Abstract
Plant invasions are closely related to environmental filtering and biointeractions; however, the variations in invasive plant niches along latitudinal gradients in heterogeneous habitats remain unclear. In this study, we conducted a two-year survey in China spanning 21° N–37° N to explore the niche characteristics of plant species within communities invaded by the amphibious alien weed Alternanthera philoxeroides in both terrestrial and aquatic habitats as well as their latitudinal trends. We found that A. philoxeroides had the greatest niche breadth in the studied communities. The species pairs with the highest niche similarity were A. philoxeroides–Digitaria sanguinalis in terrestrial communities and Cyperus rotundus–Kyllinga brevifolia in aquatic communities. The niche similarity between A. philoxeroides and its accompanying species in terrestrial habitats was significantly higher than that in aquatic habitats (t = 5.954; p < 0.001). The niche breadth of A. philoxeroides had no obvious latitudinal trend, while the niche breadth of its accompanying species in the terrestrial community significantly decreased with increasing latitude (F7, 57 = 4.364, p = 0.001). In the terrestrial communities, the niche similarity between A. philoxeroides and its accompanying species significantly decreased with increasing latitude (F7, 57 = 3.671, p = 0.003), while the niche overlap significantly increased with increasing latitude (F7, 57 = 8.916, p < 0.001). However, the aquatic species’ niche characteristics had no obvious latitudinal trends. These findings indicated that habitat heterogeneity significantly affected the species’ niche characteristics in A. philoxeroides-invaded communities. Environmental filtering at low latitudes allowed the invasive and accompanying species to evolve similar niches, while the cold climate at high latitudes increased the niche overlap between the invader and accompanying species. Our findings are crucial for predicting the dynamics of invasive plant communities under global change and for understanding the mechanisms of species coexistence.
1. Introduction
In the context of global climate change and economic integration, biological invasions are one of the five serious global environmental problems of the 21st century [1,2]. Frequent resource fluctuations and disturbances resulting from human activities have led to increasing plant invasions, causing a dramatic decline in biodiversity and great damage to the structures and functions of ecosystems [3]. Functional differences between native and alien species are crucial factors for accelerating successful plant invasions; invasive plants usually have a higher growth rate and higher photosynthesis, water use efficiency, specific leaf area, nitrogen content and interspecific competitiveness than their native accompanying species [4,5,6,7], as well as stronger responses to environmental changes [8,9,10]. A recent meta-analysis has shown that multiple invasive plants could also have a synergistic invasion effect, resulting in twice the negative effect on neighboring natives than on other alien plants [11]. However, biological invasions and native species diversity often demonstrate positive relationships in large-scale observational studies but are negatively correlated in small-scale experimental studies; this phenomenon is referred to as the invasion paradox [12,13,14]. Exploring the performance and ecological effect of invasive plants at large spatial scales is helpful for implementing effective invasion prevention and biodiversity protection measures.
A niche represents the utilization degree of various resources by plant species, while niche breadth, niche similarity and niche overlap are important indicators for measuring plant niches; in addition, these factors reflect the role of plant species in communities and their demands for living environments [15,16]. Under conditions with limited resources, species that have similar niches will compete fiercely with each other and cannot stably coexist [17,18]. The expansion of invasive plant populations is usually associated with niche changes, and their niche characteristics are determined by environmental filtering, biointeractions, adaptive evolution and other factors [15,16,19,20]. Many invasive plants have undergone niche shifts and have expanded their climate niches through adaptive evolution and phenotypic plasticity, which is of great benefit for quickly occupying the open niche, and thus facilitates their invasion process [21,22,23]. In brief, the key to the successful invasion of alien plants is their ability to create ‘niche opportunities’ through various mechanisms, such as successfully coexisting in native communities through heterogeneous resource use or by increasing their diffusion ability by competing for the same resources with native species [24,25].
Hydrothermal fluctuations caused by latitudinal variations significantly affect the niche changes in invasive plant species. Compared with native plants, invasive plants usually have higher climate tolerance and phenological plasticity, which benefit their ability to form a wide niche breadth to resist climate change [26,27,28,29]. Moreover, global warming weakens the biotic resistance of native plant communities at higher latitudes, thus creating favorable niche spaces for invasive plants in those regions [30]. The latitude–niche breadth hypothesis predicts that tropical species evolve into narrower niches due to weak seasonality and environmental variability at low latitudes, while frequent climate changes at high latitudes promote the evolution of wider niches [31]. Rapoport’s ecogeographic rules also propose that the distribution ranges of species shrink with decreasing latitude, and intense interspecific competition at low latitudes reduces species niche breadth, which negatively correlates with latitude [32,33]. The germination niche breadth and climate niche breadth of some invasive plants have been found to increase with increasing latitude; however, their water quality niche breadth decreases with increasing latitude, which is due to their long-term adaptations to water eutrophication [27,33]. However, the niche relationships between invasive and native plants along the larger latitudinal gradient are still not well understood.
The alligator weed Alternanthera philoxeroides, a type of noxious invasive weed with clonal growth, is native to South America and has extensively invaded the United States, Australia, South Africa, China and other parts of the world [34]. A. philoxeroides is an amphibious invader, and in China it has invaded aquatic and terrestrial habitats in more than 20 provinces, causing serious damage to ecological environments and economic development [35]. Previous studies have found that A. philoxeroides is more responsive to environmental fluctuations than its native congener A. sessilis, and climate warming will promote the climate niche expansion of the terrestrial ecotype of A. philoxeroides, intensifying its competitive substitution for native plants at high latitudes [36,37]. In flooded environments, A. philoxeroides increases the diameter of its stolon to strengthen its interspecific competitiveness [38]. In addition, with increasing latitudes in China, aquatic A. philoxeroides communities have higher β-diversity, while terrestrial A. philoxeroides communities have higher similarity [34]. However, the niche characteristics of A. philoxeroides and its accompanying species at large spatial scales are unclear.
In this study, we investigated the plant communities invaded by A. philoxeroides within the latitudinal range from 21° N to 37° N in mainland China, and we hypothesize that the species niche characteristics of communities invaded by A. philoxeroides along a latitudinal gradient have different trends in aquatic and terrestrial habitats. We specifically address the following questions: (1) Do the niche breadth, niche similarity and niche overlap of the A. philoxeroides communities in heterogeneous habitats differ? (2) Are the variations in each of the above three indicators different along latitudinal gradients in heterogeneous habitats?
2. Materials and Methods
2.1. Site Selection
From July to August of 2019–2021, the time of year during which the growth period of plant species is vigorous, we selected sites that were invaded by A. philoxeroides over an area of more than 100 m2 to set the sampling plots, and the plots contained low (A. philoxeroides coverage < 40%), moderate (A. philoxeroides coverage was 40%–60%) and heavy (A. philoxeroides coverage > 60%) invaded levels. Starting from 21° N, a total of 8 latitudinal clusters were set, each 2° apart (the low latitudinal gradient containing Cluster 1: 21° N–23° N and Cluster 2: 23° N–25° N; the middle latitudinal gradient containing Cluster 3: 25° N–27° N, Cluster 4: 27° N–29° N, Cluster 5: 29° N–31° N, and Cluster 6: 31° N–33° N; and the high latitudinal gradient containing Cluster 7: 33° N–35° N and Cluster 8: 35° N–37° N) [34,36,39]. Five terrestrial plots were set along each latitudinal cluster, and five aquatic plots were also set in areas with similar habitat conditions along the same latitudinal cluster [35]. The area of each plot was 10 m × 10 m, and plots with the same ecotype were set at intervals of more than 10 km [39]. In total, we set 80 sampling plots (40 terrestrial and 40 aquatic; Figure 1), with the types of terrestrial sites mainly comprising roadsides (13 plots), gardens (8 plots), abandoned land (7 plots), farmland (7 plots) and riparian zones (5 plots), while the types of aquatic sites comprised ponds (12 plots), rivers (10 plots), puddles (9 plots), water channels (6 plots) and paddy fields (3 plots) (Supplementary Table S1). For the sampling sites in which there may be a fluid transition between terrestrial and aquatic habitats or in which there may be intermittent changes in habitat type due to climate fluctuations, we combined the morphological plasticity of A. philoxeroides to identify the type of plot in question. The whole study area covered 20 cities within 10 provinces in China and spanned 3 climatic regions, including tropical, subtropical and temperate regions.
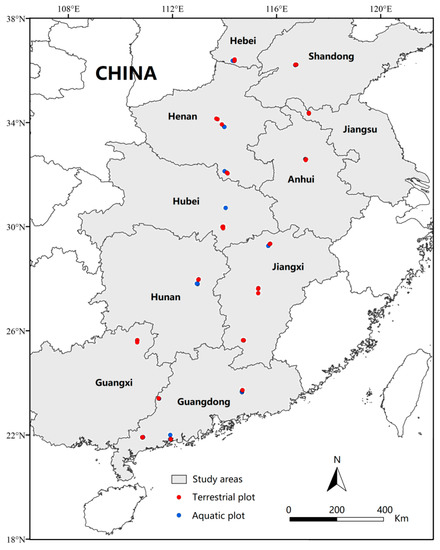
Figure 1.
Sampling plots of A. philoxeroides-invaded communities in China along latitudinal gradients (21° N to 37° N). Plot size: 10 × 10 m; transect size: 10 m; quadrat size: 0.5 × 0.5 m.
2.2. Field Survey
For terrestrial plots and some aquatic plots with shallow water levels, we evenly set three 10 m transects in each plot and then evenly set five quadrats with an area of 0.5 m × 0.5 m along each transect for the plant investigation [35,37,40]. For a few aquatic plots with deep water levels, to facilitate the investigation, we chose a blanket patch of A. philoxeroides that was close to the bank and set three continuous 10 m transects along the horizontal direction of the aquatic A. philoxeroides patches for investigation. We used the survey method of Wu et al. (2016) to record the names, height and coverage of the plant species in every quadrat [39]. We also recorded the individuals (for nonclonal plants), tillers (for graminaceous plants) or numbers of asexual branches (for clonal plants) as the abundance of plant species in each quadrat [35,39]. The longitude, latitude and altitude of every plot were recorded using a handheld GPS receiver (Garmin Inc., Olathe, KS, USA). Photos or collected samples of unknown plants were taken during the field survey, and we then used the ‘Chinese Virtual Herbarium’ (https://www.cvh.ac.cn, accessed on 10 March 2023) and the online database ‘Flora of China’ (http://www.iplant.cn/frps, accessed on 10 March 2023) to conduct species identification.
2.3. Data Calculation
2.3.1. Importance Value
We used the importance value (IV) as a comprehensive indicator to measure the dominance of plants in the A. philoxeroides-invaded communities. The IV was calculated as follows [39]:
- IV = (relative abundance + relative height + relative coverage)/3.
- The total IV was the sum of a plant species’ IV in 40 terrestrial or 40 aquatic plots.
2.3.2. Niche Characteristics
Each plot in this study was regarded as a comprehensive resource level. Based on the occurrence frequency of plant species in the plots (Table 1), we used the abundance and IV of 25 main plant species (frequency > 9) in the terrestrial plots vs. 24 main plant species (frequency > 4) in the aquatic plots as comprehensive indicators to measure their niche characteristics. The results calculated using this method better reflected the efficiency of the plant species’ utilization of environmental resources and could prevent errors caused by individual size differences in each species.
Niche breadth represents the sum of the multiple environmental resources that are used by organisms. Plants with a higher niche breadth can better utilize resources and tend to be generalized species, while plants with a lower niche breadth tend to be specialized species. We calculated niche breadth values using Levins’ method as follows [41]:
where B(L) is the species’ niche breadth, Pij is the relative IV of the ith species in the jth plot and r is the total number of sampling plots.
Niche similarity represents the similarity degree of the species’ resource utilization, and a lower niche similarity indicates that there are differences in resource utilization between plant species and that they might coexist in the community for a long time. We calculated the niche similarity value using the Schoener index as follows [42]:
where Cik is the niche similarity value between species i and k, and Pkj is the relative IV of the kth species in the jth plot.
Niche overlap represents the degree of species with similar niches sharing or competing for common resources when living in the same space; the greater the niche overlap value, the greater the similarity in resource utilization and the more intense the interspecific competition. Niche overlap was calculated as follows [43]:
where Lih is the niche overlap value of species i overlapping species h, Lhi is the niche overlap value of species h overlapping species i, Phj is the relative IV of the hth species in the jth plot, B(L)i is the niche breadth of species i and B(L)h is the niche breadth of species h.

Table 1.
Niche breadth (B(L)) of plant species with a high frequency in terrestrial and aquatic A. philoxeroides-invaded communities.
Table 1.
Niche breadth (B(L)) of plant species with a high frequency in terrestrial and aquatic A. philoxeroides-invaded communities.
| Code | Plant Species | Frequency | IV | B(L) | Code | Plant Species | Frequency | IV | B(L) |
|---|---|---|---|---|---|---|---|---|---|
| Terrestrial Community | Aquatic Community | ||||||||
| 1 | Alternanthera philoxeroides | 40 | 17.360 | 0.960 | 1 | Alternanthera philoxeroides | 40 | 23.752 | 0.943 |
| 2 | Digitaria sanguinalis | 30 | 2.936 | 0.414 | 2 | Paspalum paspaloides | 25 | 5.427 | 0.424 |
| 3 | Echinochloa crusgalli | 28 | 1.112 | 0.314 | 3 | Polygonum hydropiper | 20 | 0.727 | 0.301 |
| 4 | Bidens pilosa | 25 | 0.739 | 0.354 | 4 | Echinochloa crusgalli | 18 | 1.226 | 0.210 |
| 5 | Ipomoea nil | 23 | 0.599 | 0.294 | 5 | Bidens pilosa | 14 | 0.341 | 0.232 |
| 6 | Eleusine indica | 22 | 1.064 | 0.278 | 6 | Commelina communis | 9 | 0.534 | 0.107 |
| 7 | Cyperus votundus | 21 | 0.901 | 0.204 | 7 | Cyperus rotundus | 8 | 0.198 | 0.126 |
| 8 | Setaria viridis | 19 | 0.972 | 0.322 | 8 | Kyllinga brevifolia | 7 | 0.123 | 0.146 |
| 9 | Commelina communis | 18 | 1.301 | 0.285 | 9 | Eclipta prostrata | 7 | 0.217 | 0.086 |
| 10 | Cyperus rotundus | 16 | 0.303 | 0.269 | 10 | Humulus scandens | 7 | 0.444 | 0.099 |
| 11 | Acalypha australis | 16 | 0.198 | 0.287 | 11 | Ipomoea nil | 7 | 0.232 | 0.079 |
| 12 | Humulus scandens | 15 | 1.120 | 0.203 | 12 | Lemna minor | 6 | 0.284 | 0.115 |
| 13 | Ageratum conopids | 15 | 0.680 | 0.160 | 13 | Digitaria sanguinalis | 6 | 0.123 | 0.120 |
| 14 | Erigeron canadensis | 15 | 0.425 | 0.138 | 14 | Acorus calamus | 5 | 0.146 | 0.106 |
| 15 | Artemisia argyi | 15 | 0.342 | 0.278 | 15 | Glycine soja | 5 | 0.309 | 0.075 |
| 16 | Eclipta prostrata | 15 | 0.195 | 0.214 | 16 | Alternanthera sessilis | 5 | 0.184 | 0.074 |
| 17 | Cynodon dactylon | 14 | 0.695 | 0.108 | 17 | Eleusine indica | 5 | 0.190 | 0.096 |
| 18 | Polygonum hydropiper | 14 | 0.677 | 0.129 | 18 | Trigastrotheca stricta | 5 | 0.291 | 0.052 |
| 19 | Amaranthus blitum | 13 | 0.170 | 0.132 | 19 | Cyperus votundus | 5 | 0.169 | 0.111 |
| 20 | Erigeron annuus | 12 | 0.231 | 0.123 | 20 | Polygonum perfoliatum | 4 | 0.061 | 0.092 |
| 21 | Oxalis corniculata | 10 | 0.250 | 0.099 | 21 | Phragmites australis | 4 | 0.187 | 0.083 |
| 22 | Paspalum paspaloides | 9 | 0.435 | 0.155 | 22 | Senna tora | 4 | 0.043 | 0.060 |
| 23 | Broussonetia papyrifera | 9 | 0.296 | 0.112 | 23 | Symphyotrichum subulatum | 4 | 0.197 | 0.053 |
| 24 | Ludwigia prostrata | 9 | 0.168 | 0.113 | 24 | Microstegium nodosum | 4 | 0.146 | 0.041 |
| 25 | Portulaca oleracea | 9 | 0.106 | 0.122 | |||||
2.4. Statistical Analyses
We conducted an independent sample t test (subset for α = 0.05) to compare the differences in the plant species’ niche characteristics between terrestrial and aquatic communities using SPSS16.0 software (SPSS Inc., Chicago, IL, USA) and conducted a one-way ANOVA and multiple comparisons of least-significant difference (LSD) to examine the variations in niche characteristics along the latitudinal gradient.
3. Results
3.1. Niche Characteristics at the Level of the Whole Community
3.1.1. Niche Breadth
The species niche breadth in the terrestrial communities was 0.099–0.960, and that of the aquatic communities was 0.041–0943. Of the habitat types, the heterogeneous habitats had the largest A. philoxeroides niche breadth (terrestrial B(L) = 0.960, aquatic B(L) = 0.943) (Table 1). In the terrestrial communities, the species with higher niche breadth values were Digitaria sanguinalis, Bidens pilosa, Setaria viridis and Echinochloa crusgalli, and Oxalis corniculata had the lowest niche breadth (Table 1). In the aquatic communities, Paspalum paspaloides, Polygonum hydropiper, B. pilosa and E. crusgalli also had higher niche breadth values, and Microstegium nodosum had the lowest niche breadth (Table 1). The t test showed that there was no significant difference in the niche breadth values of the common species between the terrestrial and the aquatic communities (t = 1.598, p = 0.117).
3.1.2. Niche Similarity
A total of 25 plant species in the terrestrial community formed 300 species pairs, and the values of their niche similarity were 0–0.547. Among these values, the niche similarity of two species pairs was more than 0.5 (containing A. philoxeroides–D. sanguinalis, S. viridis–Humulus scandens), accounting for 0.67% of the total species pairs. The niche similarity values of 11 species pairs were 0.4–0.5, accounting for 3.67% of the total species pairs. The niche similarity values of 46 species pairs were 0.3–0.4, accounting for 15.33% of the total number of species pairs. The niche similarity of 241 species pairs was less than 0.3, accounting for 80.33% of the total number of species pairs. The niche similarity between A. philoxeroides and its accompanying species was generally higher than that between each accompanying species (Figure 2a).
There were 276 species pairs formed from 24 plant species in the aquatic communities, with a niche similarity of 0–0.511. The niche similarity of one species pair was more than 0.5 (C. rotundus–Kyllinga brevifolia), accounting for 0.36% of the total species pairs. The niche similarity of seven species pairs was 0.4–0.5, accounting for 2.54% of the total species pairs. The niche similarity of 20 species pairs was 0.3–0.4, accounting for 7.25% of the total species pairs. The niche similarity of 248 species pairs was less than 0.3, accounting for 89.85% of the total species pairs (Figure 2b). The t test showed that the niche similarity between A. philoxeroides and its accompanying species in the terrestrial communities was significantly higher than that between A. philoxeroides and its accompanying species in the aquatic communities (t = 5.954, p < 0.001).

Figure 2.
Niche similarity of plant species with a high frequency in terrestrial (a) and aquatic (b) A. philoxeroides-invaded communities. Arabic numbers are codes of plant species (as shown in Table 1 for terrestrial and aquatic communities, respectively).
Figure 2.
Niche similarity of plant species with a high frequency in terrestrial (a) and aquatic (b) A. philoxeroides-invaded communities. Arabic numbers are codes of plant species (as shown in Table 1 for terrestrial and aquatic communities, respectively).

3.1.3. Niche Overlap
The species niche overlap values of the terrestrial communities were 0–0.026. The niche overlap between A. philoxeroides and its accompanying species was 0.022–0.026, which was generally higher than that between each accompanying species. The niche overlap of A. philoxeroides-P. paspaloides was the highest. The niche overlap of 49 species pairs was more than 0.01, accounting for 16.33% of the total species pairs. The niche overlap values of 64 species pairs were 0.005–0.01, accounting for 21.33% of the total species pairs. The niche overlap of 187 species pairs was less than 0.005, accounting for 62.33% of the total species pairs.
In the aquatic communities, the species niche overlap values were 0–0.029. The niche overlap values between A. philoxeroides and its accompanying species were 0.017–0.029, which were generally higher than those between each accompanying species. The niche overlap values of A. philoxeroides–Symphyotrichum subulatum and A. philoxeroides–M. nodosum were both the highest. The niche overlap values of 58 species pairs were more than 0.01, accounting for 21.01% of the total species pairs. The niche overlap values of 56 species pairs were 0.005–0.01, accounting for 20.29% of the total species pairs. The niche overlap values of 162 species pairs were less than 0.005, accounting for 58.70% of the total species pairs. The t test showed that there was no significant difference in the niche overlap of A. philoxeroides–accompanying species between terrestrial and aquatic communities (t = 0.488, p = 0.630).
3.2. Latitudinal Trends of Niche Characteristics
3.2.1. Variations in Niche Breadth along the Latitudinal Gradient
The niche breadth of A. philoxeroides had no obvious fluctuations along the latitude gradient. For the accompanying species in the terrestrial communities, compared to the other species, Commelina communis, B. pilosa and Eleusine indica had greater niche breadths at low latitudes; D. sanguinalis, E. indica, E. crusgalli and S. viridis had greater niche breadths at middle latitudes; and S. viridis and H. scandens had greater niche breadths at high latitudes. Conspicuously, D. sanguinalis always had greater niche breadths along the whole latitudinal gradient (Figure 3a). The ANOVA showed that the average niche breadth of the accompanying species significantly decreased with increasing latitude in the terrestrial habitats (F7, 57 = 4.364, p = 0.001); in particular, the niche breadth of Cluster 2 at low latitudes was significantly higher than that at middle and high latitudes.
In the aquatic communities, P. hydropiper and E. crusgalli had greater niche breadths at low latitudes, P. paspaloides had an extremely large niche breadth at middle latitudes, and P. paspaloides, E. crusgalli and P. hydropiper had greater niche breadths at high latitudes (Figure 3b). The ANOVA showed that the average niche breadth of the accompanying species in the aquatic community had no significant latitudinal trend (F7, 45 = 1.170, p = 0.343).

Figure 3.
Variations in niche breadth of the accompanying species along a latitudinal gradient in terrestrial (a) and aquatic (b) A. philoxeroides-invaded communities. Definition of abbreviations: Aa—Alopecurus aequalis; Ac—Ageratum conyzoides; As—Asarum sagittarioides; Ase—Alternanthera sessilis; Bp—Bidens pilosa; Bpa—Broussonetia papyrifera; Bpl—Bolboschoenus planiculmis; Bs—Beckmannia syzigachne; Cc—Commelina communis; Cd—Cynodon dactylon; Cde—Ceratophyllum demersum; Cr—Cyperus rotundus; Cv—Cyperus votundus; Ds—Digitaria sanguinalis; Ea—Erigeron annuus; Ec—Echinochloa crusgalli; Eca—Echinochloa caudata; Ecan—Erigeron canadensis; Ecr—Eichhornia crassipes; Ei—Eleusine indica; Ep—Eclipta prostrata; Go—Galium odoratum; Gs—Glycine soja; Hs—Humulus scandens; Ia—Ipomoea aquatica; In—Ipomoea nil; Lc—Leptochloa chinensis; Lm—Lemna minor; Mn—Microstegium nodosum; Mv—Myriophyllum verticillatum; Oj—Oenanthe javanica; Pa—Phragmites australis; Pb—Panicum bisulcatum; Ph—Polygonum hydropiper; Pp—Paspalum paspaloides; Ps—Pistia stratiotes; Pu—Phyllanthus urinaria; Sc—Sesbania cannabina; Ss—Symphyotrichum subulatum; Sv—Setaria viridis; Ts—Trigastrotheca stricta; Vp—Veronica polita. A and C, A and BC indicate the significant differences at the p < 0.01 level, respectively; while the A and AB, AB and BC, BC and C indicate no significant difference, respectively.
Figure 3.
Variations in niche breadth of the accompanying species along a latitudinal gradient in terrestrial (a) and aquatic (b) A. philoxeroides-invaded communities. Definition of abbreviations: Aa—Alopecurus aequalis; Ac—Ageratum conyzoides; As—Asarum sagittarioides; Ase—Alternanthera sessilis; Bp—Bidens pilosa; Bpa—Broussonetia papyrifera; Bpl—Bolboschoenus planiculmis; Bs—Beckmannia syzigachne; Cc—Commelina communis; Cd—Cynodon dactylon; Cde—Ceratophyllum demersum; Cr—Cyperus rotundus; Cv—Cyperus votundus; Ds—Digitaria sanguinalis; Ea—Erigeron annuus; Ec—Echinochloa crusgalli; Eca—Echinochloa caudata; Ecan—Erigeron canadensis; Ecr—Eichhornia crassipes; Ei—Eleusine indica; Ep—Eclipta prostrata; Go—Galium odoratum; Gs—Glycine soja; Hs—Humulus scandens; Ia—Ipomoea aquatica; In—Ipomoea nil; Lc—Leptochloa chinensis; Lm—Lemna minor; Mn—Microstegium nodosum; Mv—Myriophyllum verticillatum; Oj—Oenanthe javanica; Pa—Phragmites australis; Pb—Panicum bisulcatum; Ph—Polygonum hydropiper; Pp—Paspalum paspaloides; Ps—Pistia stratiotes; Pu—Phyllanthus urinaria; Sc—Sesbania cannabina; Ss—Symphyotrichum subulatum; Sv—Setaria viridis; Ts—Trigastrotheca stricta; Vp—Veronica polita. A and C, A and BC indicate the significant differences at the p < 0.01 level, respectively; while the A and AB, AB and BC, BC and C indicate no significant difference, respectively.

3.2.2. Variations in Niche Similarity along the Latitudinal Gradient
The niche similarity between A. philoxeroides and its accompanying species was generally higher than that between each accompanying species, and with increasing latitude, the niche similarity showed an overall downward trend. In the terrestrial community, the species pairs of A. philoxeroides–C. communis, A. philoxeroides–D. sanguinalis, A. philoxeroides–B. pilosa, C. communis–P. paspaloides and B. pilosa–E. indica had higher niche similarity at low latitudes than at other latitudes; A. philoxeroides–E. indica, A. philoxeroides–D. sanguinalis, A. philoxeroides–S. viridis, E. crusgalli–S. viridis, E. indica–S. viridis and H. scandens–S. viridis had higher niche similarity at middle latitudes; and A. philoxeroides–H. scandens, A. philoxeroides–Ipomoea nil, H. scandens–E. crusgalli and D. sanguinalis–E. crusgalli had higher niche similarity at high latitudes (Figure 4a). The ANOVA showed that niche similarity between A. philoxeroides and its accompanying species significantly decreased with increasing latitude in the terrestrial habitats (F7, 57 = 3.671, p = 0.003), especially the niche similarity of Cluster 2 at low latitudes, which was significantly higher than that at middle and high latitudes (Figure 5a).
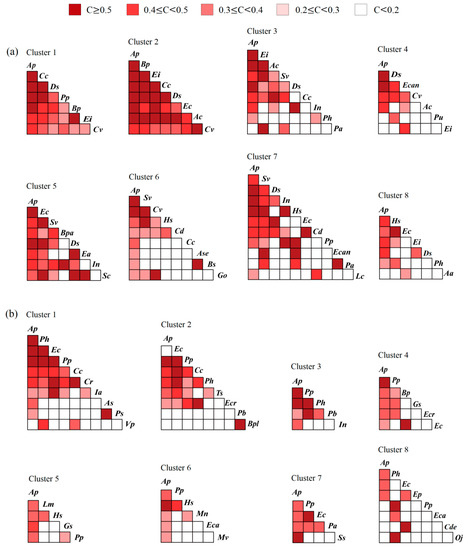
Figure 4.
Niche similarity between the dominant species of each latitudinal cluster in terrestrial (a) and aquatic (b) A. philoxeroides-invaded communities. Definition of abbreviations: Aa—Alopecurus aequalis; Ac—Ageratum conyzoides; Ap—Alternanthera philoxeroides; As—Asarum sagittarioides; Ase—Alternanthera sessilis; Bp—Bidens pilosa; Bpa—Broussonetia papyrifera; Bpl—Bolboschoenus planiculmis; Bs—Beckmannia syzigachne; Cc—Commelina communis; Cd—Cynodon dactylon; Cde—Ceratophyllum demersum; Cr—Cyperus rotundus; Cv—Cyperus votundus; Ds—Digitaria sanguinalis; Ea—Erigeron annuus; Ec—Echinochloa crusgalli; Eca—Echinochloa caudata; Ecan—Erigeron canadensis; Ecr—Eichhornia crassipes; Ei—Eleusine indica; Ep—Eclipta prostrata; Go—Galium odoratum; Gs—Glycine soja; Hs—Humulus scandens; Ia—Ipomoea aquatica; In—Ipomoea nil; Lc—Leptochloa chinensis; Lm—Lemna minor; Mn—Microstegium nodosum; Mv—Myriophyllum verticillatum; Oj—Oenanthe javanica; Pa—Phragmites australis; Pb—Panicum bisulcatum; Ph—Polygonum hydropiper; Pp—Paspalum paspaloides; Ps—Pistia stratiotes; Pu—Phyllanthus urinaria; Sc—Sesbania cannabina; Ss—Symphyotrichum subulatum; Sv—Setaria viridis; Ts—Trigastrotheca stricta; Vp—Veronica polita.
In aquatic communities, the species pairs of A. philoxeroides–Pistia stratiotes, A. philoxeroides–Eichhornia crassipes, A. philoxeroides–P. paspaloides, P. stratiotes–Asarum sagittarioides and Bolboschoenus planiculmis–Panicum bisulcatum had higher niche similarity at low latitudes than at the other latitudes; A. philoxeroides–P. paspaloides, A. philoxeroides–I. nil, A. philoxeroides–E. caudata, A. philoxeroides–Myriophyllum verticillatum and P. paspaloides–P. bisulcatum had higher niche similarity at middle latitudes; and A. philoxeroides–Phragmites australis, A. philoxeroides–S. subulatum, P. paspaloides–Oenanthe javanica and Ceratophyllum demersum–E. crusgalli had higher niche similarity at high latitudes (Figure 4b). The ANOVA showed that the niche similarity between A. philoxeroides and its accompanying species of each cluster had no significant latitudinal trend in the aquatic habitat (F7, 45 = 0.981, p = 0.459) (Figure 5b).

Figure 5.
Variations in niche similarity between A. philoxeroides and the dominant accompanying species of each latitudinal cluster in terrestrial (a) and aquatic (b) habitats. A and C, A and BC indicate the significant differences at the p < 0.01 level, respectively; while the A and AB, AB and BC, BC and C indicate no significant difference, respectively.
Figure 5.
Variations in niche similarity between A. philoxeroides and the dominant accompanying species of each latitudinal cluster in terrestrial (a) and aquatic (b) habitats. A and C, A and BC indicate the significant differences at the p < 0.01 level, respectively; while the A and AB, AB and BC, BC and C indicate no significant difference, respectively.

3.2.3. Variations in Niche Overlap along the Latitudinal Gradient
The niche overlap values of A. philoxeroides and its accompanying species were generally higher than those of the species accompanying A. philoxeroides. With increasing latitude, the niche overlap values between plant species in the A. philoxeroides-invaded communities showed an overall downward trend (Figure 6). In the terrestrial communities, the species pairs of A. philoxeroides–D. sanguinalis, A. philoxeroides–B. pilosa, B. pilosa–E. indica and Ageratum conyzoides–C. votundus had higher niche overlap values at low latitudes than at other latitudes; A. philoxeroides–A. conyzoides, A. philoxeroides–C. communis, A. philoxeroides–C. votundus, E. indica–Erigeron canadensis and E. crusgalli–Sesbania cannabina had higher niche overlap values at middle latitudes; and A. philoxeroides–S. viridis, A. philoxeroides–D. sanguinalis, A. philoxeroides–I. nil, A. philoxeroides–H. scandens, A. philoxeroides–E. crusgalli and A. philoxeroides–Cynodon dactylon had higher niche overlap values at high latitudes (Figure 6a). The ANOVA showed that the niche overlap between A. philoxeroides and its accompanying species in Cluster 7 at high latitudes was significantly higher than that at low and middle latitudes (F7, 57 = 8.916, p < 0.001) (Figure 7a).
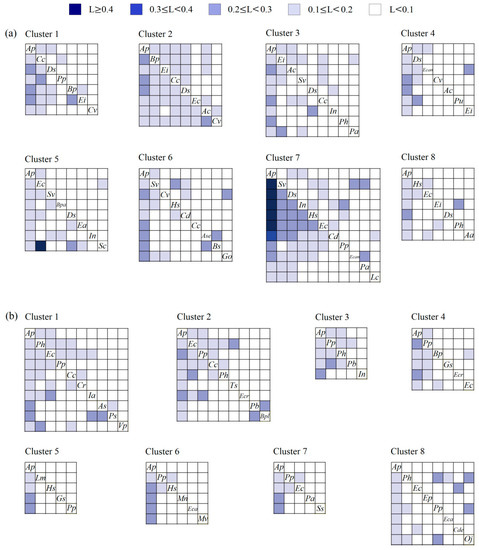
Figure 6.
Niche overlap between the dominant species of each latitudinal cluster in terrestrial (a) and aquatic (b) A. philoxeroides-invaded communities. Definition of abbreviations: Aa—Alopecurus aequalis; Ac—Ageratum conyzoides; Ap—Alternanthera philoxeroides; As—Asarum sagittarioides; Ase—Alternanthera sessilis; Bp—Bidens pilosa; Bpa—Broussonetia papyrifera; Bpl—Bolboschoenus planiculmis; Bs—Beckmannia syzigachne; Cc—Commelina communis; Cd—Cynodon dactylon; Cde—Ceratophyllum demersum; Cr—Cyperus rotundus; Cv—Cyperus votundus; Ds—Digitaria sanguinalis; Ea—Erigeron annuus; Ec—Echinochloa crusgalli; Eca—Echinochloa caudata; Ecan—Erigeron canadensis; Ecr—Eichhornia crassipes; Ei—Eleusine indica; Ep—Eclipta prostrata; Go—Galium odoratum; Gs—Glycine soja; Hs—Humulus scandens; Ia—Ipomoea aquatica; In—Ipomoea nil; Lc—Leptochloa chinensis; Lm—Lemna minor; Mn—Microstegium nodosum; Mv—Myriophyllum verticillatum; Oj—Oenanthe javanica; Pa—Phragmites australis; Pb—Panicum bisulcatum; Ph—Polygonum hydropiper; Pp—Paspalum paspaloides; Ps—Pistia stratiotes; Pu—Phyllanthus urinaria; Sc—Sesbania cannabina; Ss—Symphyotrichum subulatum; Sv—Setaria viridis; Ts—Trigastrotheca stricta; Vp—Veronica polita.
In the aquatic communities, A. philoxeroides–P. stratiotes, A. philoxeroides–E. crassipes, A. philoxeroides–P. paspaloides, P. stratiotes–A. sagittarioides and B. planiculmis had higher niche overlap values at low latitudes than at other latitudes; A. philoxeroides–P. paspaloides, A. philoxeroides–I. nil, A. philoxeroides–E. caudata, A. philoxeroides–M. verticillatum and P. paspaloides–P. bisulcatum had higher niche overlap at middle latitudes; and A. philoxeroides–P. australis, A. philoxeroides–S. subulatum, P. paspaloides–O. javanica, C. demersum–E. crusgalli, P. paspaloides–P. hydropiper and O. javanica–P. hydropiper had higher niche overlap at high latitudes (Figure 6b). The ANOVA showed that the niche overlap between A. philoxeroides and its accompanying species of each cluster had no significant latitudinal trend in the aquatic habitats (F7, 45 = 1.042, p = 0.419) (Figure 7b).

Figure 7.
Variations in niche overlap between A. philoxeroides and the dominant accompanying species of each latitudinal cluster in terrestrial (a) and aquatic (b) habitats. A and B indicate the significant differences at the p < 0.01 level.
Figure 7.
Variations in niche overlap between A. philoxeroides and the dominant accompanying species of each latitudinal cluster in terrestrial (a) and aquatic (b) habitats. A and B indicate the significant differences at the p < 0.01 level.

4. Discussion
4.1. Causes of Niche Characteristic Formation in Heterogeneous Habitats
Of the species examined in this study, A. philoxeroides had the greatest niche breadth in both aquatic and terrestrial habitats, reflecting that this invader had a strong ecological adaptation capacity and high resource utilization efficiency, which may be attributed to the cold-tolerant genotype and the growth–defense tradeoff of A. philoxeroides, which has benefited from adapting to harsh habitat conditions and thus occupies a broader niche space [44,45,46]. This conclusion also supports the niche breadth hypothesis, which states that there is a positive correlation between the size of a species’ geographical range and its niche breadth [17], and that plant populations that continuously match habitat conditions will have greater opportunities for distribution expansion [33,47]. However, D. sanguinalis, B. pilosa and S. viridis in the terrestrial habitats and P. paspaloides and P. hydropiper in the aquatic habitats also had greater niche breadths, which might be attributed to their bio-properties. D. sanguinalis and S. viridis had tillering growth and produced massive seeds with strong transmission ability through generative propagation. The tall individuals of B. pilosa and P. hydropiper would imply that they compete for more light resources, while P. paspaloides was found to be a perennial species and had both vegetative and generative propagations (Table S2); it could spread rapidly through the rhizomes and stolons. All of these superior functional traits promoted their stable existence at each stage of community succession. Although the niche breadth of most species was positively correlated with their IV in the A. philoxeroides community, some dominant accompanying species, such as S. viridis and C. communis, were incidentally present, indicating that IV was not the unique factor determining niche breadth, and niche breadth might also be determined by the interactive effect of species distribution frequency and habitat heterogeneity [48].
In our study, niche similarity in the terrestrial communities was generally higher than that in the aquatic communities, which might be due to the higher species diversity of the terrestrial communities [37]. Neutral theory states that species similarity rather than species difference contributes to high community diversity; thus, continuous biointeractions among multiple species in terrestrial A. philoxeroides communities promote coexistence groups with high niche similarity [49]. In addition, Wu et al. (2022) found that A. philoxeroides invasions caused higher community homogenization in terrestrial habitats than in aquatic habitats, and functional groups of Gramineae and Compositae that have strong invasion resistance widely exist in the community [34], which causes the plant species in terrestrial habitats to have closer geographical affinity and facilitates their convergent evolution. Moreover, environmental filtering caused by strong disturbances (e.g., human disturbances and insect feeding) in terrestrial habitats have led to the repeated colonization of plant species in the same species pool, which have caused similar species to show obvious aggregation and thus facilitated the coexistence of species with high niche similarity [19]. However, the structure of aquatic communities is relatively simple and is more sensitive to environmental changes; thus, aquatic plant species might require a smaller niche similarity to maintain survival [50].
Species niche overlap in terrestrial and aquatic habitats was low, indicating that plant species in the A. philoxeroides-invaded communities tended to reduce their niche overlap to coexist in large-scale spaces [18]. However, the niche overlap between A. philoxeroides and its accompanying species was higher than that between each accompanying species, indicating that A. philoxeroides, as an invader, had stronger competitiveness; it competed for resources with most plant species and even with narrow-niche species. Interestingly, the niche similarity and niche overlap of species pairs such as A. philoxeroides–D. sanguinalis and A. philoxeroides–P. paspaloides were all high, indicating that they had highly similar living habits and resource requirements and that more intense interspecific competition might thus occur with community succession. Previous studies have found that plant invasions cause a decline in niche overlap among native plants and thus enhance the competition between invasive and native plants [51]. Similarly, we also found that the accompanying species with large niche breadth did not form species pairs with higher niche overlap in the A. philoxeroides community, indicating that the resource utilization capacity of the accompanying species was weaker than that of A. philoxeroides at large spatial scales and that they might choose to reduce interspecific competition and/or improve mutual assistance for resisting invasion [29].
4.2. Causes of Niche Breadth Variations along the Latitudinal Gradient in Heterogeneous Habitats
Niche theory predicts that with changes in the environmental gradient, the difference in species specificity during the process of adaptive evolution will lead to changes in community structures [16]. Habitat heterogeneity can improve the phenotypic plasticity of plant species, while latitudinal gradients facilitate the continuous evolution of species ecological strategies [45]. In our study, the niche breadths of A. philoxeroides in both terrestrial and aquatic habitats were high, and no obvious latitudinal trend was observed, suggesting that the niche breadth of this species might be determined by both adaptive variation and phenotypic plasticity; thus, a larger niche breadth could benefit A. philoxeroides in resisting environmental fluctuations [26,45]. Previous studies have found that the phenotypic plasticity of A. philoxeroides increased with increasing latitude, reflecting its strong adaptations to changes in temperature, precipitation and herbivory along a latitudinal gradient, thus leading to the large niche breadth of A. philoxeroides in each latitudinal cluster [52,53,54]. This scenario may also have occurred because A. philoxeroides, as a pioneer species, has strong environmental tolerance; it can share similar habitat conditions at different latitudes and spread to higher latitudinal regions under climate change [36,55,56].
Temperature is the crucial factor affecting plant growth, and the distributions of plant species in terrestrial ecosystems are more vulnerable to thermal variations than to other variations [10]. Low-latitude regions have sufficient heat and exacerbate the competition between each accompanying species, while the cold climate of high latitudes might mitigate interspecific competition. However, invasive plants can continuously establish populations in new habitats at high latitudes and reduce the niche breadth of the accompanying plants by disturbing their competition and coexistence patterns [47]. In our study, species with larger niche breadths, such as H. scandens, which had both vegetative and generative propagations, and S. viridis, which had tall individuals (Table S2), also appeared in high-latitude terrestrial communities, which may be due to the continuous evolution of native species to increase their competitive tolerance and thus reduce their sensitivity to bioinvasions [57]. As the community size increased, the proportion of species with extreme specialization also increased, and the difference in their niche breadths was further amplified. Compared to terrestrial communities, aquatic communities that have simpler structures and lower species diversity are more vulnerable to plant invasions [10,32]. A. philoxeroides has extensively invaded various aquatic habitats, and tends to form monodominant populations, which results in the niche breadths of the aquatic accompanying species having no obvious latitudinal trend.
4.3. Causes of Niche Similarity and Overlap Variations along the Latitudinal Gradient in Heterogeneous Habitats
In the terrestrial community, the niche similarity between A. philoxeroides and its accompanying species at low latitudes was relatively high, indicating that the accompanying species tended to possess climate adaptability similar to that of A. philoxeroides, and thermal stability promoted the coevolution of plant species in the low-latitude regions. Each accompanying species also had high niche similarity for preferably adapting to the coexistence environment, which was conducive to the formation of higher species diversity at low latitudes [58,59], and environmental filtering led to obvious aggregation in these regions [19]. A. philoxeroides has high phenotypic plasticity and compensatory growth capacity at high latitudes [39], but the cold climate at high latitudes limits the growth of native species. Only a few species with strong environmental tolerance can share resources with A. philoxeroides and thus have higher niche similarity, and most of the remaining species pairs have low niche similarity [45]. The niche similarity between A. philoxeroides and its accompanying species in the aquatic habitats had no obvious latitudinal trend because the water bodies buffered temperature fluctuations caused by latitude changes; thus, they were more stable than terrestrial habitats [60]. However, A. philoxeroides invasion was more significant in aquatic habitats, and the accompanying species thus suffered from stronger competitive exclusion [35].
A. philoxeroides had high niche overlap with its accompanying species in the heterogeneous habitats, indicating that it had a higher competitive advantage than native species. In terrestrial habitats, the niche overlap between A. philoxeroides and its accompanying species significantly increased with increasing latitude, particularly in latitudinal Cluster 7, which may imply that A. philoxeroides strengthened competition with its accompanying species by increasing the niche overlap under increased environmental stress; in turn, the resource utilization of the accompanying species decreased, reducing their adaptability to harsh environments and thus inhibiting native plant establishment at high latitudes [58,59]. However, the niche overlap between A. philoxeroides and its accompanying species in aquatic habitats had no obvious latitudinal trend, possibly due to aquatic plant species experiencing severe A. philoxeroides invasion at all latitudinal clusters [61]. In this study, niche overlap between A. philoxeroides and P. stratiotes and E. crassipes was higher at low latitudes; these are all globally invasive weeds with vegetative propagation, indicating that they may have similar survival needs for dissolved oxygen, light and water flow and may also provide interspecific mutual assistance [62,63]. Attention must be paid to this phenomenon of collaborative aquatic plant invasions at low latitudes because with rapid global warming, the superior hydrothermal conditions at low latitudes will accelerate the niche expansion and coevolution of aquatic invasive plants originating from tropical America [61,64]. In addition, there were over 10 forms and varieties of A. philoxeroides in the natural ecosystems, which made the taxonomy of A. philoxeroides one of the most complicated taxa among Alternanthera [65,66]; thus, it is necessary to fully assess the influence of taxonomic differences of A. philoxeroides on its niche characteristics in future studies.
5. Conclusions
We found that, of the studied species, A. philoxeroides had the greatest niche breadth in both aquatic and terrestrial habitats and that the niche similarity and overlap between A. philoxeroides and its accompanying species were all high. The niche similarity of A. philoxeroides in the terrestrial communities was significantly higher than that in the aquatic communities, while the niche similarity and overlap between each accompanying species were low. The niche breadth of A. philoxeroides had no obvious latitudinal trend, which was attributed to its strong environmental tolerance. In the terrestrial communities, the niche similarity of A. philoxeroides and the niche breadth of its accompanying species all significantly decreased with increasing latitude, while the niche overlap between A. philoxeroides and its accompanying species significantly increased with increasing latitude. However, the niche characteristics in the aquatic communities had no obvious latitudinal trend. These findings indicate that habitat heterogeneity significantly affects the niche characteristics of the A. philoxeroides community and that A. philoxeroides invasion might weaken the interspecific competition of its accompanying species. At low latitudes, environmental filtering resulted in species with similar niches in the A. philoxeroides communities showing obvious aggregation, and the invasive and native species tended to evolve similar environmental capacities. At high latitudes, the restrictions of the cold climate led to an increase in niche overlap between A. philoxeroides and its accompanying species.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/d15050651/s1, Table S1: Sampling plots of A. philoxeroides-invaded communities in terrestrial and aquatic habitats; Table S2: The functional traits of species with high-frequency in terrestrial and aquatic A. philoxeroides-invaded communities.
Author Contributions
Conceptualization, H.W. and B.R.; methodology, H.W.; formal analysis, H.W. and S.D.; investigation, H.W., S.D., Y.W. and L.W.; data curation, H.W. and S.D.; writing—original draft preparation, H.W. and S.D.; writing—review and editing, H.W., S.D. and B.R.; funding acquisition, H.W. and B.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The National Natural Science Foundation of China (31800460), The Xinyang Academy of Ecological Research Open Foundation (2023XYMS16) and The Nanhu Scholars Program for Young Scholars of Xinyang Normal University (XYNU) (2023A017).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available upon request.
Acknowledgments
We would like to thank Shaoqi Jia, Junda Wang, Qiubo Ji, Nannan Xiao and Wenhao Wang for their help in field surveys.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Carboni, M.; Livingstone, S.W.; Isaac, M.E.; Cadotte, M.W. Invasion drives plant diversity loss through competition and ecosystem modification. J. Ecol. 2021, 109, 3587–3601. [Google Scholar] [CrossRef]
- Pabst, R.; Dias, F.S.; Borda-de-Água, L.; Rodríguez-González, P.M.; Capinha, C. Assessing and predicting the distribution of riparian invasive plants in continental Portugal. Front. Ecol. Evol. 2022, 10, 875578. [Google Scholar] [CrossRef]
- Lebbink, G.; Dwyer, J.; Fensham, R. An invasive grass species has both local and broad-scale impacts on diversity: Potential mechanisms and implications. J. Veg. Sci. 2021, 32, 1240–1253. [Google Scholar] [CrossRef]
- Yue, M.; Shen, H.; Ye, W.; Li, W.; Chen, J. Winter low temperature disturbance in the southern subtropics of China promotes the competitiveness of an invasive plant. Biol. Invasions 2021, 23, 2913–2925. [Google Scholar] [CrossRef]
- Gross, N.; Liancourt, P.; Butters, R.; Duncan, R.; Hulme, P.E. Functional equivalence, competitive hierarchy and facilitation determine species coexistence in highly invaded grasslands. New Phytol. 2015, 206, 175–186. [Google Scholar] [CrossRef]
- Wang, C.; Yu, Y.; Cheng, H.; Du, D. Which factor contributes most to the invasion resistance of native plant communities under the co-invasion of two invasive plant species? Sci. Total Environ. 2022, 813, 152628. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J.Q. Abiotic and biotic determinants of plant diversity in aquatic communities invaded by water hyacinth (Eichhornia crassipes (Mart.) Solms). Front. Plant Sci. 2020, 11, 1306. [Google Scholar] [CrossRef]
- Dubey, D.; Kumar, S.; Dutta, V. In-situ anatomical and elemental response of aquatic macrophytes against nutrient enrichment in freshwater tropical lakes. Appl. Water Sci. 2022, 12, 163. [Google Scholar] [CrossRef]
- Huang, X.; Ke, F.; Li, Q.; Zhao, Y.; Guan, B.; Li, K. Functional traits underlying performance variations in the overwintering of the cosmopolitan invasive plant water hyacinth (Eichhornia crassipes) under climate warming and water drawdown. Ecol. Evol. 2022, 12, e9181. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A. Invasive non-native plants have a greater effect on neighbouring natives than other non-natives. Nat. Plants 2016, 2, 1613. [Google Scholar] [CrossRef]
- Byers, J.E.; Noonburg, E.G. Scale dependent effects of biotic resistance. Ecology 2003, 84, 1428–1433. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Von Holle, B. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Powell, K.I.; Chase, J.M.; Knight, T.M. Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science 2013, 339, 316–318. [Google Scholar] [CrossRef]
- Tilman, D. Niche tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 10854–10861. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Brönnimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef]
- Cai, Q.; Welk, E.; Ji, C.; Fang, W.; Sabatini, F.M.; Zhu, J.; Zhu, J.; Tang, Z.; Attorre, F.; Campos, J.A.; et al. The relationship between niche breadth and range size of beech (Fagus) species worldwide. J. Biogeogr. 2021, 48, 1240–1253. [Google Scholar] [CrossRef]
- Pastore, A.I.; Barabás, G.; Bimler, M.D.; Mayfield, M.M.; Miller, T.E. The evolution of niche overlap and competitive differences. Nat. Ecol. Evol. 2021, 5, 330–337. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Tucker, C.M. Should Environmental Filtering be Abandoned? Trends Ecol. Evol. 2017, 32, 429–437. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Courchamp, F.; Roura-Pascual, N.; Jeschke, J.M. Biological invasions reveal how niche change affects the transferability of species distribution models. Ecology 2022, 103, e3719. [Google Scholar] [CrossRef]
- Yuan, Y.; Tang, X.; Liu, M.; Liu, X.; Tao, J. Species distribution models of the Spartina alterniflora Loisel in its origin and invasive country reveal an ecological niche shift. Front. Plant Sci. 2021, 12, 738769. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D. Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 2012, 485, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Schweiger, O.; Ingolf, K. Niche expansion of the invasive plant species Ageratina adenophora despite evolutionary constraints. J. Biogeogr. 2019, 46, 1306–1315. [Google Scholar]
- Enders, M.; Havemann, F.; Ruland, F.; Bernard-Verdier, M.; Catford, J.A.; Gómez-Aparicio, L.; Haider, S.; Heger, T.; Kueffer, C.; Kühn, I.; et al. A conceptual map of invasion biology: Integrating hypotheses into a consensus network. Glob. Ecol. Biogeogr. 2020, 29, 978–991. [Google Scholar] [CrossRef]
- Yang, Y.H.; Bao, L.P. Scale-dependent changes in species richness caused by invader competition. Ecol. Model. 2022, 469, 109996. [Google Scholar] [CrossRef]
- Sexton, J.P.; Montiel, J.; Shay, J.E.; Stephens, M.R.; Slatyer, R.A. Evolution of ecological niche breadth. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 183–206. [Google Scholar] [CrossRef]
- Finch, J.; Walck, J.L.; Hidayati, S.N.; Kramer, A.T.; Lason, V.; Havens, K. Germination niche breadth varies inconsistently among three Asclepias congeners along a latitudinal gradient. Plant Biol. 2019, 21, 425–438. [Google Scholar] [CrossRef]
- Schuster, M.J.; Wragg, P.D.; Reich, P.B. Phenological niche overlap between invasive buckthorn (Rhamnus cathartica) and native woody species. For. Ecol. Manag. 2021, 498, 11956. [Google Scholar] [CrossRef]
- Stotz, G.C.; Pec, G.J.; Cahill, J.F. Is biotic resistance to invaders dependent upon local environmental conditions or primary productivity? A meta-analysis. Basic Appl. Ecol. 2016, 17, 377–387. [Google Scholar] [CrossRef]
- Welshofer, K.B.; Zarnetske, P.L.; Lany, N.K.; Read, Q.D. Short-term responses to warming vary between native vs. exotic species and with latitude in an early successional plant community. Oecologia 2018, 187, 333–342. [Google Scholar] [CrossRef]
- Cirtwill, A.R.; Stouffer, D.B.; Romanuk, T.N. Latitudinal gradients in biotic niche breadth vary across ecosystem types. Proc. R. Soc. B-Biol. Sci. 2015, 282, 20151589. [Google Scholar] [CrossRef]
- Vázquez, D.P.; Stevens, R.D. The latitudinal gradient in niche breadth: Concepts and evidence. Am. Nat. 2004, 164, E1–E19. [Google Scholar] [CrossRef]
- Alahuhta, J.; Virtala, A.; Hjort, J.; Ecke, F.; Johnson, L.B.; Sass, L.; Heino, J. Average niche breadths of species in lake macrophyte communities respond to ecological gradients variably in four regions on two continents. Oecologia 2017, 184, 219–235. [Google Scholar] [CrossRef]
- Wu, H.; Dong, S.J.; Rao, B.Q. Latitudinal trends in the structure, similarity and beta diversity of plant communities invaded by Alternanthera philoxeroides in heterogeneous habitats. Front. Plant Sci. 2022, 13, 1021337. [Google Scholar] [CrossRef]
- Wu, H.; Carrillo, J.; Ding, J.Q. Species diversity and environmental determinants of aquatic and terrestrial communities invaded by Alternanthera philoxeroides. Sci. Total Environ. 2017, 581, 666–675. [Google Scholar] [CrossRef]
- Lu, X.M.; Siemann, E.; Shao, X.; Wei, H.; Ding, J. Climate warming affects biological invasions by shifting interactions of plants and herbivores. Glob. Chang. Biol. 2013, 19, 2339–2347. [Google Scholar] [CrossRef]
- Wu, H.; Ismail, M.; Ding, J.Q. Global warming increases the interspecific competitiveness of the invasive plant alligator weed, Alternanthera philoxeroides. Sci. Total Environ. 2017, 575, 1415–1422. [Google Scholar] [CrossRef]
- Wang, T.; Hu, J.; Wang, R.; Liu, C.; Yu, D. Tolerance and resistance facilitate the invasion success of Alternanthera philoxeroides in disturbed habitats: A reconsideration of the disturbance hypothesis in the light of phenotypic variation. Environ. Exp. Bot. 2018, 153, 135–142. [Google Scholar] [CrossRef]
- Wu, H.; Carrillo, J.; Ding, J.Q. Invasion by alligator weed, Alternanthera philoxeroides, is associated with decreased species diversity across the latitudinal gradient in China. J. Plant Ecol. 2016, 93, 311–319. [Google Scholar] [CrossRef]
- Lu, X.M.; Siemann, E.; He, M.Y.; Wei, H.; Shao, X.; Ding, J. Climate warming increases biological control agent impact on a non-target species. Ecol. Lett. 2015, 18, 48–56. [Google Scholar] [CrossRef]
- Ma, Y.; Li, Q.; Pan, S.; Liu, C.; Han, M.; Brancelj, A. Niche and interspecific associations of Pseudoanabaena limnetica-Exploring the influencing factors of its succession stage. Ecol. Indic. 2022, 138, 108806. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef]
- Martorell, C.; Almanza-Celis, C.A.; Pérez-García, E.A.; Sánchez-Ken, J.G. Co-existence in a species-rich grassland: Competition, facilitation and niche structure over a soil depth gradient. J. Veg. Sci. 2015, 26, 674–685. [Google Scholar] [CrossRef]
- Luo, L.; Kong, X.; Gao, Z.; Zheng, Y.; Yang, Y.; Li, X.; Yang, D.; Geng, Y.; Yang, Y. Comparative transcriptome analysis reveals ecological adaption of cold tolerance in northward invasion of Alternanthera philoxeroides. BMC Genom. 2020, 21, 532. [Google Scholar] [CrossRef] [PubMed]
- Stotz, G.C.; Salgado-Luarte, C.; Escobedo, V.M.; Valladares, F.; Gianoli, E. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef]
- Müller, C.; Junker, R.R. Chemical phenotype as important and dynamic niche dimension of plants. New Phytol. 2022, 234, 1168–1174. [Google Scholar] [CrossRef]
- Lancaster, L.T. On the macroecological significance of eco-evolutionary dynamics: The range shift-niche breadth hypothesis. Philos. Trans. R. Soc. B-Biol. Sci. 2022, 377, 20210013. [Google Scholar] [CrossRef]
- Bar-Massada, A. Complex relationships between species niches and environmental heterogeneity affect species co-occurrence patterns in modelled and real communities. Proc. R. Soc. B-Biol. Sci. 2015, 282, 20150927. [Google Scholar] [CrossRef]
- Segura, A.M.; Calliari, D.; Kruk, C.; Conde, D.; Bonilla, S.; Fort, H. Emergent neutrality drives phytoplankton species coexistence. Proc. R. Soc. B-Biol. Sci. 2011, 278, 2355–2361. [Google Scholar] [CrossRef]
- Pennington, R.T.; Richardson, J.E.; Lavin, M. Insights into the historical construction of species-rich biomes from dated plant phylogenies, neutral ecological theory and phylogenetic community structure. New Phytol. 2006, 172, 605–616. [Google Scholar] [CrossRef]
- dos Santos Ribas, L.G.; de Cassia-Silva, C.; Petsch, D.K.; Silveira, M.L.; Lima-Ribeiro, M.S. The potential invasiveness of an aquatic macrophyte reflects founder effects from native niche. Biol. Invasions 2018, 20, 3347–3355. [Google Scholar] [CrossRef]
- He, D.; Chen, Y.; Zhao, K.; Cornelissen, J.H.C.; Chu, C. Intra- and interspecific trait variations reveal functional relationships between specific leaf area and soil niche within a subtropical forest. Ann. Bot. 2018, 121, 1173–1182. [Google Scholar] [CrossRef]
- van Boheemen, L.A.; Atwater, D.Z.; Hodgins, K.A. Rapid and repeated local adaptation to climate in an invasive plant. New Phytol. 2019, 222, 614–627. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, M.; Pan, Y.; Huang, H.; Pan, X.; Sosa, A.; Hou, Y.; Zhu, Z.; Li, B. Rapid evolution of latitudinal clines in growth and defence of an invasive weed. New Phytol. 2021, 230, 845–856. [Google Scholar] [CrossRef]
- Morin, X.; Chuine, I. Niche breadth, competitive strength and range size of tree species: A trade-off based framework to understand species distribution. Ecol. Lett. 2006, 9, 185–195. [Google Scholar] [CrossRef]
- Foster, S.L.; Kharouba, H.M.; Kharouba, H.M. Testing the assumption of environmental equilibrium in an invasive plant species over a 130 year history. Ecography 2022, 2022, e06284. [Google Scholar] [CrossRef]
- Golivets, M.; Wallin, K.F. Neighbour tolerance, not suppression, provides competitive advantage to non-native plants. Ecol. Lett. 2018, 21, 745–759. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Gilbert, B.; Levine, J.M. Plant invasions and the niche. J. Ecol. 2009, 97, 609–615. [Google Scholar] [CrossRef]
- Carscadden, K.A.; Emery, N.C.; Arnillas, C.A.; Cadotte, M.W.; Afkhami, M.E.; Gravel, D.; Livingstone, S.W.; Wiens, J.J. Niche breadth: Causes and consequences for ecology, evolution, and conservation. Q. Rev. Biol. 2020, 95, 179–214. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; Shadrin, N.V. Extreme hydrological events destabilize aquatic ecosystems and open doors for alien species. Quat. Int. 2018, 475, 11–15. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J.Q. Global change sharpens the double-edged sword of aquatic alien plants in China and beyond. Front. Plant Sci. 2019, 10, 787. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Brunel, S. Assessing and managing the current and future pest risk from water hyacinth, (Eichhornia crassipes), an invasive aquatic plant threatening the environment and water security. PLoS ONE 2016, 11, e0120054. [Google Scholar] [CrossRef] [PubMed]
- Jawed, A.; Kar, P.; Verma, R.; Shukla, K.; Hemanth, P.; Thakur, V.K.; Pandey, L.M.; Gupta, R.K. Integration of biological control with engineered heterojunction nano-photocatalysts for sustainable and effective management of water hyacinth weed. J. Environ. Chem. Eng. 2022, 10, 106976. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Liang, X.R.; Harms, N.E.; Tan, F.; Lin, Y.; Feng, H.; Wang, J.; Li, Q.; Jia, Y.; Lu, X.; et al. Spatio-temporal pattern of cross-continental invasion: Evidence of climatic niche shift and predicted range expansion provide management insights for smooth cordgrass. Ecol. Indic. 2022, 140, 109052. [Google Scholar] [CrossRef]
- Jia, X.; Pan, X.-Y.; Sosa, A.; Li, B.; Chen, J. Differentiation in growth and biomass allocation among three native Alternanthera philoxeroides varieties from Argentina. Plant Species Biol. 2010, 25, 85–92. [Google Scholar] [CrossRef]
- Jiang, P.; Xu, G.; He, Y.; Sun, T.; Liu, C.; Chen, C.; Zuo, R.; Sun, C. Complete chloroplast genome of Alternanthera philoxeroides by de novo sequencing. Mitochondrial DNA Part B-Resour. 2021, 6, 1826–1828. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).