
Cetaceans in the Mediterranean Sea: Encounter Rate, Dominant Species, and Diversity Hotspots

 , , ,
, , ,  , , , ,
, , , ,  , , , , ,
, , , , ,  , , , , ,
, , , , ,  ,
,  ,
,  , , , , add
Show full author list
, , , , add
Show full author list
Abstract
:1. Introduction
2. Material and Methods
2.1. Encounter Rate
2.2. Species Prevalence
2.3. Shannon Diversity Index
3. Results
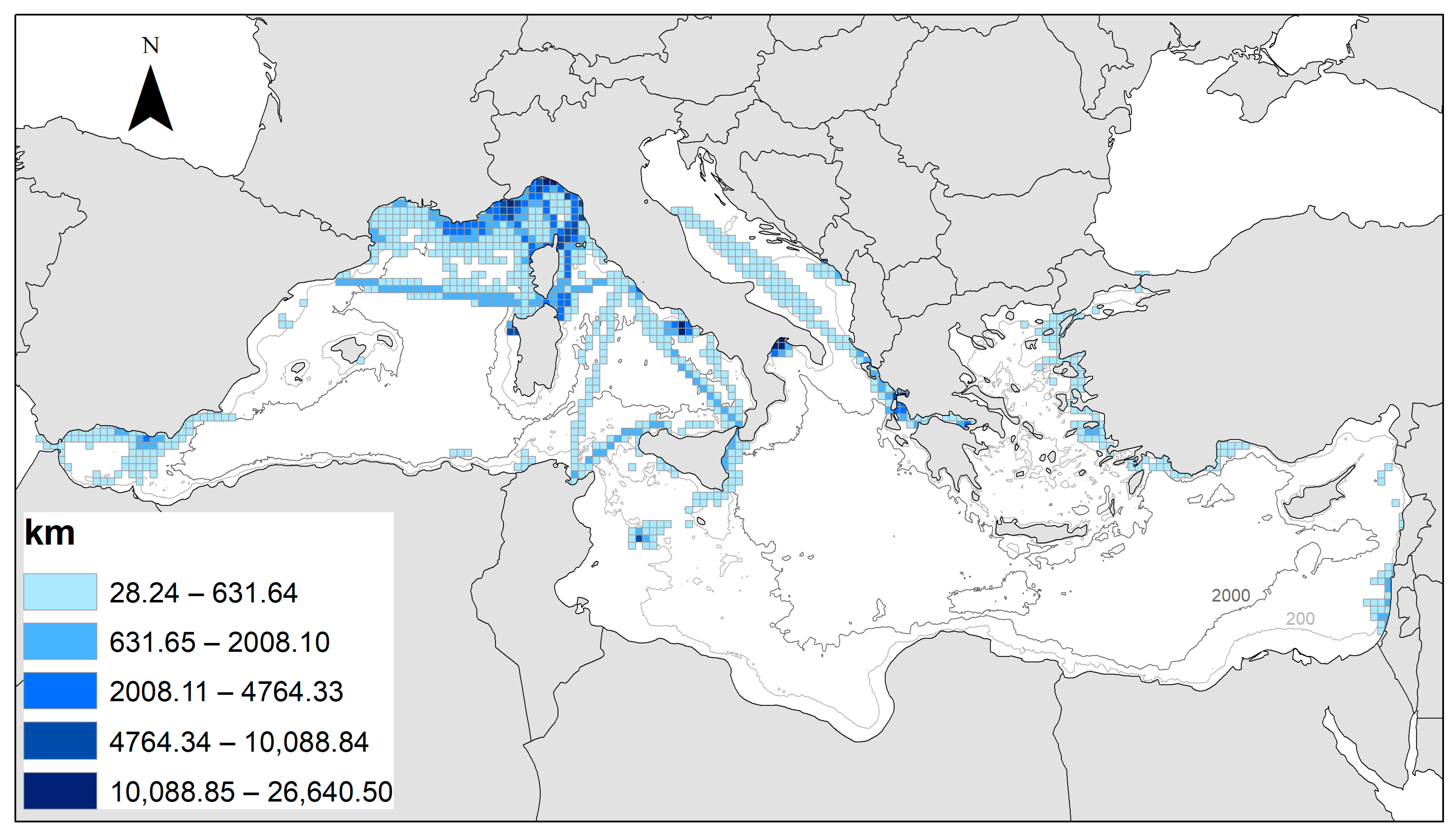
3.1. Mapping the Species ER and Prevalence
3.2. Encounter Rate and Shannon Diversity Index in the Mediterranean Sea
3.3. Comparison between Subzones
4. Discussion and Conclusions
Conservation Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pujo-Pay, M.; Conan, P.; Oriol, L.; Cornet-Barthaux, V.; Falco, C.; Ghiglione, J.-F.; Goyet, C.; Moutin, T.; Prieur, L. Integrated survey of elemental stoichiometry (C, N, P) from the western to eastern Mediterranean Sea. Biogeosciences 2011, 8, 883–899. [Google Scholar] [CrossRef]
- Tanhua, T.; Hainbucher, D.; Schröder, K.; Cardin, V.; Álvarez, M.; Civitarese, G. The Mediterranean Sea system: A review and an introduction to the special issue. Ocean Sci. Discuss. 2013, 10, 581–617. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Hsü, K.J.; Ryan, W.B.F.; Cita, M.B. Late Miocene desiccation of the Mediterranean. Nature 1973, 242, 240–244. [Google Scholar] [CrossRef]
- Krijgsman, W.; Hilgen, F.J.; Raffi, I.; Sierro, F.J.; Wilson, D.S. Chronology, causes and progression of the Messinian salinity crisis. Nature 1999, 400, 652–655. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. In Life in the Mediterranean Sea: A Look at Habitat Changes; Stambler, N., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012. [Google Scholar]
- Notarbartolo di Sciara, G.; Tonay, A.M. Conserving Whales, Dolphins and Porpoises in the Mediterranean Sea, Black Sea and Adjacent Areas: An ACCOBAMS Status Report, 2021; ACCOBAMS: Monaco, 2021. [Google Scholar]
- Notarbartolo di Sciara, G. Cetacean species occurring in the Mediterranean and Black Seas. In Cetaceans of the Mediterranean and Black Seas: State of Knowledge and Conservation Strategies; A report to the ACCOBAMS Secretariat; Notarbartolo di Sciara, G., Ed.; ACCOBAMS: Monaco, 2002; Section 3; 17p. [Google Scholar]
- Boisseau, O.; Lacey, C.; Lewis, T.; Moscrop, A.; Danbolt, M.; McLanaghan, R. Encounter rates of cetaceans in the Mediterranean Sea and contiguous Atlantic area. J. Mar. Biol. Assoc. UK 2010, 90, 1589–1599. [Google Scholar] [CrossRef]
- Arcangeli, A.; Campana, I.; Bologna, M.A. Influence of seasonality on cetacean diversity, abundance, distribution and habitat use in the western Mediterranean Sea: Implications for conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 995–1010. [Google Scholar] [CrossRef]
- Panigada, S.; Lauriano, G.; Donovan, G.; Pierantonio, N.; Cañadas, A.; Vázquez, J.A.; Burt, L. Estimating cetacean density and abundance in the Central and Western Mediterranean Sea through aerial surveys: Implications for management. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 141, 41–58. [Google Scholar] [CrossRef]
- Pace, D.S.; Giacomini, G.; Campana, I.; Paraboschi, M.; Pellegrino, G.; Silvestri, M.; Alessi, J.; Angeletti, D.; Cafaro, V.; Pavan, G.; et al. An integrated approach for cetacean knowledge and conservation in the central Mediterranean Sea using research and social media data sources. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1302–1323. [Google Scholar] [CrossRef]
- Gaskin, D.E. The harbour porpoise Phocoena phocoena (L.): Regional populations, status, and information on direct and indirect catches. Rep. Int. Whal. Comm. 1984, 34, 569–586. [Google Scholar]
- Öztürk, B.; Öztürk, A.A. On the biology of the Turkish straits system. Bull. Inst. Oceanogr. Numero Spec. 1996, 17, 205–217. [Google Scholar]
- Cucknell, A.C.; Frantzis, A.; Boisseau, O.; Romagosa, M.; Ryan, C.; Tonay, A.M.; Alexiadou, P.; Öztürk, A.A.; Moscrop, A. Harbour porpoises in the Aegean Sea, Eastern Mediterranean: The species’ presence is confirmed. Mar. Biodivers. Rec. 2016, 9, 1–13. [Google Scholar] [CrossRef]
- Frantzis, A.; Gordon, J.; Hassidis, G.; Komnenou, A. The enigma of the harbour porpoise presence in the Mediterranean Sea. Mar. Mamm. Sci. 2001, 17, 937–944. [Google Scholar] [CrossRef]
- Fontaine, M.C. Chapter Eleven—Harbour Porpoises, Phocoena phocoena, in the Mediterranean Sea and Adjacent Regions: Biogeographic Relicts of the Last Glacial Period. Adv. Mar. Biol. 2016, 75, 333–358. [Google Scholar] [CrossRef]
- Bearzi, G.; Genov, T.; Natoli, A.; Gonzalvo, J.; Pierce, G.J. Delphinus delphis (Inner Mediterranean subpopulation) (errata version published in 2022). In The IUCN Red List of Threatened Species; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2002; p. e.T189865869A210844387. [Google Scholar] [CrossRef]
- Cagnolaro, L. Profilo sistematico e tipologico delle raccolte di cetacei attuali dei musei italiani. Suppl. Museol. Sci. 1996, 13, 193–212. [Google Scholar]
- Cagnolaro, L.; Podestà, M.; Affronte, M.; Agnelli, P.; Cancelli, F.; Capanna, E.; Carlini, R.; Cataldini, G.; Cozzi, B.; Insacco, G. Collections of extant cetaceans in Italian museums and other scientific institutions. A comparative review. Nat. Hist. Sci. 2012, 153, 145–202. [Google Scholar] [CrossRef]
- Verborgh, P.; Gauffier, P.; Esteban, R.; Giménez, J.; Cañadas, A.; Salazar-Sierra, J.M.; de Stephanis, R. Conservation Status of Long-Finned Pilot Whales, Globicephala melas, in the Mediterranean Sea. Adv. Mar. Biol. 2016, 75, 173–203. [Google Scholar] [CrossRef]
- Podestà, M.; Azzellino, A.; Cañadas, A.; Frantzis, A.; Moulins, A.; Rosso, M.; Tepsich, P.; Lanfredi, C. Cuvier’s beaked whale, Ziphius cavirostris, distribution and occurrence in the Mediterranean Sea: High-use areas and conservation threats. Adv. Mar. Biol. 2016, 75, 103–114. [Google Scholar] [CrossRef]
- Mannocci, L.; Roberts, J.J.; Halpin, P.N.; Authier, M.; Boisseau, O.; Bradai, M.N.; Cañadas, A.; Chicote, C.; David, L.; Di-Méglio, N.; et al. Assessing cetacean surveys throughout the Mediterranean Sea: A gap analysis in environmental space. Sci. Rep. 2018, 8, 3126. [Google Scholar] [CrossRef]
- Gnone, G.; Bellingeri, M.; Paraboschi, M.; Campana, I.; Alessi, J.; Nuti, S.; Salvioli, F.; Tepsich, P.; Rosso, M.; Moulins, A.; et al. TursioMed: Final Scientific Report. The final scientific report of the TursioMed project, funded by Blue Planet Virginia Böger Stiftung X.X. 2021; unpublished work. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423 and 623–656. [Google Scholar] [CrossRef]
- Silverman, B.W. Density Estimation for Statistics and Data Analysis; Chapman and Hall: New York, NY, USA, 1986. [Google Scholar]
- Correia, A.M.; Tepsich, P.; Rosso, M.; Caldeira, R.; Sousa-Pinto, I. Cetacean occurrence and spatial distribution: Habitat modelling for offshore waters in the Portuguese EEZ (NE Atlantic). J. Mar. Syst. 2015, 143, 73–85. [Google Scholar] [CrossRef]
- Correia, A.M.; Gandra, M.; Liberal, M.; Valente, R.; Gil, Á.; Rosso, M.; Pierce, G.J.; Sousa-Pinto, I. A dataset of cetacean occurrences in the Eastern North Atlantic. Sci. Data 2019, 6, 177. [Google Scholar] [CrossRef]
- Panigada, S.; Notarbartolo di Sciara, G.; Zanardelli Panigada, M.; Airoldi, S.; Borsani, J.F.; Jahoda, M. Fin whales (Balaenoptera physalus) summering in the Ligurian Sea: Distribution, encounter rate, mean group size and relation to physiographic variables. J. Cetacean Res. Manag. 2005, 7, 137–145. [Google Scholar]
- Cañadas, A.; Sagarminaga, R.; Garcia-Tiscar, S. Cetacean distribution related with depth and slope in the Mediterranean waters off southern Spain. Deep Sea Res. 2002, 49, 2053–2073. [Google Scholar] [CrossRef]
- Panigada, S.; Zanardelli, M.; Mackenzie, M.; Donovan, C.; Melin, F.; Hammond, P.S. Modelling Habitat Preferences for Fin Whales and Striped Dolphins in the Pelagos Sanctuary (Western Mediterranean Sea) with Physiographic and Remote Sensing Variables. Remote Sens. Environ. 2008, 112, 3400–3412. [Google Scholar] [CrossRef]
- Notarbartolo di Sciara, G.; Agardy, T.; Hyrenbach, D.; Scovazzi, T.; van Klaveren, P. The Pelagos Sanctuary for Mediterranean marine mammals. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 367–391. [Google Scholar] [CrossRef]
- Bearzi, G. Delphinus delphis (Mediterranean subpopulation). In The IUCN Red List of Threatened Species; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2003; p. e.T41762A10557372. [Google Scholar] [CrossRef]
- Frantzis, A.; Alexiadou, P.; Paximadis, G.; Politi, E.; Gannier, A. Current knowledge of the cetacean fauna of the Greek Seas. J. Cetacean Res. Manag. 2003, 5, 219–232. [Google Scholar]
- Gnone, G.; Bellingeri, M.; Molinari, Y.; Dhermain, F.; Labach, H.; Díaz López, B.; David, L.; Di Meglio, N.; Azzinari, G.; Azzinari, C.; et al. The Seabed makes the dolphins: Physiographic features shape the size and structure of the bottlenose dolphin geographical units. J. Mar. Sci. Eng. 2022, 10, 1036. [Google Scholar] [CrossRef]
- Pace, D.S.; Panunzi, G.; Arcangeli, A.; Moro, S.; Jona-Lasinio, G.; Martino, S. Seasonal distribution of an opportunistic apex predator (Tursiops truncatus) in marine coastal habitats of the Western Mediterranean Sea. Front. Mar. Sci. 2022, 9, 939692. [Google Scholar] [CrossRef]
- Gannier, A. Summer distribution and relative abundance of delphinids in the Mediterranean Sea. Rev. D’ecologie Terre Société Natl. Prot. Nat. 2005, 60, 223–238. [Google Scholar] [CrossRef]
- Azzellino, A.; Gaspari, S.; Airold, S.; Nani, B. Habitat use and preferences of cetaceans along the continental slope and the adjacent pelagic waters in the western Ligurian Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 296–323. [Google Scholar] [CrossRef]
- Pace, D.S.; Arcangeli, A.; Mussi, B.; Vivaldi, C.; Ledon, C.; Lagorio, S.; Giacomini, G.; Pavan, G.; Ardizzone, G. Habitat suitability modeling in different sperm whale social groups. J. Wildl. Manag. 2018, 82, 1062–1073. [Google Scholar] [CrossRef]
- Awbery, T.; Akkaya, A.; Lyne, P.; Rudd, L.; Hoogenstrijd, G.; Nedelcu, M.; Kniha, D.; Erdoğan, M.A.; Persad, C.; Amaha Öztürk, A.; et al. Spatial Distribution and Encounter Rates of Delphinids and Deep Diving Cetaceans in the Eastern Mediterranean Sea of Turkey and the Extent of Overlap With Areas of Dense Marine Traffic. Front. Mar. Sci. 2022, 9, 860242. [Google Scholar] [CrossRef]
- Videau, C.; Sournia, A.; Prieur, L.; Fiala, M. Phytoplankton and primary production characteristics at selected sites in the geostrophic Almeria-Oran front system (SW Mediterranean Sea). J. Mar. Syst. 1994, 5, 235–250. [Google Scholar] [CrossRef]
- Morán, X.A.G.; Estrada, M. Short-term variability of photosynthetic parameters and particulate and dissolved primary production in the Alboran Sea (SW Mediterranean). Mar. Ecol. Prog. Ser. 2001, 212, 53–67. [Google Scholar] [CrossRef]
- Aïssi, M.; Arcangeli, A.; Crosti, R.; Daly Yahia, M.N.; Loussaief, B.; Moulins, A.; Pellegrino, G.; Rosso, M.; Tringali, L.M.; Tepsich, P. Cetacean occurrence and spatial distribution in the central Mediterranean Sea using ferries as platform of observation. Russ. J. Mar. Biol. 2015, 41, 343–350. [Google Scholar] [CrossRef]
- IUCN-MMPATF. Shelf of the Gulf of Lyon IMMA Factsheet. IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. 2017. Available online: https://www.marinemammalhabitat.org/portfolio-item/shelf-gulf-lion/ (accessed on 9 August 2022).
- Ricci, P.; Carlucci, R.; Capezzuto, F.; Carluccio, A.; Cipriano, G.; D’Onghia, G.; Maiorano, P.; Sion, L.; Tursi, A.; Libralato, S. Contribution of Intermediate and High Trophic Level Species to Benthic-Pelagic Coupling: Insights From Modelling Analysis. Front. Mar. Sci. 2022, 9, 887464. [Google Scholar] [CrossRef]
- Azzellino, A.; Airoldi, S.; Lanfredi, C.; Podestà, M.; Zanardelli, M. Cetacean response to environmental and anthropogenic drivers of change: Results of a 25-year distribution study in the northwestern Mediterranean Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 146, 104–117. [Google Scholar] [CrossRef]
- Azzellino, A.; Fossi, M.C.; Gaspari, S.; Lanfredi, C.; Lauriano, G.; Marsili, L.; Panigada, S.; Podestà, P. An index based on the biodiversity of cetacean species to assess the Environmental Status of marine ecosystems. Mar. Environ. Res. 2014, 100, 94–111. [Google Scholar] [CrossRef]
- ACCOBAMS. Estimates of Abundance and Distribution of Cetaceans, Marine Mega-Fauna and Marine Litter in the Mediterranean Sea from 2018–2019 Surveys; Panigada, S., Boisseau, O., Canadas, A., Lambert, C., Laran, S., McLanaghan, R., Moscrop, A., Eds.; ACCOBAMS Survey Initiative Project: Monaco, 2021; p. 177. [Google Scholar]
- DIRECTIVE 2008/56/EC of the European Parliament and of the Council, 17 June 2008. Establishing a framework for community action in the field of marine environmental policy (Marine Strategy Framework Directive). Off. J. Eur. Union 2008, 164, 19–40.
- IUCN-MMPATF. Alborán Sea IMMA. Full Accounts of Mediterranean IMMA Factsheet. IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. 2017. Available online: https://www.marinemammalhabitat.org/wp-content/uploads/imma-factsheets/Mediterranean/Alboran-Sea-Mediterranean.pdf (accessed on 9 August 2022).
- IUCN-MMPATF. North West Mediterranean Sea, Slope and Canyon system IMMA Factsheet. IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. 2017. Available online: https://www.marinemammalhabitat.org/portfolio-item/north-western-mediterranean-sea-slope-canyon-system/ (accessed on 9 August 2022).
- IUCN-MMPATF. Campanian and Pontino Archipelagos IMMA. Full Accounts of Mediterranean IMMA Factsheet. IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. 2017. Available online: https://www.marinemammalhabitat.org/portfolioitem/campanian-and-pontino-archipelagos/ (accessed on 18 August 2022).
- IUCN-MMPATF Waters of Ischia and Ventotene IMMA Factsheet. IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. 2017. Available online: https://www.marinemammalhabitat.org/portfolioitem/waters-ischia-ventotene/ (accessed on 18 October 2022).
- IMMA E-ATLAS. Available online: https://www.marinemammalhabitat.org/imma-eatlas/ (accessed on 18 October 2022).
- IUCN-MMPATF. Hellenic Trench IMMA. Full Accounts of Mediterranean IMMA Factsheet. IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force. 2017. Available online: https://www.marinemammalhabitat.org/portfolioitem/hellenic-trench/ (accessed on 9 August 2022).
- Panigada, S.; Donovan, G.P.; Druon, J.N.; Lauriano, G.; Pierantonio, N.; Pirotta, E.; Zanardelli, M.; Zerbini, A.N.; Notarbartolo di Sciara, G. Satellite tagging of Mediterranean fin whales: Working towards the identification of critical habitats and the focussing of mitigation measures. Sci. Rep. 2017, 7, 3365. [Google Scholar] [CrossRef] [PubMed]
- Huertas, I.E.; Ríos, A.F.; García-Lafuente, J.; Navarro, G.; Makaoui, A.; Sánchez-Román, A.; Rodriguez-Galvez, S.; Orbi, A.; Ruíz, J.; Pérez, F.F. Atlantic forcing of the Mediterranean oligotrophy. Glob. Biogeochem. Cycles 2012, 26, GB2022. [Google Scholar] [CrossRef]
- Béthoux, J.P. Le phosphore et l’azote en Méditerranée, bilans et fertilité potentielle. Mar. Chem. 1981, 10, 141–158. [Google Scholar] [CrossRef]
- Estrada, M. Primary production in the northwestern Mediterranean. Sci. Mar. 1996, 60, 55–64. [Google Scholar]
- Ludwig, W.; Bouwman, A.F.; Dumont, E.; Lespinas, F. Water and nutrient fluxes from major Mediterranean and Black Sea rivers: Past and future trends and their implications for the basin-scale budgets. Glob. Biogeochem. Cycles 2010, 24, GB0A13. [Google Scholar] [CrossRef]



























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Group | Study Area | Subzone | Sampling Period | Sampling Effort (km) | N Sightings | Data Source |
|---|---|---|---|---|---|---|
| Alnilam Research and Conservation | Alboran Sea | M1 | 2004–2011 | 27,970 | 1396 | Intercet |
| SUBMON | Catalonia | M2 | 2010–2018 | 1504 | 22 | Intercet |
| Association BREACH | Gulf of Lion | M2 | 2013–2016 | 3700 | 48 | Intercet |
| EcoOcean Institut (and partners *) | Gulf of Lion, French Riviera, Corsica W | M2 | 2005–2015 | 48,735 | 1835 | Intercet |
| Accademia del Leviatano (FLT Med Network) | NW Mediterranean | M2 | 2012–2015 | 57,567 | 1006 | Intercet |
| Tethys Research Institute—CSR | Liguria W, French Riviera | M2 | 2004–2018 | 83,377 | 2661 | Intercet |
| Fondazione CIMA (FLT Med Network) | Ligurian Sea | M2 | 2008–2013 | 46,690 | 2057 | Intercet |
| Fondazione Acquario di Genova | E Liguria | M2 | 2004–2018 | 67,456 | 1177 | Intercet |
| University of Genoa—DISTAV | Eastern Liguria, Tuscany | M2 | 2005–2008 | 15,513 | 49 | Intercet |
| CE.TU.S Cetacean Research Centre | Tuscany | M2 | 2004–2018 | 23,580 | 507 | Intercet |
| ARPAT | Tuscany | M2 | 2010–2011 | 2851 | 26 | Intercet |
| Association CARI | Northern Corsica | M2 | 2013–2015 | 1301 | 5 | Intercet |
| MIRACETI | Gulf of Lion, French Riviera, Corsica | M2 | 2004–2018 | 37,359 | 665 | Intercet |
| Office de l’Environnement de la Corse | Strait of Bonifacio | M2 | 2009–2012 | 3249 | 42 | Intercet |
| SEA ME Sardinia onlus | Northeast Sardinia | M2 | 2011–2013 | 4291 | 507 | Intercet |
| Bottlenose Dolphin Research Institute | Northeast Sardinia | M2 | 2004–2013 | 8586 | 334 | Intercet |
| Associazione CRAMA | Northwest Sardinia | M2 | 2010, 2016 | 376 | 12 | Intercet |
| MareTerra Onlus | NE Sardinia, Lampedusa | M2 | 2012–2016 | 11,853 | 218 | Intercet |
| University of Pisa (FLT Med Network) | Northern Tyrrhenian Sea | M2 | 2008–2016 | 71,147 | 653 | Intercet |
| University La Sapienza | Eastern Tyrrhenian Sea | M3 | 2017–2018 | 833 | 15 | Data owner |
| Oceanomare Delphis Onlus | Eastern Tyrrhenian Sea | M3 | 2004–2016 | 54,772 | 1083 | Intercet |
| Ketos (FLT Med Network) | Tyrrhenian Sea, Strait of Sicily | M3, M4 | 2004–2017 | 53,260 | 673 | Intercet |
| MeRiS-Mediterraneo Ricerca e Sviluppo APS | Strait of Sicily | M4 | 2016, 2018 | 1531 | 9 | Intercet |
| University of Torino—DBIOS | Lampedusa (Strait of Sicily) | M4 | 2004–2006 | 7650 | 209 | Intercet |
| Jonian Dolphin Conservation | Gulf of Taranto (NW Ionian Sea) | M5 | 2009–2017 | 52,505 | 893 | Data owner |
| Tethys Research Institute—IDP | Gulf of Ambracia | M5 | 2004–2016 | 46,749 | 859 | Intercet |
| Thalassa | Ionian Greece | M5 | 2004–2018 | 12,302 | 203 | Intercet |
| Gaia Res. Institute and University of Torino—DBIOS (FLT Med Network) | NE Ionian Sea and Adriatic Sea | M5–M6 | 2014–2018 | 23,642 | 123 | Intercet |
| DMAD—Marine Mammals Research Association | Southeast Adriatic, Bosphorus, Aegean Sea, Eastern Mediterranean | M7 | 2011–2018 | 5727 | 229 | Intercet |
| Archipelagos Inst. of Marine Conservation | Aegean Sea | M7 | 2017–2018 | 4051 | 95 | Data owner |
| Istanbul University and Turkish Marine Research Foundation | Aegean Sea, Eastern Mediterranean | M7–M8 | 2005–2008 | 6901 | 117 | Intercet |
| Morris Kahn Res. Station, Un. of Haifa | Eastern Mediterranean | M8 | 2017–2018 | 8960 | 35 | Intercet |
| TOTAL | 795,989 | 17,763 |
| Species | N Sightings | |||
|---|---|---|---|---|
| All Data | 0–200 m | 200–2000 m | >2000 m | |
| Stenella coeruleoalba | 8954 (50.4%) | 450 (10.0%) | 5446 (65.8%) | 3058 (61.4%) |
| Tursiops truncatus | 4043 (22.8%) | 3622 (80.6%) | 389 (4.7%) | 32 (0.6%) |
| Balaenoptera physlaus | 2196 (12.4%) | 92 (2.0%) | 673 (8.1%) | 1431 (28.7%) |
| Physeter macrocephalus | 1001 (5.6%) | 5 (0.1%) | 717 (8.7%) | 279 (5.6%) |
| Delphinus delphis | 695 (3.9%) | 288 (6.4%) | 360 (4.3%) | 47 (0.9%) |
| Ziphius cavirostris | 312 (1.8%) | 1 (0.02%) | 277 (3.3%) | 34 (0.7%) |
| Globicephala melas | 294 (1.7%) | 0 (0.0%) | 238 (2.9%) | 56 (1.1%) |
| Grampus griseus | 239 (1.3%) | 10 (0.2%) | 182 (2.2%) | 47 (0.9%) |
| Phocoena phocoena relicta | 26 (0.1%) | 26 (0.6) | 0 (0.0%) | 0 (0.0%) |
| Total | 17,760 | 4494 | 8282 | 4984 |
| Alboran Sea | A (<4.5 m) | B (4.75–10.5 m) | C (12–29 m) |
|---|---|---|---|
| N cells | 84 | 84 | NA |
| Total effort (km) | 10,664 | 14,990 | NA |
| Average ER (±SE) | 0.0104 (±0.002) | 0.0224 (±0.004) | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gnone, G.; Bellingeri, M.; Airoldi, S.; Gonzalvo, J.; David, L.; Di-Méglio, N.; Cañadas, A.M.; Akkaya, A.; Awbery, T.; Mussi, B.; et al. Cetaceans in the Mediterranean Sea: Encounter Rate, Dominant Species, and Diversity Hotspots. Diversity 2023, 15, 321. https://doi.org/10.3390/d15030321
Gnone G, Bellingeri M, Airoldi S, Gonzalvo J, David L, Di-Méglio N, Cañadas AM, Akkaya A, Awbery T, Mussi B, et al. Cetaceans in the Mediterranean Sea: Encounter Rate, Dominant Species, and Diversity Hotspots. Diversity. 2023; 15(3):321. https://doi.org/10.3390/d15030321
Chicago/Turabian StyleGnone, Guido, Michela Bellingeri, Sabina Airoldi, Joan Gonzalvo, Léa David, Nathalie Di-Méglio, Ana M. Cañadas, Aylin Akkaya, Tim Awbery, Barbara Mussi, and et al. 2023. "Cetaceans in the Mediterranean Sea: Encounter Rate, Dominant Species, and Diversity Hotspots" Diversity 15, no. 3: 321. https://doi.org/10.3390/d15030321
APA StyleGnone, G., Bellingeri, M., Airoldi, S., Gonzalvo, J., David, L., Di-Méglio, N., Cañadas, A. M., Akkaya, A., Awbery, T., Mussi, B., Campana, I., Azzolin, M., Dede, A., Tonay, A. M., Monaco, C., Pellegrino, G., Tepsich, P., Moulins, A., Arcangeli, A., ... Azzellino, A. (2023). Cetaceans in the Mediterranean Sea: Encounter Rate, Dominant Species, and Diversity Hotspots. Diversity, 15(3), 321. https://doi.org/10.3390/d15030321