

On the Occurrence of Arbuscular Mycorrhizal Fungi in a Bryophyte Community of Punta Lara Natural Reserve, Buenos Aires, Argentina

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
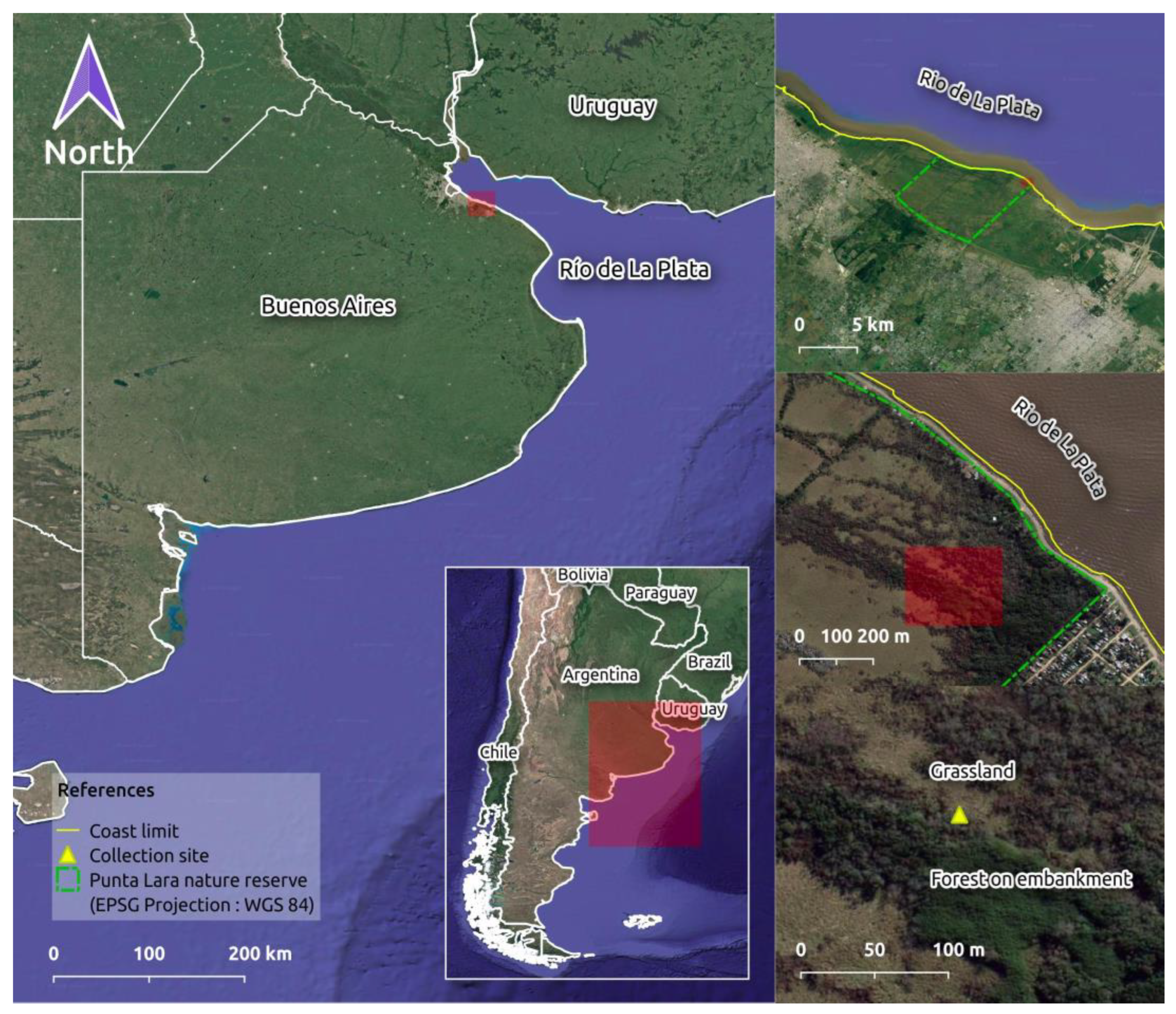
2.1. Study Area
2.2. Sampling
2.3. Laboratory Analysis
2.4. Molecular Characterization of Fungi Associated with the Bryophyte Community
3. Results
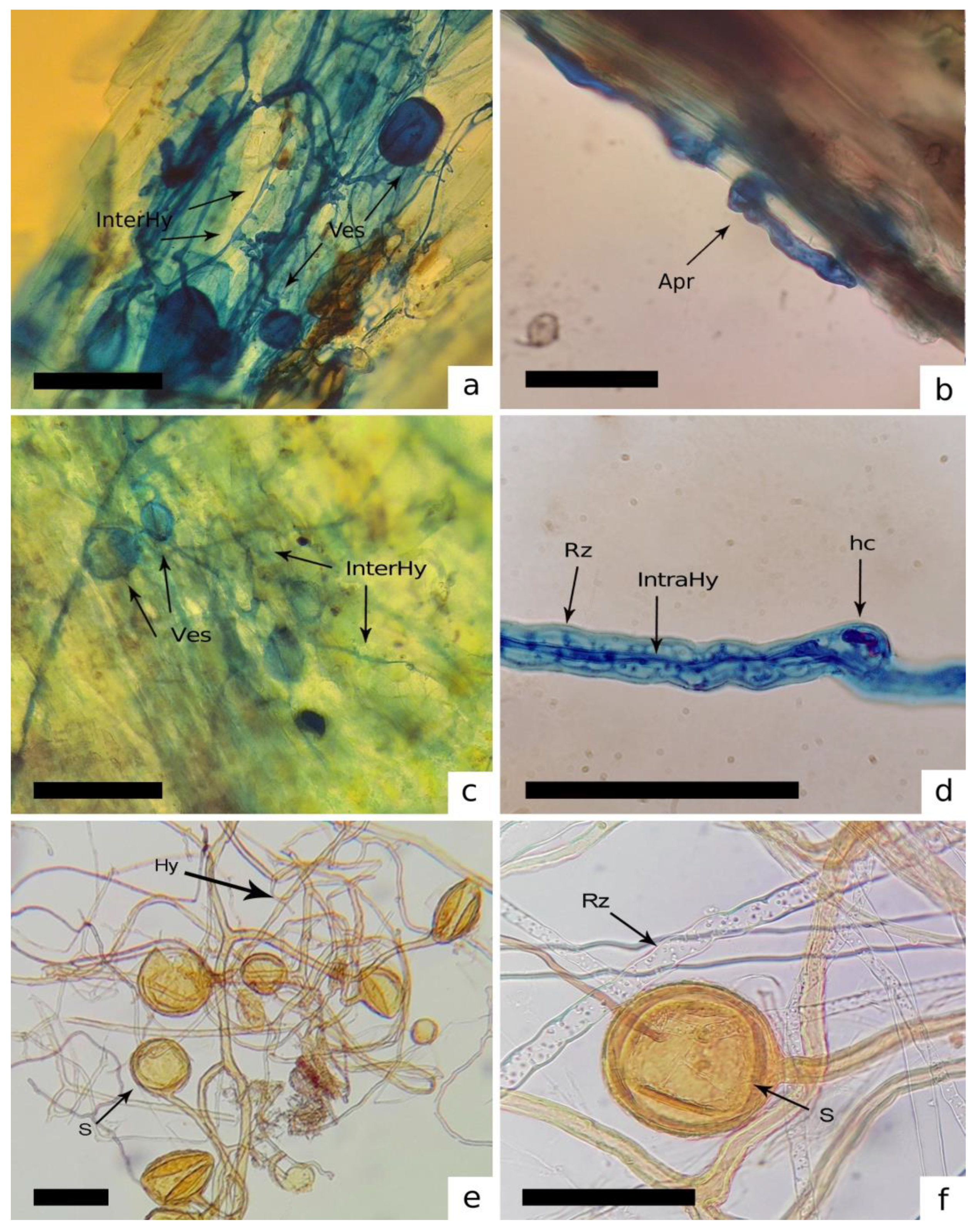
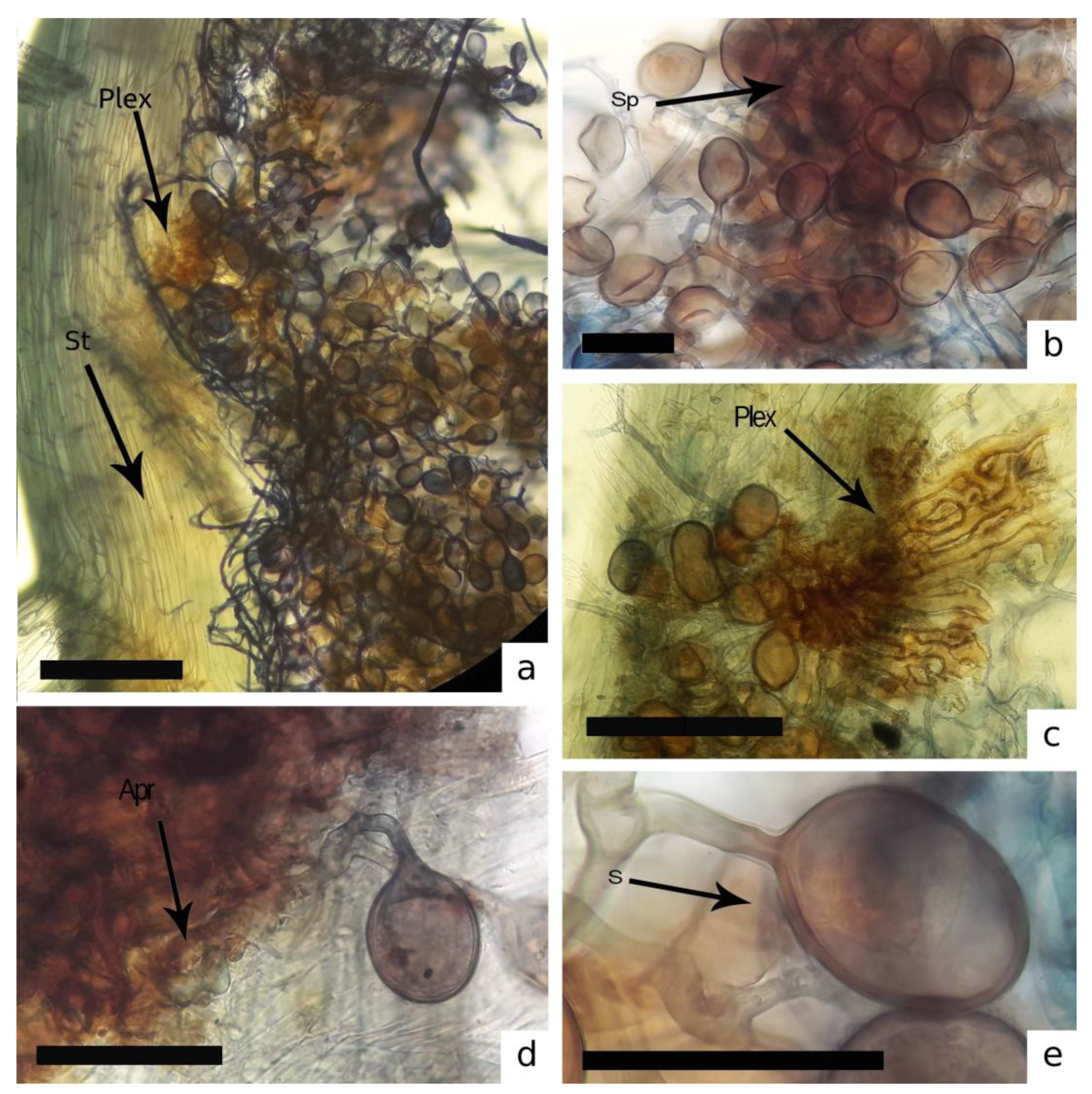
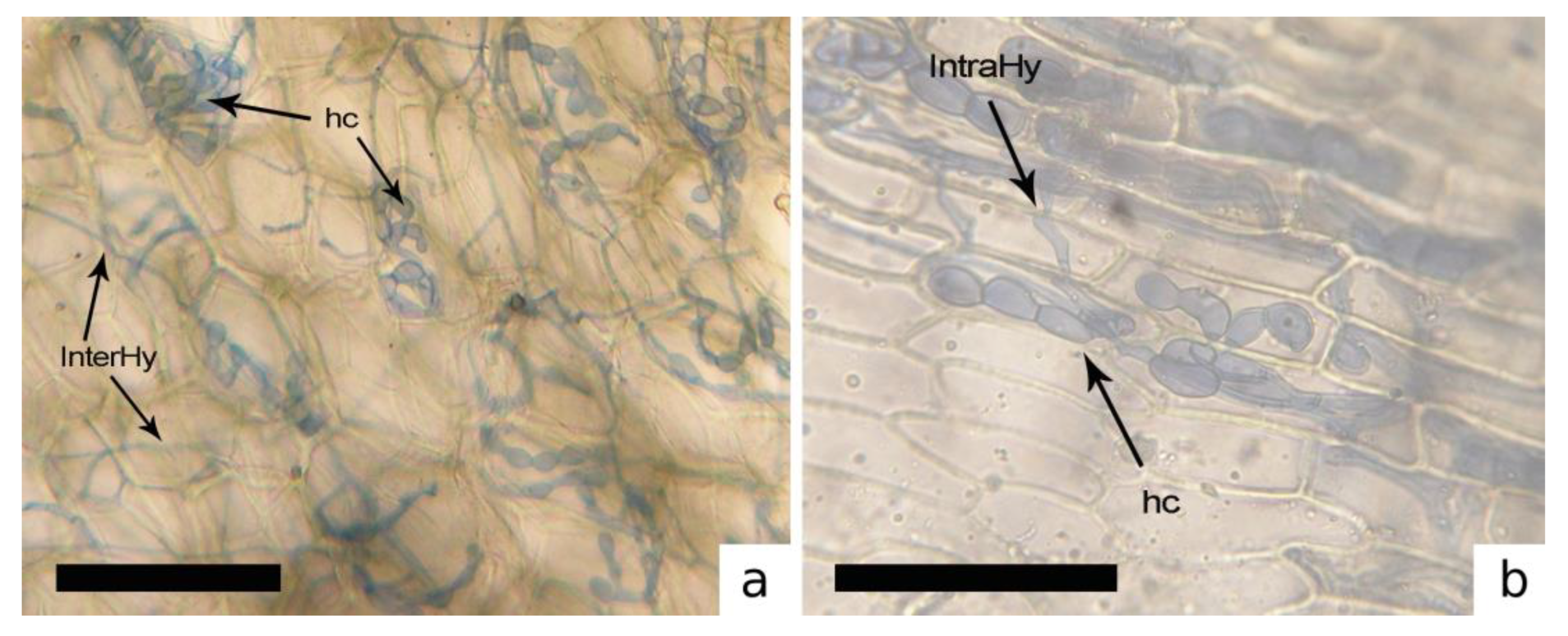
3.1. AMF Associated with Bryophytes Community
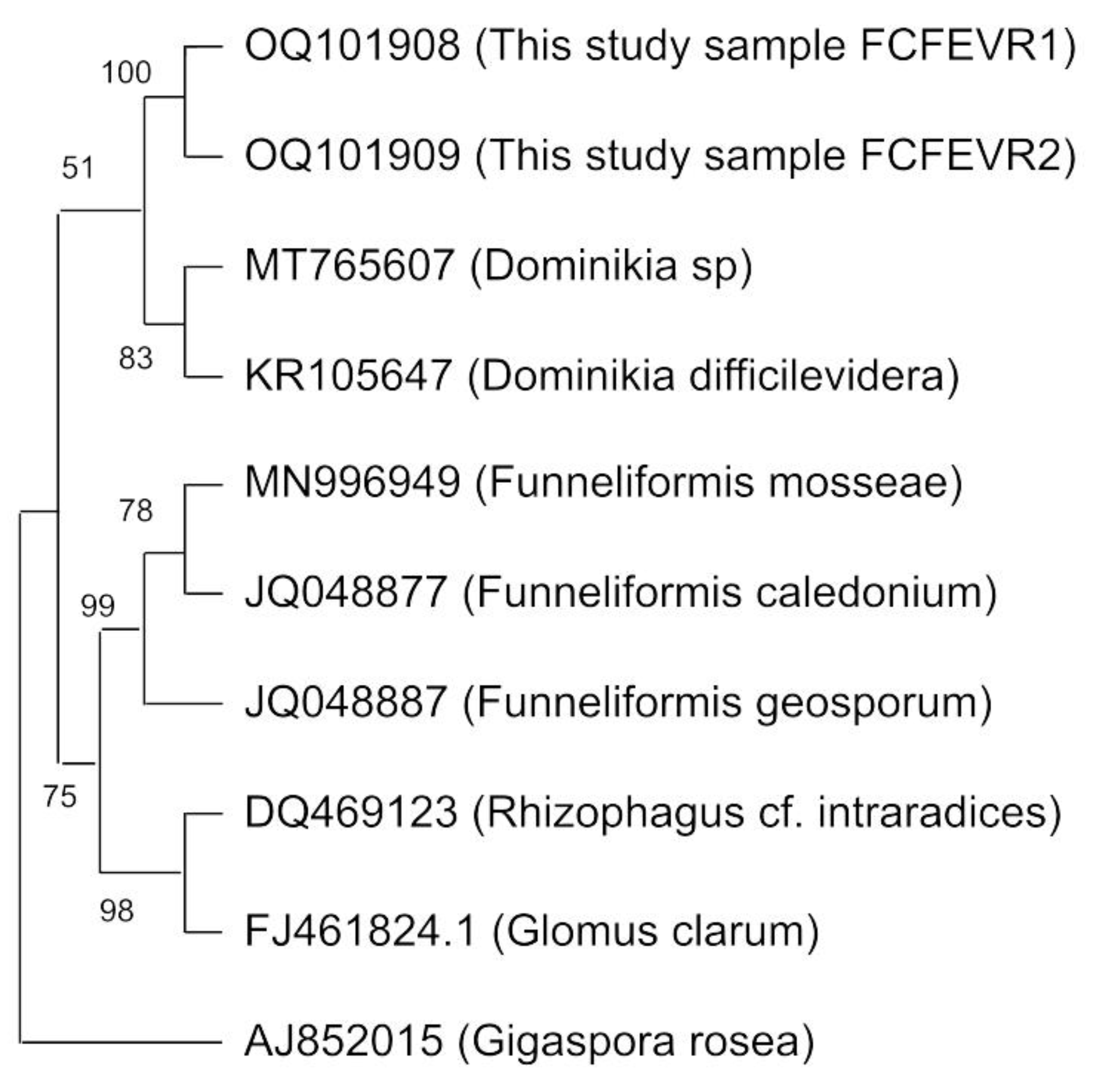
3.2. Molecular Characterization of Fungi Associated to the Bryophyte Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.L.; Li, L.; Wang, B.; Chen, Z.; Knoop, V.; Groth-Malonek, M.; Estabrook, G.F. The deepest divergences in land plants inferred from phylogenomic evidence. Proc. Natl. Acad. Sci. USA 2006, 103, 15511–15516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic: London, UK, 2008. [Google Scholar] [CrossRef]
- De Sousa, F.; Foster, P.G.; Donoghue, P.C.; Schneider, H.; Cox, C.J. Nuclear protein phylogenies support the monophyly of the three bryophyte groups (Bryophyta Schimp.). New Phytol. 2019, 222, 565–575. [Google Scholar] [CrossRef]
- Redecker, D.; Kodner, R.; Graham, L.E. Glomalean fungi from the Ordovician. Science 2000, 289, 1920–1921. [Google Scholar] [CrossRef] [Green Version]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Fossil Fungi; Academic: Burlington, MA, USA, 2015. [Google Scholar]
- Shaw, A.J.; Szövényi, P.; Shaw, B. Bryophyte diversity and evolution: Windows into the early evolution of land plants. Am. J. Bot. 2011, 98, 352–369. [Google Scholar] [CrossRef]
- Stech, M.; Quandt, D. 20,000 species and five key markers: The status of molecular bryophyte phylogenetics. Phytotaxa 2014, 9, 196–228. [Google Scholar] [CrossRef]
- Parke, J.L.; Linderman, R.G. Association of vesicular–arbuscular mycorrhizal fungi with the moss Funaria hygrometrica. Canad. J. Bot. 1980, 58, 1898–1904. [Google Scholar] [CrossRef]
- Rabatin, S.C. The occurrence of the vesicular-arbuscularmycorrhizal fungus Glomus tenuis with moss. Mycologia 1980, 72, 191–195. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, L.D. Arbuscular mycorrhizal structure and fungi associated with mosses. Mycorrhiza 2007, 17, 319–325. [Google Scholar] [CrossRef]
- Döbbeler, P. Biodiversity of bryophilous ascomycetes. Biodivers. Conserv. 1997, 6, 721–738. [Google Scholar] [CrossRef]
- Pressel, S.; Bidartondo, M.I.; Ligrone, R.; Duckett, J.G. Fungal symbioses in bryophytes: New insights in the twenty first century. Phytotaxa 2010, 9, 238–253. [Google Scholar] [CrossRef]
- Davey, M.L.; Currah, R.S. Interactions between mosses (Bryophyta) and fungi. Botany 2006, 84, 1509–1519. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Schüßler, A. Glomus claroideum forms an arbuscular mycorrhiza-like symbiosis with the hornwort Anthoceros punctatus. Mycorrhiza 2000, 10, 15–21. [Google Scholar] [CrossRef]
- Russell, J.; Bulman, S. The liverwort Marchantia foliacea forms a specialized symbiosis with arbuscular mycorrhizal fungi in the genus Glomus. New Phytol. 2005, 165, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Ligrone, R.; Carafa, A.; Lumini, E.; Bianciotto, V.; Bonfante, P.; Duckett, J.G. Glomeromycotean associations in liverworts: A molecular, cellular, and taxonomic analysis. Am. J. Bot. 2007, 94, 1756–1777. [Google Scholar] [CrossRef] [Green Version]
- Liepiņa, L. Occurrence of fungal structures in bryophytes of the boreo-nemoral zone. Environ. Exp. Biol. 2012, 10, 35–40. [Google Scholar]
- Cottet, A.C.; Messuti, M.I. Identificación del tipo morfológico de micorriza arbuscular en Phaeoceros laevis (Anthocerotophyta). Bol. Soc. Argent. Bot. 2017, 52, 291–293. [Google Scholar] [CrossRef]
- Cottet, A.C.; Messuti, M.I. New evidence about the interactions between liverworts in the genus Symphyogyna (Pallaviciniaceae) and arbuscular mycorrhizal fungi. Symbiosis 2019, 79, 117–121. [Google Scholar] [CrossRef]
- Cottet, A.C.; Messuti, M.I. New record of arbuscular mycorrhizal fungi in Nothoceros fuegiensis (Dendrocerotaceae, Anthocerotophyta). Acta Bot. Mex. 2020, 127, 1–5. [Google Scholar] [CrossRef]
- Cottet, A.C.; Messuti, M.I. Nuevo registro de micorriza arbuscular en Asterella chilensis (Aytoniaceae, Marchantiophyta), Patagonia, Argentina. Bol. Soc. Argent. Bot. 2022, 57, 1–10. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Oberholzer, H.R.; Jansa, J.; Egli, S. Diversity and biogeography of arbuscular mycorrhizal fungi in agricultural soils. Biol. Fertil. Soils 2017, 53, 777–797. [Google Scholar] [CrossRef]
- Mhlanga, B.; Ercoli, L.; Piazza, G.; Thierfelder, C.; Pellegrino, E. Occurrence and diversity of arbuscular mycorrhizal fungi colonising off-season and in-season weeds and their relationship with maize yield under conservation agriculture. Biol. Fertil. Soils 2022, 58, 917–935. [Google Scholar] [CrossRef]
- Fernandez-Gnecco, G.A.; Smalla, K.; Maccario, L.; Sørensen, S.J.; Barbieri, P.A.; Consolo, V.F.; Covacevich, F.; Babin, D. Microbial community analysis of soils under different soybean cropping regimes in the Argentinean south-eastern Humid Pampas. FEMS Microbiol. Ecol. 2021, 97, fiab007. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gnecco, G.A.; Covacevich, F.; Consolo, V.F.; Behr, J.H.; Sommermann, L.; Moradtalab, N.; Maccario, L.; Sørensen, S.J.; Deubel, A.; Schellenberg, I.; et al. Effect of long-term agricultural management on the soil microbiota differs between seasons. Front. Soil Sci. 2022, 8, 837508. [Google Scholar] [CrossRef]
- Abarca, C.; Barrera, M.D.; Cabello, M.; Valdés, F.E.; Velázquez, M.S. Invasion of a xeric forest by an exotic tree species in Argentina: Impacts on the diversity of arbuscular mycorrhizal fungi and pre-existing mutualistic relationships. Acta Bot. Brasil. 2021, 35, 269–275. [Google Scholar] [CrossRef]
- Roesler, I.; Agostini, M.G. Inventario de los Vertebrados de la Reserva Natural Punta Lara, Provincia de Buenos Aires, Argentina; Aves Argentinas/Asociación Ornitológica del Plata: Buenos Aires, Argentina, 2012; pp. 1–169. [Google Scholar]
- Climate.org. Available online: https://es.climate-data.org/america-del-sur/argentina/buenos-aires/punta-lara-764839/ (accessed on 12 January 2023).
- Hurtado, M.; Giménez, J.; Cabral, M.; Da Silva, M.; Martínez, O.R.; Camillón, M.C.; Sánchez, C.A. Análisis Ambiental del Partido de La Plata: Aportes al Ordenamiento Territorial. Instituto de Geomorfología y Suelos. La Plata: Centro de Investigaciones de Suelos y Aguas de Uso Agropecuario, Municipalidad de La Plata, Consejo Federal de Inversiones. 2006. Available online: https://libros.unlp.edu.ar/index.php/unlp/catalog/view/275/257/824-1 (accessed on 12 January 2023).
- Frahm, J.P. Manual of Tropical Bryology, 3rd ed.; Tropical Bryology: Meckenheimer Allee, Germany, 2003; pp. 1–195. [Google Scholar]
- Churchill, S.P.; Linares, C.E.L. Prodromus Bryologiae Novo-Granatensis: Introducción a la Flora de Musgos de Colombia; Guadalupe Ltda: Santa Fe de Bogotá, Colombia, 1995; pp. 1–927. [Google Scholar]
- Hedenäs, L. Amblystegiaceae (Musci). Fl. Neotrop. Monogr. 2003, 89, 1–107. [Google Scholar]
- Pursell, R.A. Fissidentaceae. Fl. Neotrop. Monogr. 2007, 101, 1–279. [Google Scholar]
- Cottet, A.C.; Scervino, J.M.; Messuti, M.I. An improved staining protocol for the assessment of arbuscular mycorrhizal in bryophytes. Bol. Soc. Argent. Bot. 2018, 53, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gerdemann, J.W.; Nicolson, T.H. Spores of Mycorrhizal Endogone Species Extracted from Soil by Wet Sieving and Decanting. Trans. Brit. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Błaszkowski, J. Glomeromycota; W. Szafer Institute of Botany, Polish Academy of Sciences: Warszawa, Poland, 2012. [Google Scholar]
- Oehl, F.; Sanchez-Castro, I.; de Sousa, N.M.F.; Silva, G.; Palenzuela, J. Dominikia bernensis, a new arbuscular mycorrhizal fungus from a swiss no-till farming site, and D. aurea, D. compressa and D. indica, three new combinations in Dominikia. Nova Hedwig. 2015, 101, 65–76. [Google Scholar] [CrossRef]
- Weining, S.; Langridge, P. Identification and Mapping of Polymorphisms in Cereals Based on the Polymerase Chain Reaction. Theor. Appl. Genet. 1991, 82, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Kjøller, R.; Rosendahl, S. Detection of arbuscular mycorrhizal fungi (Glomales) in roots by nested PCR and SSCP (Single Stranded Conformation Polymorphism). Plant Soil 2012, 226, 189–196. [Google Scholar] [CrossRef]
- Covacevich, F.; Hernández Guijarro, K.; Crespo, E.M.; Lumini, E.; Rivero Mega, M.S.; Lugo, M.A. Arbuscular Mycorrhizal Fungi from Argentinean Highland Puna Soils Unveiled by Propagule Multiplication. Plants 2021, 10, 1803. [Google Scholar] [CrossRef]
- Benbouza, H.; Jacquemin, J.M.; Baudoin, J.P.; Mergeai, G. Optimization of a reliable, fast, cheap and sensitive silver staining method to detect SSR markers in polyacrylamide gels. Biotechnol. Agron. Soc. Environ. 2006, 10, 77–81. [Google Scholar]
- Carleton, T.J.; Read, D.J. Ectomycorrhizas and nutrient transfer in conifer–feather moss ecosystems. Can. J. Bot. 1991, 69, 778–785. [Google Scholar] [CrossRef]
- Fonseca, H.M.; Berbara, R.L. Does Lunularia cruciata form symbiotic relationships with either Glomus proliferum or G. intraradices? Mycol. Res. 2008, 112, 1063–1068. [Google Scholar] [CrossRef]
- Ptaszyńska, A.; Mułenko, W.; Żarnowiec, J. Bryophytes microniches inhabited by microfungi. Ann. Univ. Mariae Curie-Skłodowska 2009, 64, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Klymiuk, A.A.; Taylor, T.N.; Taylor, E.L.; Krings, M. Paleomycology of the Princeton Chert II. Dark-septate fungi in the aquatic angiosperm Eorhiza arnoldii indicate a diverse assemblage of root-colonizing fungi during the Eocene. Mycologia 2013, 105, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Muthuraja, R.; Muthukumar, T.; Sathiyadash, K.; Uma, E.; Priyadharsini, P. Arbuscular mycorrhizal (AM) and dark septate endophyte (DSE) fungal association in lycophytes and ferns of the Kolli Hills, Eastern Ghats, Southern India. Am. Fern J. 2014, 104, 67–102. [Google Scholar] [CrossRef]
- Thangavelu, M.; Raji, M. Arbuscular mycorrhizal and dark septate endophyte fungal associations in Asparagus. Turk. J. Bot. 2016, 40, 662–675. [Google Scholar] [CrossRef]
- Davey, M.L.; Nybakken, L.; Kauserud, H.; Ohlson, M. Fungal biomass associated with the phyllosphere of bryophytes and vascular plants. Mycol. Res. 2009, 113, 1254–1260. [Google Scholar] [CrossRef] [PubMed]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Arnold, A.E. Community analysis reveals close affinities between endophytic and endolichenic fungi in mosses and lichens. Microb. Ecol. 2010, 60, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Kottke, I.; Nebel, M. The evolution of mycorrhiza-like associations in liverworts: An update. New Phytol. 2005, 167, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J.; Ryszka, P.; Kozłowska, A.N.N.A. Dominikia litorea, a new species in the Glomeromycotina, and biogeographic distribution of Dominikia. Phytotaxa 2018, 338, 241–254. [Google Scholar] [CrossRef]
- Cofré, M.N.; Soteras, F.; de Rosario Iglesias, M.; Velázquez, M.; Abarca, C.; Risio, L.; Ontivero, E.; Cabello, M.N.; Domínguez, L.S.; Lugo, M.A. Biodiversity of arbuscular mycorrhizal fungi in south america: A review. In Mycorrhizal fungi in South America; Pagano, M.C., Lugo, M.A., Eds.; Springer International Publishing: New York, NY, USA, 2019; pp. 49–72. [Google Scholar]
- Tedersoo, L. The Global Soil Mycobiome consortium dataset for boosting fungal diversity research. Fungal Divers. 2021, 111, 573–588. [Google Scholar] [CrossRef]
- Rosendahl, S.; Stukenbrock, E. Community structure of arbuscular mycorrhizal fungi in undisturbed vegetation revealed by analyses of LSU rDNA sequences. Molec. Ecol. 2004, 13, 3179–3186. [Google Scholar] [CrossRef]
- Thougnon Islas, A.J.; Hernandez Guijarro, K.; Eyherabide, M.; Sainz Rozas, H.R.; Echeverría, H.E.; Covacevich, F. Can soil properties and agricultural land use affect arbuscular mycorrhizal fungal communities indigenous from the Argentinean Pampas soils? Appl. Soil Ecol. 2006, 101, 47–56. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| InterHy–IntraHy | hc | ves | s | Sp | DSE | |
|---|---|---|---|---|---|---|
| Dumortiera hirsuta | x | x | x | x | - | x |
| Cyclodictyon albicans | x | x | x | x | - | x |
| Fissidens elegans | - | - | - | - | - | x |
| Hygroamblystegium varium | x | x | - | x | - | x |
| Isopterygium tenerum | - | - | - | x | x | x |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdés, F.E.; Peralta, D.F.; Velázquez, M.S.; Covacevich, F.; Becerra, A.G.; Cabello, M.N. On the Occurrence of Arbuscular Mycorrhizal Fungi in a Bryophyte Community of Punta Lara Natural Reserve, Buenos Aires, Argentina. Diversity 2023, 15, 442. https://doi.org/10.3390/d15030442
Valdés FE, Peralta DF, Velázquez MS, Covacevich F, Becerra AG, Cabello MN. On the Occurrence of Arbuscular Mycorrhizal Fungi in a Bryophyte Community of Punta Lara Natural Reserve, Buenos Aires, Argentina. Diversity. 2023; 15(3):442. https://doi.org/10.3390/d15030442
Chicago/Turabian StyleValdés, Fabricio Emanuel, Denilson Fernandes Peralta, María Silvana Velázquez, Fernanda Covacevich, Alejandra Gabriela Becerra, and Marta Noemí Cabello. 2023. "On the Occurrence of Arbuscular Mycorrhizal Fungi in a Bryophyte Community of Punta Lara Natural Reserve, Buenos Aires, Argentina" Diversity 15, no. 3: 442. https://doi.org/10.3390/d15030442
APA StyleValdés, F. E., Peralta, D. F., Velázquez, M. S., Covacevich, F., Becerra, A. G., & Cabello, M. N. (2023). On the Occurrence of Arbuscular Mycorrhizal Fungi in a Bryophyte Community of Punta Lara Natural Reserve, Buenos Aires, Argentina. Diversity, 15(3), 442. https://doi.org/10.3390/d15030442