
Breeding Population and Nesting Habitat of Skuas in the Harmony Point Antarctic Specially Protected Area



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
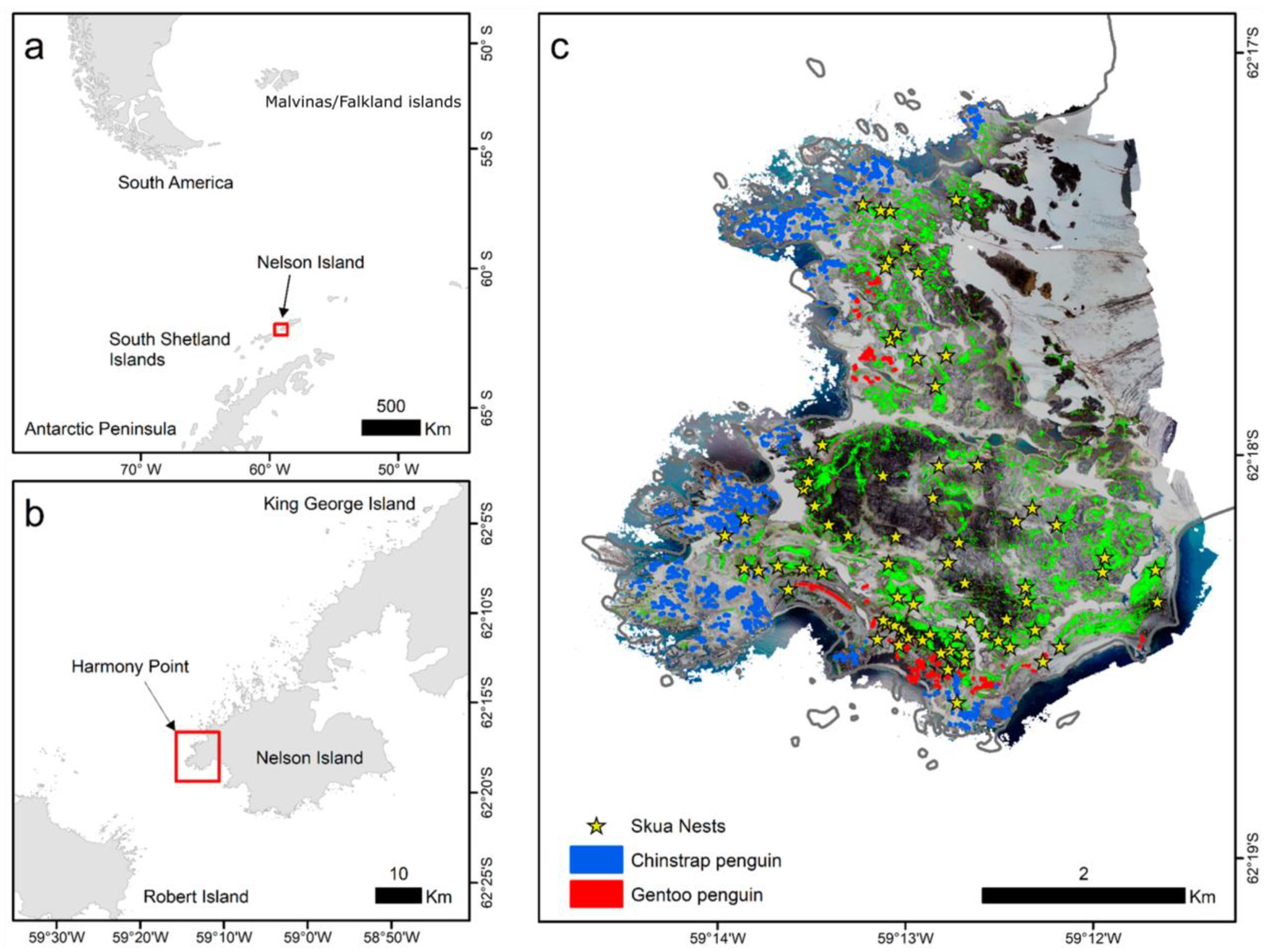
2.1. Nest Mapping
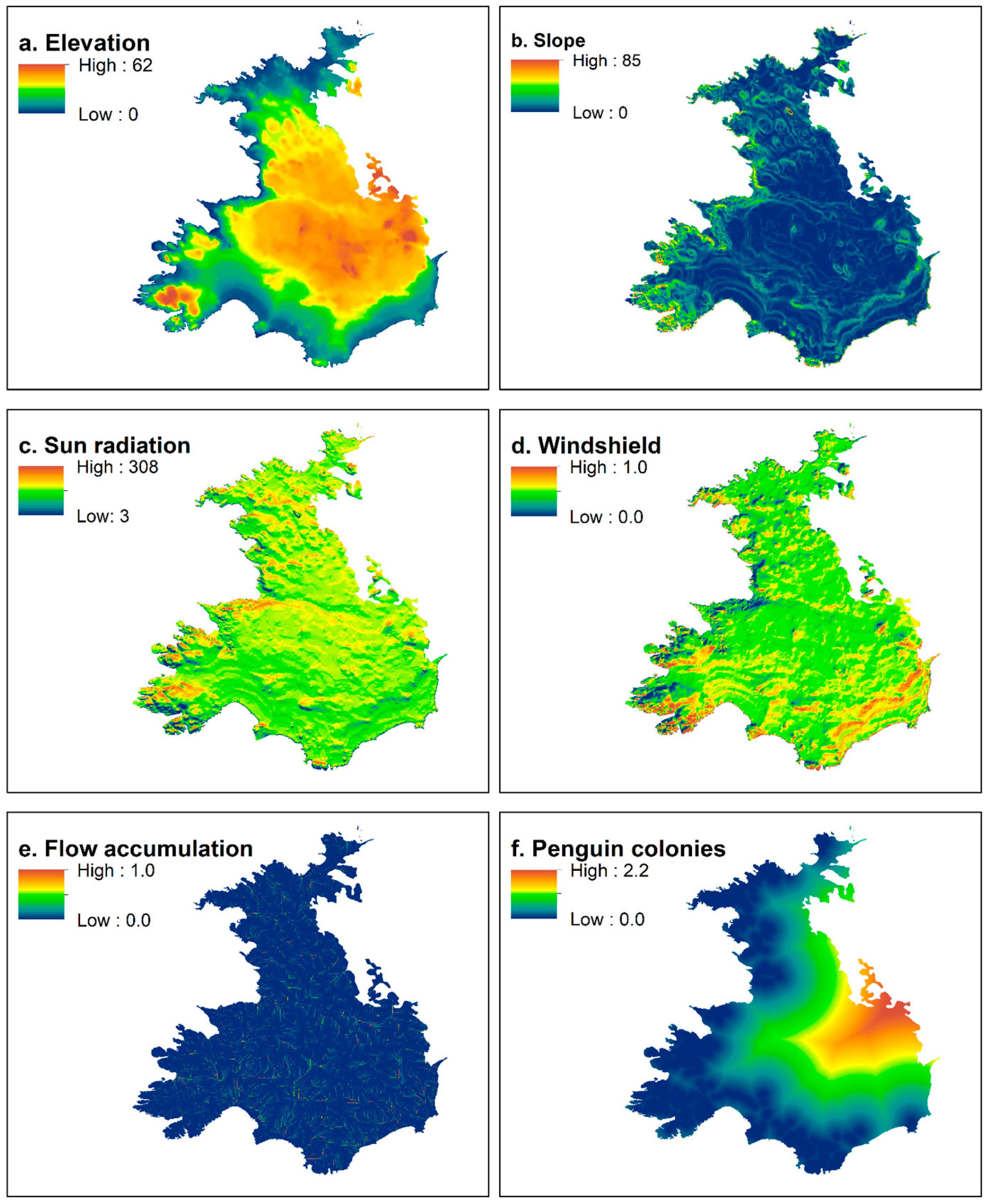
2.2. Nest Habitat
2.3. Nesting Habitat Suitability
3. Results
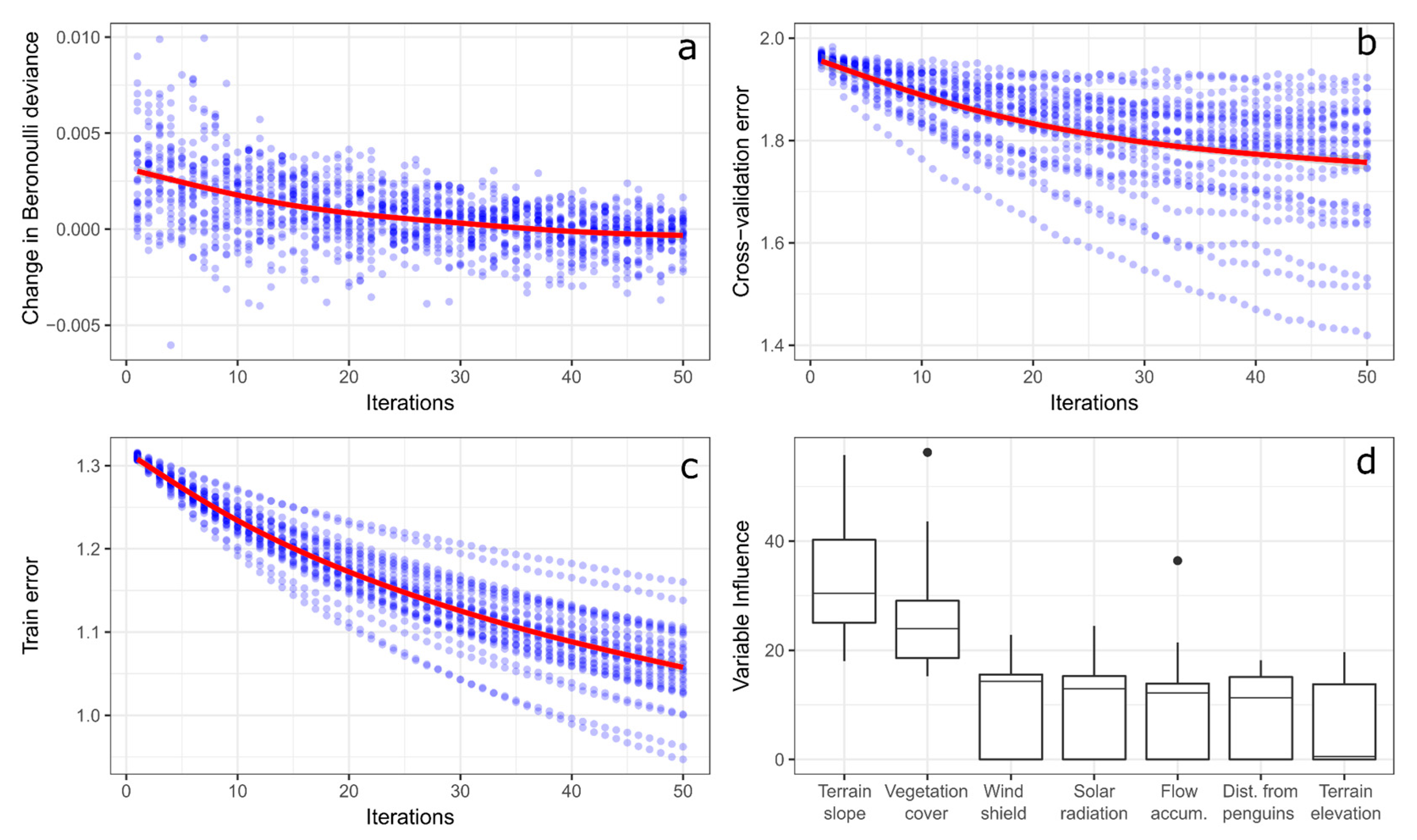
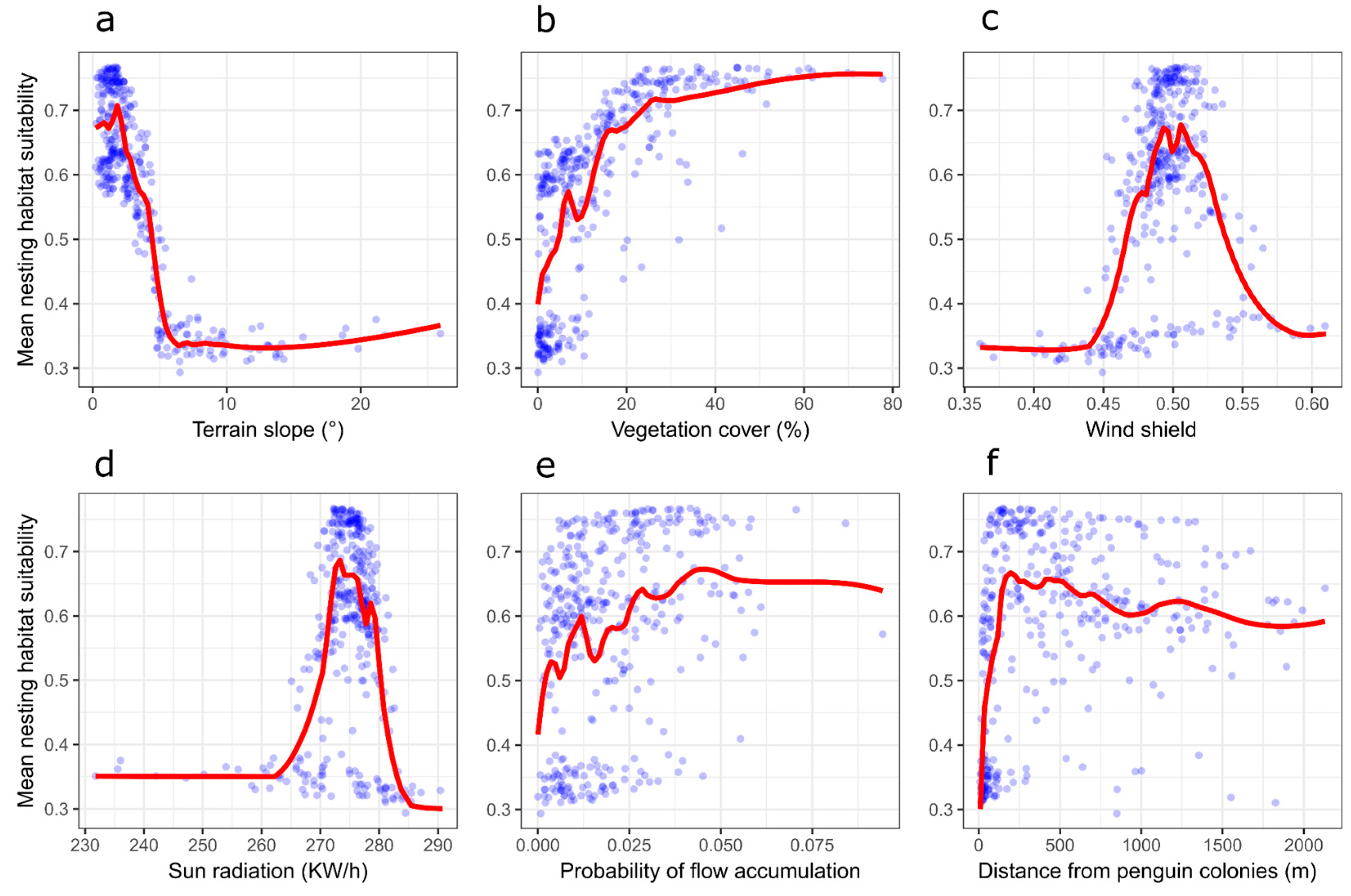
3.1. Nest Habitat Model Performance and Variable Contribution
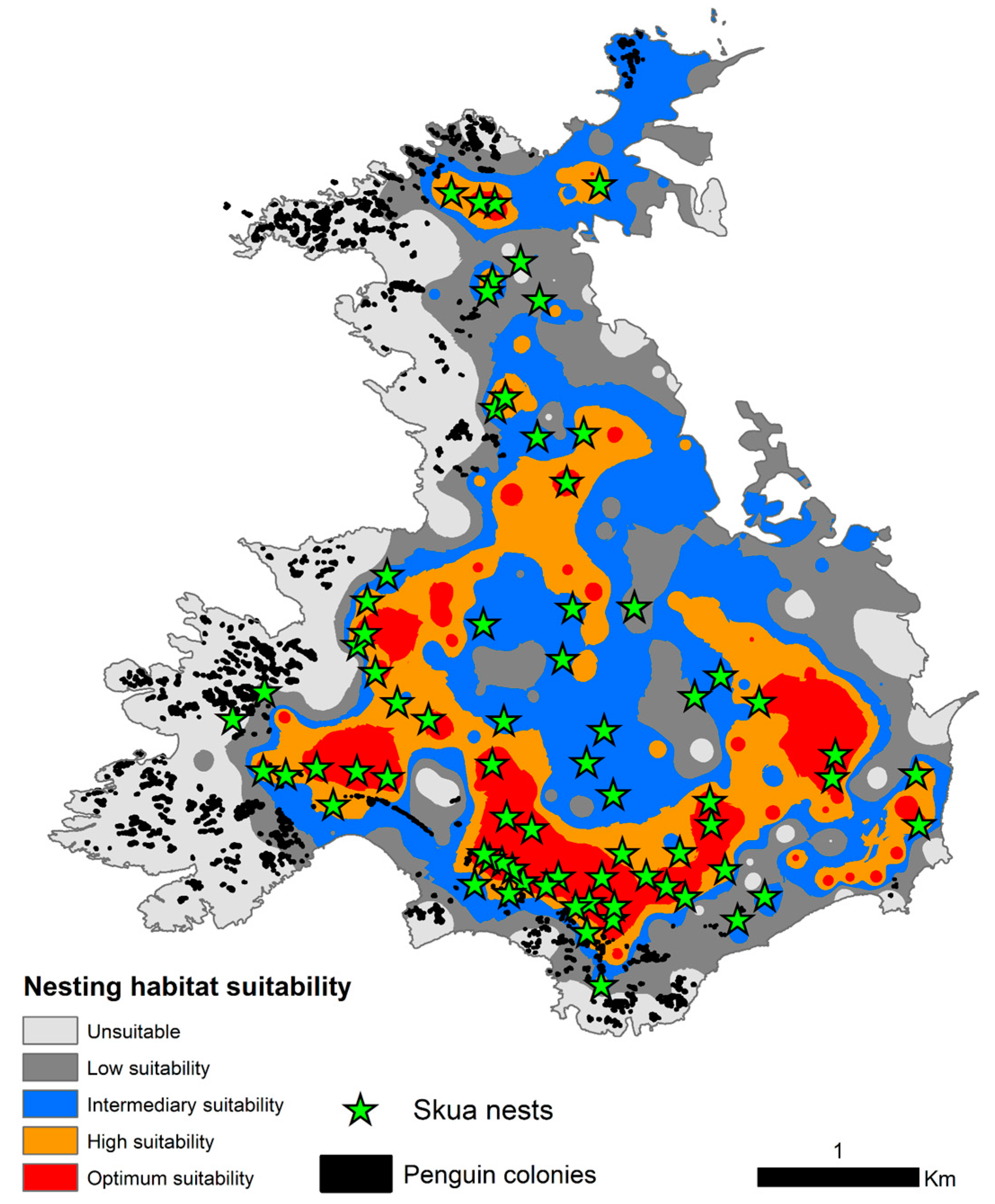
3.2. Nesting Habitat
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coetzee, B.W.T.; Convey, P.; Chown, S.L. Expanding the Protected Area Network in Antarctica Is Urgent and Readily Achievable. Conserv. Lett. 2017, 10, 670–680. [Google Scholar] [CrossRef]
- Wauchope, H.S.; Shaw, J.D.; Terauds, A. A Snapshot of Biodiversity Protection in Antarctica. Nat. Commun. 2019, 10, 946. [Google Scholar] [CrossRef]
- Harris, C.M.; Lorenz, K.; Fishpool, L.D.C.; Lascelles, B.; Cooper, J.; Coria, N.R.; Croxall, J.P.; Emmerson, L.M.; Fraser, W.R.; Fijn, R.C.; et al. Important Bird Areas in Antarctica 2015; BirdLife International and Environmental Research & Assessment Ltd.: Cambridge, UK, 2015. [Google Scholar]
- Lee, J.R.; Raymond, B.; Bracegirdle, T.J.; Chadès, I.; Fuller, R.A.; Shaw, J.D.; Terauds, A. Climate Change Drives Expansion of Antarctic Ice-Free Habitat. Nature 2017, 547, 49–54. [Google Scholar] [CrossRef]
- Brooks, S.T.; Jabour, J.; van den Hoff, J.; Bergstrom, D.M. Our Footprint on Antarctica Competes with Nature for Rare Ice-Free Land. Nat. Sustain. 2019, 2, 185–190. [Google Scholar] [CrossRef]
- Kennicutt, M.C.; Chown, S.L.; Cassano, J.J.; Liggett, D.; Peck, L.S.; Massom, R.; Rintoul, S.R.; Storey, J.; Vaughan, D.G.; Wilson, T.J.; et al. A Roadmap for Antarctic and Southern Ocean Science for the next Two Decades and Beyond. Antarct. Sci. 2015, 27, 3–18. [Google Scholar] [CrossRef]
- Grémillet, D.; Boulinier, T. Spatial Ecology and Conservation of Seabirds Facing Global Climate Change: A Review. Mar. Ecol. Prog. Ser. 2009, 391, 121–137. [Google Scholar] [CrossRef]
- Pereira, J.M.; Paiva, V.H.; Krüger, L.; Votier, S.C. Tracking Seabirds for Conservation and Marine Spatial Planning. In Seabird Biodiversity and Human Activities; Ramos, J.A., Pereira, L., Eds.; CRC Press: Boca Raton, FL, USA, 2022; pp. 59–74. ISBN 9781003047520. [Google Scholar]
- Krüger, L. Identifying and Establishing Marine Protected Areas Worldwide. In Seabird Biodiversity and Human Activities; CRC Press: Boca Raton, FL, USA, 2022; pp. 243–257. [Google Scholar]
- Higgins, P.J.; Davies, S.J. Handbook of Australian, New Zealand and Antarctic Birds; Oxford University Press: Melbourne, Australia, 1996. [Google Scholar]
- Carneiro, A.P.B.; Polito, M.J.; Sander, M.; Trivelpiece, W.Z. Abundance and Spatial Distribution of Sympatrically Breeding Catharacta Spp. (Skuas) in Admiralty Bay, King George Island, Antarctica. Polar Biol. 2010, 33, 673–682. [Google Scholar] [CrossRef]
- Ibañez, A.E.; Graña Grilli, M.; Figueroa, A.; Pari, M.; Montalti, D. Declining Health Status of Brown Skua (Stercorarius Antarcticus Lonnbergi) Parents and Their Offspring during Chick Development. Polar Biol. 2018, 41, 193–200. [Google Scholar] [CrossRef]
- Hahn, S.; Peter, H.-U. Feeding Territoriality and the Reproductive Consequences in Brown Skuas Catharacta Antarctica Lonnbergi. Polar Biol. 2003, 26, 552–559. [Google Scholar] [CrossRef]
- Pezzo, F.; Olmastroni, S.; Corsolini, S.; Focardi, S. Factors Affecting the Breeding Success of the South Polar Skua Catharacta Maccormicki at Edmonson Point, Victoria Land, Antarctica. Polar Biol. 2001, 24, 389–393. [Google Scholar] [CrossRef]
- Salzman, A.G. The Selective Importance of Heat Stress in Gull Nest Location. Ecology 1982, 63, 742–751. [Google Scholar] [CrossRef]
- Olin, A.; Dück, L.; Berglund, P.; Karlsson, E.; Bohm, M.; Olsson, O.; Hentati-Sundberg, J. Breeding Failures and Reduced Nest Attendance in Response to Heat Stress in a High-Latitude Seabird. Mar. Ecol. Prog. Ser. 2023, HEATav3. [Google Scholar] [CrossRef]
- Patricia Silva, M.; Favero, M.; Casaux, R.; Baroni, A. The Status of Breeding Birds at Harmony Point, Nelson Island, Antarctica in Summer 1995/96. Mar. Ornithol. 1998, 26, 75–78. [Google Scholar]
- Krüger, L. An Update on the Southern Giant Petrels Macronectes Giganteus Breeding at Harmony Point, Nelson Island, Maritime Antarctic Peninsula. Polar Biol. 2019, 42, 1205–1208. [Google Scholar] [CrossRef]
- Oosthuizen, W.C.; Krüger, L.; Jouanneau, W.; Lowther, A.D. Unmanned Aerial Vehicle (UAV) Survey of the Antarctic Shag (Leucocarbo Bransfieldensis) Breeding Colony at Harmony Point, Nelson Island, South Shetland Islands. Polar Biol. 2020, 43, 187–191. [Google Scholar] [CrossRef]
- Harrison, P. Seabirds: An Identification Guide; Croom Helm, Beckenham: London, UK, 1983. [Google Scholar]
- Peter, H.-U.; Kaiser, M.; Gebauer, A. Ecological and Morphological Investigations on South Polar Skuas (Catharacta Maccormicki) and Brown Skuas (Catharacta Skua Lonnbergi) on Fildes Peninsula, King George Island, South Shetland Islands. Zool. Jb. Syst. Jena 1990, 117, 201–218. [Google Scholar]
- Geldart, E.A.; Barnas, A.F.; Semeniuk, C.A.D.; Gilchrist, H.G.; Harris, C.M.; Love, O.P. A Colonial-Nesting Seabird Shows No Heart-Rate Response to Drone-Based Population Surveys. Sci. Rep. 2022, 12, 18804. [Google Scholar] [CrossRef]
- Duda, T.; Canty, M. Unsupervised Classification of Satellite Imagery: Choosing a Good Algorithm. Int. J. Remote Sens. 2002, 23, 2193–2212. [Google Scholar] [CrossRef]
- Belgiu, M.; Drǎguţ, L. Comparing Supervised and Unsupervised Multiresolution Segmentation Approaches for Extracting Buildings from Very High Resolution Imagery. ISPRS J. Photogramm. Remote Sens. 2014, 96, 67–75. [Google Scholar] [CrossRef]
- Armstrong, E.A. The behaviour of birds in continuous daylight. Ibis (Lond. 1859). 1954, 96, 1–30. [Google Scholar] [CrossRef]
- Burton, R.W. Breeding Biology of the Brown Skua, Catharacta Skua Lonnbergi (Mathews), at Signy Island, South Orkney Islands. Br. Antarct. Surv. Bull. 1968, 15, 9–28. [Google Scholar]
- Hahn, S.; Peter, H.-U.; Bauer, S. Skuas at Penguin Carcass: Patch Use and State-Dependent Leaving Decisions in a Top-Predator. Proc. R. Soc. B Biol. Sci. 2005, 272, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models; Cambridge University Press: Cambridge, UK, 2017; ISBN 9781139028271. [Google Scholar]
- ARAUJO, M.; NEW, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Soberón, J. Species Distribution Modeling and Ecological Niche Modeling: Getting the Concepts Right. Nat. Conserv. 2012, 10, 102–107. [Google Scholar] [CrossRef]
- Quillfeldt, P.; Engler, J.O.; Silk, J.R.D.; Phillips, R.A. Influence of Device Accuracy and Choice of Algorithm for Species Distribution Modelling of Seabirds: A Case Study Using Black-Browed Albatrosses. J. Avian Biol. 2017, 48, 1549–1555. [Google Scholar] [CrossRef]
- VanDerWal, J.; Shoo, L.P.; Graham, C.; Williams, S.E. Selecting Pseudo-Absence Data for Presence-Only Distribution Modeling: How Far Should You Stray from What You Know? Ecol. Modell. 2009, 220, 589–594. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting Pseudo-Absences for Species Distribution Models: How, Where and How Many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Ridgeway, G. Generalized Boosted Models: A Guide to the Gbm Package; 20 December 2020; Available online: https://cran.r-project.org/web/packages/gbm/vignettes/gbm.pdf (accessed on 20 December 2022).
- Greenwell, B.; Boehmke, B.; Cunningham, J. Gbm: Generalized Boosted Regression Models, R Package Version 2.1.5; 2019. Available online: https://CRAN.R-project.org/package=gbm (accessed on 20 December 2022).
- Boehmke, B.; Greenwell, B. Hands-On Machine Learning with R; Chapman and Hall/CRC: London, UK, 2019; ISBN 9780367816377. [Google Scholar]
- Pereira, N.R.; Loiola, R.; Rodrigues, S.; de Oliveira, C.; Büttenbender, S.; Guterres, S.; Pohlmann, A.; Farsky, S. Mechanisms of the Effectiveness of Poly(ε-Caprolactone) Lipid-Core Nanocapsules Loaded with Methotrexate on Glioblastoma Multiforme Treatment. Int. J. Nanomed. 2018, 13, 4563–4573. [Google Scholar] [CrossRef]
- Farzin Shabani Assessing Accuracy Methods of Species Distribution Models: AUC, Specificity, Sensitivity and the True Skill Statistic. Glob. J. Hum.-Soc. Sci. 2018, 18, 7–18.
- Pietz, P. Feeding and Nesting Ecology of Sympatric South Polar and Brown Skuas. Auk Ornithol. Adv. 1987, 194, 617–627. [Google Scholar] [CrossRef]
- Quintana, R.D.; Travaini, A. Characteristics of nest sites of skuas and kelp gull in the Antarctic peninsula. J. F. Ornithol. 2000, 71, 236–249. [Google Scholar] [CrossRef]
- Pizarro, M.; Contreras, R.A.; Köhler, H.; Zúñiga, G.E. Desiccation Tolerance in the Antarctic Moss Sanionia Uncinata. Biol. Res. 2019, 52, 46. [Google Scholar] [CrossRef] [PubMed]
- Oswald, S.A.; Bearhop, S.; Furness, R.W.; Huntley, B.; Hamer, K.C. Heat Stress in a High-Latitude Seabird: Effects of Temperature and Food Supply on Bathing and Nest Attendance of Great Skuas Catharacta Skua. J. Avian Biol. 2008, 39, 163–169. [Google Scholar] [CrossRef]
- Michielsen, R.J.; Ausems, A.N.M.A.; Jakubas, D.; Pętlicki, M.; Plenzler, J.; Shamoun-Baranes, J.; Wojczulanis-Jakubas, K. Nest Characteristics Determine Nest Microclimate and Affect Breeding Output in an Antarctic Seabird, the Wilson’s Storm-Petrel. PLoS ONE 2019, 14, e0217708. [Google Scholar] [CrossRef] [PubMed]
- Ritz, M.S.; Hahn, S.; Janicke, T.; Peter, H.-U. Hybridisation between South Polar Skua (Catharacta Maccormicki) and Brown Skua (C. Antarctica Lonnbergi) in the Antarctic Peninsula Region. Polar Biol. 2006, 29, 153–159. [Google Scholar] [CrossRef]
- Petry, M.V.; Valls, F.C.L.; de Souza Petersen, E.; Krüger, L.; da Cruz Piuco, R.; dos Santos, C.R. Breeding Sites and Population of Seabirds on Admiralty Bay, King George Island, Antarctica. Polar Biol. 2016, 39, 1343–1349. [Google Scholar] [CrossRef]
- Petry, M.V.; Valls, F.C.L.; Petersen, E.S.; Finger, J.V.G.; Krüger, L. Population Trends of Seabirds at Stinker Point, Elephant Island, Maritime Antarctica. Antarct. Sci. 2018, 30, 220–226. [Google Scholar] [CrossRef]
- Graña Grilli, M.; Montalti, D. Trophic Interactions between Brown and South Polar Skuas at Deception Island, Antarctica. Polar Biol. 2012, 35, 299–304. [Google Scholar] [CrossRef]
- Southwell, C.; Wotherspoon, S.; Emmerson, L. Emerging Evidence of Resource Limitation in an Antarctic Seabird Metapopulation after 6 Decades of Sustained Population Growth. Oecologia 2021, 196, 693–705. [Google Scholar] [CrossRef]
- Graña Grilli, M.; Libertelli, M.; Montalti, D. Diet of South Polar Skua Chicks in Two Areas of Sympatry with Brown Skua. Waterbirds 2011, 34, 495–498. [Google Scholar] [CrossRef]
- Phillips, R.A.; Phalan, B.; Forster, I.P. Diet and Long-Term Changes in Population Size and Productivity of Brown Skuas Catharacta Antarctica Lonnbergi at Bird Island, South Georgia. Polar Biol. 2004, 27, 555–561. [Google Scholar] [CrossRef]
- Krüger, L. Decreasing Trends of Chinstrap Penguin Breeding Colonies in a Region of Major and Ongoing Rapid Environmental Changes Suggest Population Level Vulnerability. Diversity 2023, 15, 327. [Google Scholar] [CrossRef]
- Jin, S.; Cao, S.; Li, R.; Gao, H.; Na, G. Trophic Transfer of Polycyclic Aromatic Hydrocarbons through the Food Web of the Fildes Peninsula, Antarctica. Environ. Sci. Pollut. Res. 2023, 30, 55057–55066. [Google Scholar] [CrossRef] [PubMed]
- Finger, J.V.G.; Corá, D.H.; Convey, P.; Cruz, F.S.; Petry, M.V.; Krüger, L. Anthropogenic Debris in an Antarctic Specially Protected Area in the Maritime Antarctic. Mar. Pollut. Bull. 2021, 172, 112921. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, J.; Bresesti, F.; Lozoya, J.P.; De Feo, B.; Krojmal, E.; Lacerot, G.; Braun, C.; Teixeira de Mello, F. Diet and Debris Ingestion of Skuas on Fildes Peninsula, King George Island, Antarctica. Mar. Pollut. Bull. 2022, 185, 114211. [Google Scholar] [CrossRef]
- de Seixas, M.M.M.; de Araújo, J.; Krauss, S.; Fabrizio, T.; Walker, D.; Ometto, T.; Matsumiya Thomazelli, L.; Vanstreels, R.E.T.; Hurtado, R.F.; Krüger, L.; et al. H6N8 Avian Influenza Virus in Antarctic Seabirds Demonstrates Connectivity between South America and Antarctica. Transbound. Emerg. Dis. 2022, 69, e3436–e3446. [Google Scholar] [CrossRef]
- Dias, M.P.; Martin, R.; Pearmain, E.J.; Burfield, I.J.; Small, C.; Phillips, R.A.; Yates, O.; Lascelles, B.; Borboroglu, P.G.; Croxall, J.P. Threats to Seabirds: A Global Assessment. Biol. Conserv. 2019, 237, 525–537. [Google Scholar] [CrossRef]
- Salmerón, N.; Belle, S.; Cruz, F.S.; Alegria, N.; Finger, J.V.G.; Corá, D.H.; Petry, M.V.; Hernández, C.; Cárdenas, C.A.; Krüger, L. Contrasting Environmental Conditions Precluded Lower Availability of Antarctic Krill Affecting Breeding Chinstrap Penguins in the Antarctic Peninsula. Sci. Rep. 2023, 13, 5265. [Google Scholar] [CrossRef]
- Hughes, K.A.; Convey, P.; Turner, J. Developing Resilience to Climate Change Impacts in Antarctica: An Evaluation of Antarctic Treaty System Protected Area Policy. Environ. Sci. Policy 2021, 124, 12–22. [Google Scholar] [CrossRef]
- Brooks, C.M.; Chown, S.L.; Douglass, L.L.; Raymond, B.P.; Shaw, J.D.; Sylvester, Z.T.; Torrens, C.L. Progress towards a Representative Network of Southern Ocean Protected Areas. PLoS ONE 2020, 15, e0231361. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santa Cruz, F.; Krüger, L. Breeding Population and Nesting Habitat of Skuas in the Harmony Point Antarctic Specially Protected Area. Diversity 2023, 15, 638. https://doi.org/10.3390/d15050638
Santa Cruz F, Krüger L. Breeding Population and Nesting Habitat of Skuas in the Harmony Point Antarctic Specially Protected Area. Diversity. 2023; 15(5):638. https://doi.org/10.3390/d15050638
Chicago/Turabian StyleSanta Cruz, Francisco, and Lucas Krüger. 2023. "Breeding Population and Nesting Habitat of Skuas in the Harmony Point Antarctic Specially Protected Area" Diversity 15, no. 5: 638. https://doi.org/10.3390/d15050638
APA StyleSanta Cruz, F., & Krüger, L. (2023). Breeding Population and Nesting Habitat of Skuas in the Harmony Point Antarctic Specially Protected Area. Diversity, 15(5), 638. https://doi.org/10.3390/d15050638