F.; Bouchez, A. Next-generation sequencing to inventory taxonomic diversity in eukaryotic communities: A test for freshwater diatoms. Mol. Ecol. Resour. 2013, 13, 607–619. [Google Scholar] [CrossRef]
- Schloss, P.D. Reintroducing mothur: 10 years later. Appl. Environ. Microbiol. 2020, 86, e02343–e02419. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Annenkova, N.V.; Giner, C.R.; Logares, R. Tracing the origin of planktonic protists in an ancient lake. Microorganisms 2020, 8, 543. [Google Scholar] [CrossRef] [PubMed]
- David, G.M.; Moreira, D.; Reboul, G.; Annenkova, N.V.; Galindo, L.J.; Bertolino, P.; López-Archilla, A.I.; Jardillier, L.; López-Garcia, P. Environmental drivers of plankton protist communities along latitudinal and vertical gradients in the oldest and deepest freshwater lake. Environ. Microbiol. 2021, 23, 1436–1451. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. Peer J. 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplis, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Pomazkina, G.V.; Belykh, O.I.; Domysheva, V.M.; Sakirko, M.V.; Gnatovskii, R.Y. Structure and dynamics of the phytoplankton in Southern Baikal (Russia). Int. J. Algae 2010, 12, 64–79. [Google Scholar] [CrossRef]
- Pomazkina, G.V.; Rodionova, Y.V.; Khanaev, I.V.; Scherbakova, T.A. The State of Benthic Diatom Communities in Listvennichnyi Bay of Lake Baikal (Russia). Int. J. Algae 2018, 20, 387–392. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, M.; Albert, R.-L.; Anntila, E.-L.; Bengtsson, R.; Bigler, C.; Eskola, T.; Gälman, V.; Gottschalk, S.; Herlitz, E.; Jarlman, A.; et al. Harmonization is more important than experience—Results of the first Nordic-Baltic diatom intercalibration exercise 2007 (stream monitoring). J. Appl. Phycol. 2009, 21, 471–482. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morozov, A.; Galachyants, Y.; Marchenkov, A.; Zakharova, Y.; Petrova, D. Revealing the Differences in Ulnaria acus and Fragilaria radians Distribution in Lake Baikal via Analysis of Existing Metabarcoding Data. Diversity 2023, 15, 280. https://doi.org/10.3390/d15020280
Morozov A, Galachyants Y, Marchenkov A, Zakharova Y, Petrova D. Revealing the Differences in Ulnaria acus and Fragilaria radians Distribution in Lake Baikal via Analysis of Existing Metabarcoding Data. Diversity. 2023; 15(2):280. https://doi.org/10.3390/d15020280
Chicago/Turabian StyleMorozov, Alexey, Yuri Galachyants, Artem Marchenkov, Yulia Zakharova, and Darya Petrova. 2023. "Revealing the Differences in Ulnaria acus and Fragilaria radians Distribution in Lake Baikal via Analysis of Existing Metabarcoding Data" Diversity 15, no. 2: 280. https://doi.org/10.3390/d15020280