


Who Lives in the Hot Heart of the Cold Sea? A New Species of Provanna (Caenogastropoda: Provannidae) from the Hydrothermal Zone of Piip Volcano, Northwestern Pacific

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Sample Processing
2.3. Morphological Studies
2.4. Molecular Studies
2.5. Data Analysis and Visualization
3. Results
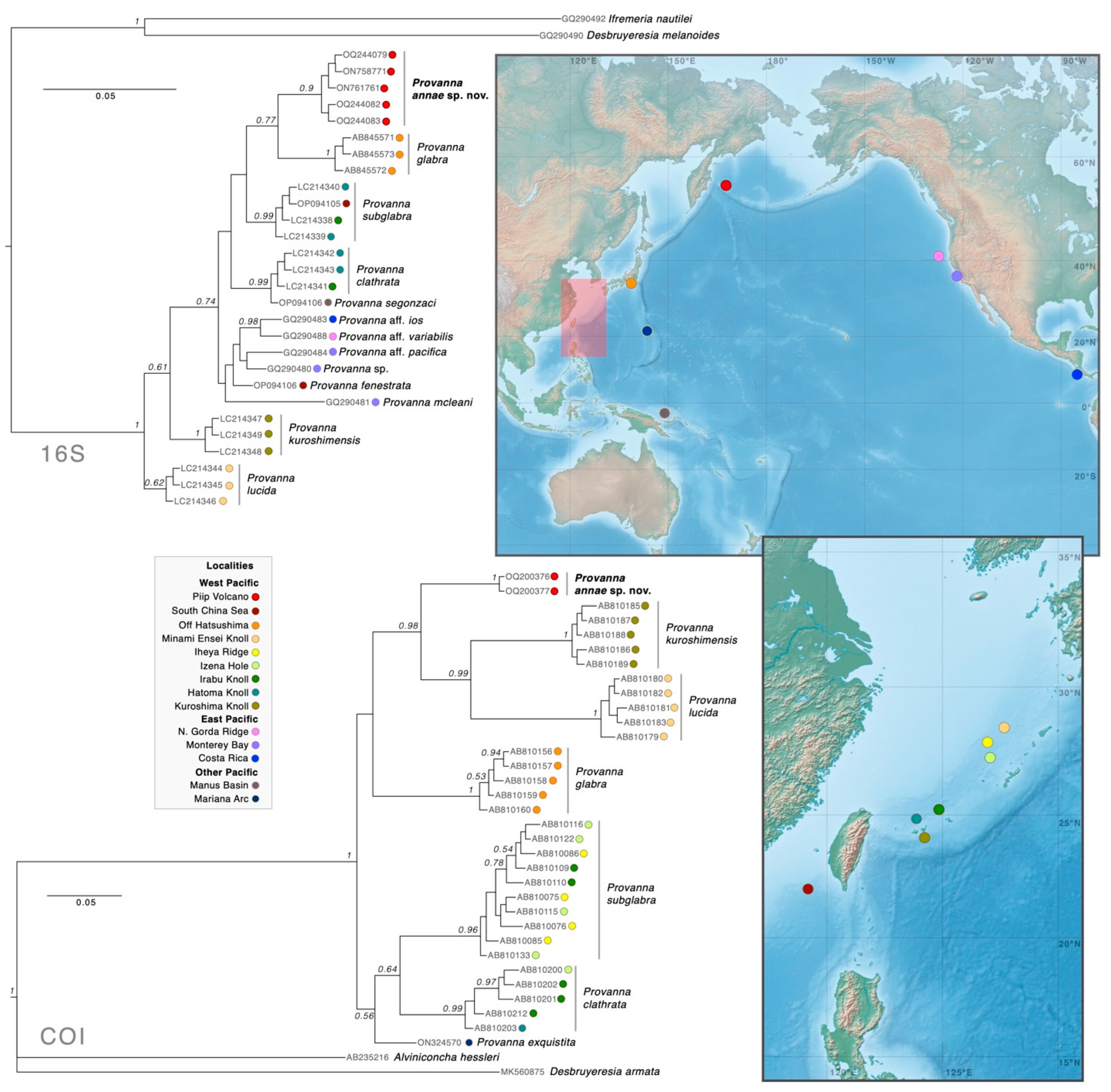
4. Discussion
4.1. Biogeography
4.2. Feeding of Provanna
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Golikov, A.N. Arctic Ocean Gastropod Prosobranhcs. In The Arctic Seas: Climatology, Oceanography, Geology and Biology; Herman, Y., Ed.; Springer: New York, NY, USA, 1989; pp. 325–340. [Google Scholar]
- Nekhaev, I.O.; Krol, E.N. Hidden under Ice and Mud: Diversity of Shell-Bearing Microgastropods in the Eastern Arctic Seas. Syst. Biodivers 2020, 18, 794–809. [Google Scholar] [CrossRef]
- Karaseva, N.P.; Rimskaya-Korsakova, N.N.; Smirnov, R.V.; Udalov, A.A.; Mokievsky, V.O.; Gantsevich, M.M.; Malakhov, V.V. Distribution of Gutless Siboglinid Worms (Annelida, Siboglinidae) in Russian Arctic Seas in Relation to Gas Potential. Diversity 2022, 14, 1061. [Google Scholar] [CrossRef]
- Krol, E.N.; Nekhaev, I.O. Redescription of Leptogyra Bujnitzkii (Gorbunov, 1946) Comb. Nov., the First Representative of the Gastropod Subclass Neomphaliones from the High Arctic. Zootaxa 2020, 4759, 446–450. [Google Scholar] [CrossRef]
- Rybakova, E.; Krylova, E.; Mordukhovich, V.; Galkin, S.; Alalykina, I.; Smirnov, I.; Sanamyan, N.; Nekhaev, I.; Vinogradov, G.; Shilov, V.; et al. Methane Seep Communities on the Koryak Slope in the Bering Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2022, 206, 105203. [Google Scholar] [CrossRef]
- Galkin, S.V.; Mordukhovich, V.V.; Krylova, E.M.; Denisov, V.A.; Malyutin, A.N.; Mikhailik, P.E.; Polonik, N.S.; Sanamyan, N.P.; Shilov, V.A.; Ivin, V.V.; et al. Comprehensive Research of Ecosystems of Hydrothermal Vents and Cold Seeps in the Bering Sea (Cruise 82 of the R/V Akademik M.A. Lavrentyev). Oceanology 2019, 59, 618–621. [Google Scholar] [CrossRef]
- Nekhaev, I.O.; Chaban, E.M.; Kantor, Y.I.; Kuchsh, D.A.; Matveeva, K.; Rybakova, E. Shell-Bearing Gastropoda from the Methane Seeps and Hydrothermal Vents of the Bering Sea: A Preliminary Description. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2022, 204, 105164. [Google Scholar] [CrossRef]
- Rybakova, E.; Krylova, E.; Mordukhovich, V.; Galkin, S.; Alalykina, I.; Sanamyan, N.; Nekhaev, I.; Vinogradov, G.; Shilov, V.; Pakhnevich, A.; et al. Mega- and Macrofauna of the Hydrothermally Active Submarine Piip Volcano (the Southwestern Bering Sea). Deep. Sea Res. Part II Top. Stud. Oceanogr. 2023, 208, 105268. [Google Scholar] [CrossRef]
- Chaban, E.M.; Schepetov, D.M.; Ekimova, I.A.; Nekhaev, I.O.; Chernyshev, A.V. The First Record of the Family Parvaplustridae (Gastropoda, Heterobranchia) from Hydrothermal Vent Fields of the Piip Volcano, Bering Sea with the Description of a New Species of the Genus Parvaplustrum. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2022, 202, 105135. [Google Scholar] [CrossRef]
- Nekhaev, I.O. Skenea Profunda (Vetigastropoda: Skeneidae) in the Central Arctic. Ruthenica Russ. Malacol. J. 2022, 32, 105–109. [Google Scholar] [CrossRef]
- Nekhaev, I.O. Taxonomic Review of the Genus Boreocingula (Gastropoda: Rissoidae) in the Arctic and Cold Temperate Waters. Polar Biol. 2019, 42, 889–905. [Google Scholar] [CrossRef]
- Sasaki, T.; Ogura, T.; Watanabe, H.K.; Fujikura, K. Four New Species of Provanna (Gastropoda: Provannidae) from Vents and a Seep off Nansei-Shoto Area, Southwestern Japan. Venus J. Malacol. Soc. Jpn. 2016, 74, 1–17. [Google Scholar] [CrossRef]
- Criscione, F.; Ponder, W.F. A Phylogenetic Analysis of Rissooidean and Cingulopsoidean Families (Gastropoda: Caenogastropoda). Mol. Phylogenet. Evol. 2013, 66, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.T.; Taylor, J.D.; Glover, E.A. Molecular Phylogeny of the Lucinoidea (Bivalvia): Non-Monophyly and Separate Acquisition of Bacterial Chemosymbiosis. J. Molluscan Stud. 2004, 70, 187–202. [Google Scholar] [CrossRef]
- Golosova, O.; Henderson, R.; Vaskin, Y.; Gabrielian, A.; Grekhov, G.; Nagarajan, V.; Oler, A.J.; Quiñones, M.; Hurt, D.; Fursov, M.; et al. Unipro UGENE NGS Pipelines and Components for Variant Calling, RNA-Seq and ChIP-Seq Data Analyses. PeerJ 2014, 2, e644. [Google Scholar] [CrossRef]
- Ogura, T.; Watanabe, H.K.; Chen, C.; Sasaki, T.; Kojima, S.; Ishibashi, J.I.; Fujikura, K. Population History of Deep-Sea Vent and Seep Provanna Snails (Mollusca: Abyssochrysoidea) in the Northwestern Pacific. PeerJ 2018, 6, e5673. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.B.; Warén, A.; Lee, R.W.; Kano, Y.; Kaim, A.; Davis, A.; Strong, E.E.; Vrijenhoek, R.C. Rubyspira, New Genus and Two New Species of Bone-Eating Deep-Sea Snails with Ancient Habits. Biol. Bull. 2010, 219, 166–177. [Google Scholar] [CrossRef]
- Schliep, K.; Potts, A.J.; Morrison, D.A.; Grimm, G.W. Intertwining Phylogenetic Trees and Networks. Methods Ecol. Evol. 2017, 8, 1212–1220. [Google Scholar] [CrossRef]
- Hohna, S.; Landis, M.J.; Heath, T.A.; Boussau, B.; Lartillot, N.; Moore, B.R.; Huelsenbeck, J.P.; Ronquist, F. RevBayes: Bayesian Phylogenetic Inference Using Graphical Models and an Interactive Model-Specification Language. Syst. Biol. 2016, 65, 726–736. [Google Scholar] [CrossRef]
- Warén, A.; Bouchet, P. Four New Species of Provanna Dall (Prosobranchia, Cerithiacea?) From East Pacific Hydrothermal Sites. Zool. Scr. 1986, 15, 157–164. [Google Scholar] [CrossRef]
- Warén, A.; Ponder, W.F. New Species, Anatomy, and Systematic Position of the Hydrothermal Vent and Hydrocarbon Seep Gastropod Family Provannidae Fam.n. (Caenogastropoda). Zool. Scr. 1991, 20, 27–56. [Google Scholar] [CrossRef]
- Kantor, Y.; Zvonareva, S.; Krylova, E. New species of the genus Astyris (Gastropoda: Neogastropoda: Buccinoidea: Columbellidae) from the vicinity of hydrothermal vents of the Piip Volcano, southwestern Bering Sea. Zootaxa 2023, 5263, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Ekimova, I.; Valdés, Á.; Stanovova, M.; Mikhlina, A.; Antokhina, T.; Neretina, T.; Chichvarkhina, O.; Schepetov, D. Connected across the Ocean: Taxonomy and Biogeography of Deep-Water Nudibranchia from the Northwest Pacific Reveal Trans-Pacific Links and Two Undescribed Species. Org. Divers. Evol. 2021, 21, 753–782. [Google Scholar] [CrossRef]
- Sirenko, B.I.; Merkuliev, A.V.; Krol, E.N.; Danilin, D.; Nekhaev, I.O. To a Species Composition of Subfamily Moelleriinae Hickman et McLean, 1990 (Mollusca: Gastropoda: Colloniidae) in the Northwestern Pacific. Ruthenica 2018, 28, 19–25. [Google Scholar] [CrossRef]
- Zhang, J.; Yurchenko, O.; Lutaenko, K.A.; Kalachev, A.; Nekhaev, I.O.; Aguilar, R.; Zhan, Z.; Ogburn, M.B. A Tale of Two Soft-Shell Clams: An Integrative Taxonomic Analysis Confirms Mya Japonica as a Valid Species Distinct from Mya Arenaria (Bivalvia: Myidae). Zool. J. Linn. Soc. 2018, 184, 605–622. [Google Scholar] [CrossRef]
- Hasegawa, K. A Review of Bathyal Rissoidae in the Sea of Japan and Adjacent Waters (Gastropoda: Rissooidea). Natl. Mus. Nat. Sci. Monogr. 2014, 44, 75–148. [Google Scholar]
- Karaseva, N.P.; Rimskaya-Korsakova, N.N.; Ekimova, I.A.; Kokarev, V.N.; Simakov, M.I.; Gantsevich, M.M.; Malakhov, V.V. The First Discovery of Pogonophora (Annelida, Siboglinidae) in the East Siberian Sea Coincides with the Areas of Methane Seeps. Dokl. Biol. Sci. 2021, 501, 187–191. [Google Scholar] [CrossRef]
- Edmonds, H.N.; Michael, P.J.; Baker, E.T.; Connelly, D.P.; Snow, J.E.; Langmuir, C.H.; Dick, H.J.B.; Mühe, R.; German, C.R.; Graham, D.W. Discovery of Abundant Hydrothermal Venting on the Ultraslow-Spreading Gakkel Ridge in the Arctic Ocean. Nature 2003, 421, 252–256. [Google Scholar] [CrossRef]
- Stewart, F.J.; Newton, I.L.G.; Cavanaugh, C.M. Chemosynthetic Endosymbioses: Adaptations to Oxic-Anoxic Interfaces. Trends Microbiol. 2005, 13, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.; Cary, S.; Hessler, R.; Vetter, R.; Felbeck, H.; Ohta, S.; Childress, J. Chemoautotrophic Symbiosis in a Hydrothermal Vent Gastropod. Biol. Bull. 1988, 174, 373–378. [Google Scholar] [CrossRef]
- Windoffer, R.; Giere, O. Symbiosis of the Hydrothermal Vent Gastropod Ifremeria Nautilei (Provannidae) with Endobacteria—Structural Analyses and Ecological Considerations. Biol. Bull. 1997, 193, 381–392. [Google Scholar] [CrossRef]
- Warén, A.; Bouchet, P. New Records, Species, Genera, and a New Family of Gastropods from Hydrothermal Vents and Hydrocarbon Seeps. Zool. Scr. 1993, 22, 1–90. [Google Scholar] [CrossRef]
- Warén, A.; Bouchet, P. New Gastropods from Deep-Sea Hydrocarbon Seeps off West Africa. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 2326–2349. [Google Scholar] [CrossRef]
- Ball, A.D.; Purdy, K.J.; Glover, E.A.; Taylor, J.D. Ctenidial Structure and Three Bacterial Symbiont Morphotypes in Anodontia (Euanodontia) Ovum (Reeve, 1850) from the Great Barrier Reef, Australia (Bivalvia: Lucinidae). J. Molluscan Stud. 2009, 75, 175–185. [Google Scholar] [CrossRef]
- Bates, A.E. Persistence, Morphology, and Nutritional State of a Gastropod Hosted Bacterial Symbiosis in Different Levels of Hydrothermal Vent Flux. Mar. Biol. 2007, 152, 557–568. [Google Scholar] [CrossRef]
- de Burgh, M.E.; Singla, C.L. Bacterial Colonization and Endocytosis on the Gill of a New Limpet Species from a Hydrothermal Vent. Mar. Biol. 1984, 84, 1–6. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | N | E | Depth, m | Date | Locality and Sampled Substrate |
|---|---|---|---|---|---|
| LV75-4/1 | 55°24.99′ | 167°16.494′ | 390 | 16 June 2016 | Northern summit, stones with bacterial mats |
| LV75-5/1 | 55°24.99′ | 167°16.494′ | 387 | 17 June 2016 | Northern summit, stones with bacterial mats |
| LV75-10/1 | 55°24.996′ | 167°16.488′ | 402 | 20 June 2016 | Northern summit, stones with bacterial mats |
| LV82-2/7 | 55°24.948′ | 167°16.596′ | 400 | 13 June 2018 | Northern summit, bacterial mat |
| LV82-3/1 | 55°22.938′ | 167°15.666′ | 469 | 14 June 2018 | Northern summit, Calyptogena pacifica with sediment |
| LV82-8/3 | 55°22.902′ | 167°15.672′ | 471 | 17 June 2018 | Southern summit, Calyptogena pacifica and stones with sediment |
| LV82-8/4 | 55°22.908′ | 167°15.684′ | 472 | 17 June 2018 | Southern summit, sediments from hydrothermal edifice |
| Gene | Initial Denaturation | Cycle | Final Annealing | |||
|---|---|---|---|---|---|---|
| Number of Cycles | Denaturation | Annealing | Elongation | |||
| COI | 94 °C/120 s | 30 | 94 °C/30 s | 45 °C/30 s | 72 °C/30 s | 72 °C/40 s |
| 16S | 94 °C/300 s | 40 | 94 °C/30 s | 52 °C/30 s | 72 °C/60 s | 72 °C/600 s |
| 28S | 95 °C/180 s | 35 | 94 °C/20 s | 52 °C/60 s | 72 °C/120 s | 72 °C/600 s |
| Station | Individual No. | Notes | Genbank Accession Number | ||
|---|---|---|---|---|---|
| COI | 16S | 28S | |||
| LV82-8/3 | 3 | Paratype | - | OQ244079 | - |
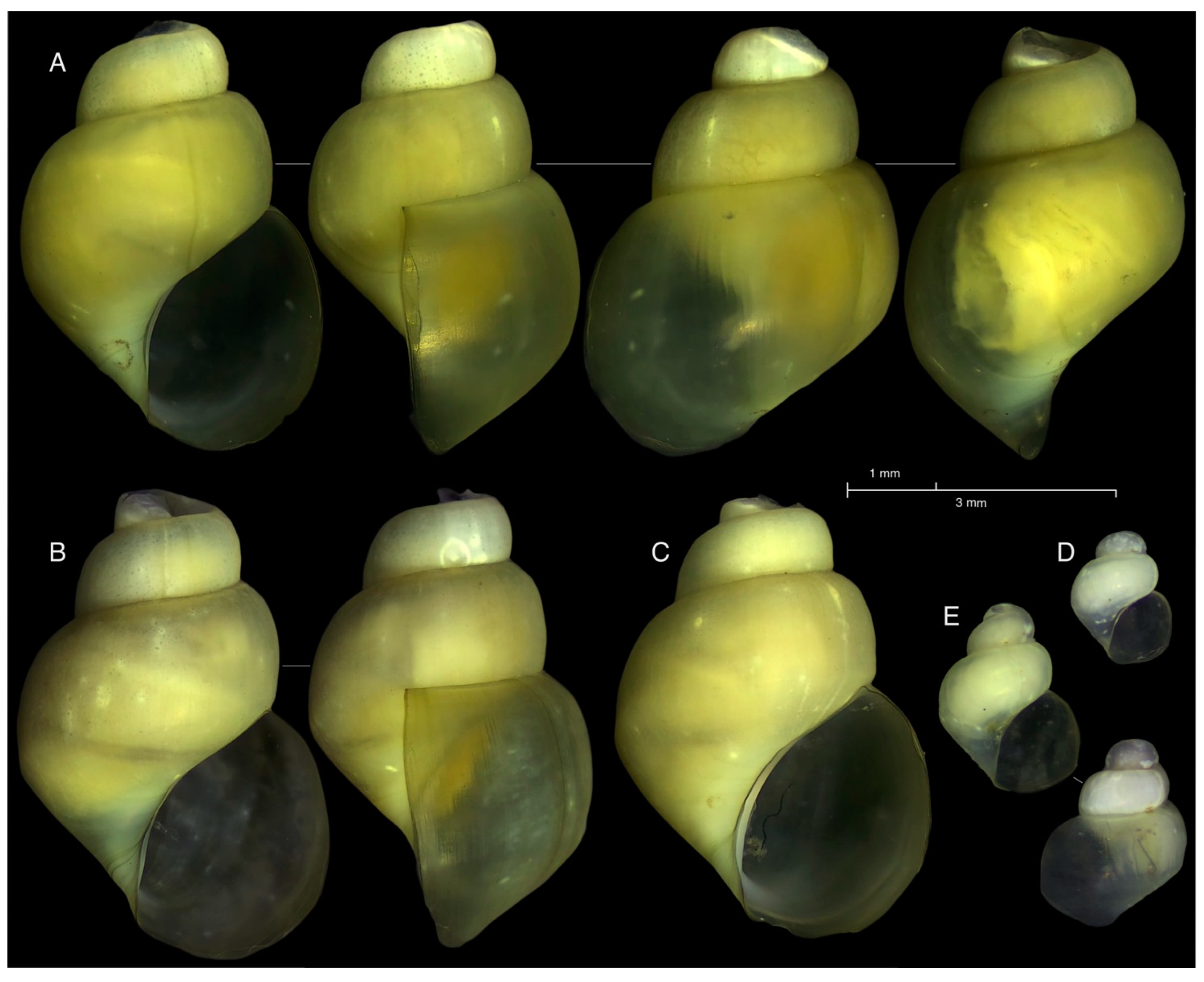
| 4 | Paratype, Figure 2B | OQ200377 | ON758771 | OQ690022 | |
| 5 | Paratype, Figure 2C | OQ200376 | ON761761 | OQ690023 | |
| 10 | Holotype, Figure 2A | - | OQ244082 | - | |
| 11 | Paratype, Figure 5A,B | - | OQ244083 | OQ690024 | |
| LV75-10/1 | 1 | - | - | - | OQ690019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekhaev, I.O. Who Lives in the Hot Heart of the Cold Sea? A New Species of Provanna (Caenogastropoda: Provannidae) from the Hydrothermal Zone of Piip Volcano, Northwestern Pacific. Diversity 2023, 15, 581. https://doi.org/10.3390/d15040581
Nekhaev IO. Who Lives in the Hot Heart of the Cold Sea? A New Species of Provanna (Caenogastropoda: Provannidae) from the Hydrothermal Zone of Piip Volcano, Northwestern Pacific. Diversity. 2023; 15(4):581. https://doi.org/10.3390/d15040581
Chicago/Turabian StyleNekhaev, Ivan O. 2023. "Who Lives in the Hot Heart of the Cold Sea? A New Species of Provanna (Caenogastropoda: Provannidae) from the Hydrothermal Zone of Piip Volcano, Northwestern Pacific" Diversity 15, no. 4: 581. https://doi.org/10.3390/d15040581
APA StyleNekhaev, I. O. (2023). Who Lives in the Hot Heart of the Cold Sea? A New Species of Provanna (Caenogastropoda: Provannidae) from the Hydrothermal Zone of Piip Volcano, Northwestern Pacific. Diversity, 15(4), 581. https://doi.org/10.3390/d15040581