
Looking at the Expansion of Three Demersal Lessepsian Fish Immigrants in the Greek Seas: What Can We Get from Spatial Distribution Modeling?



Abstract
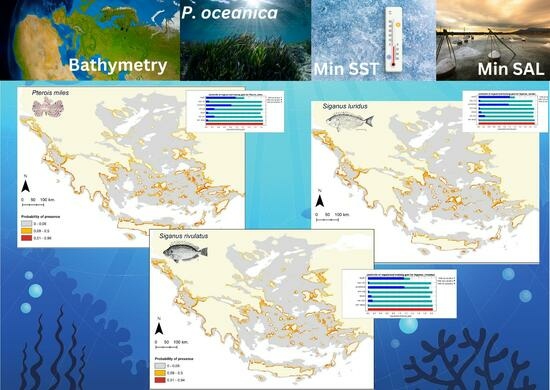
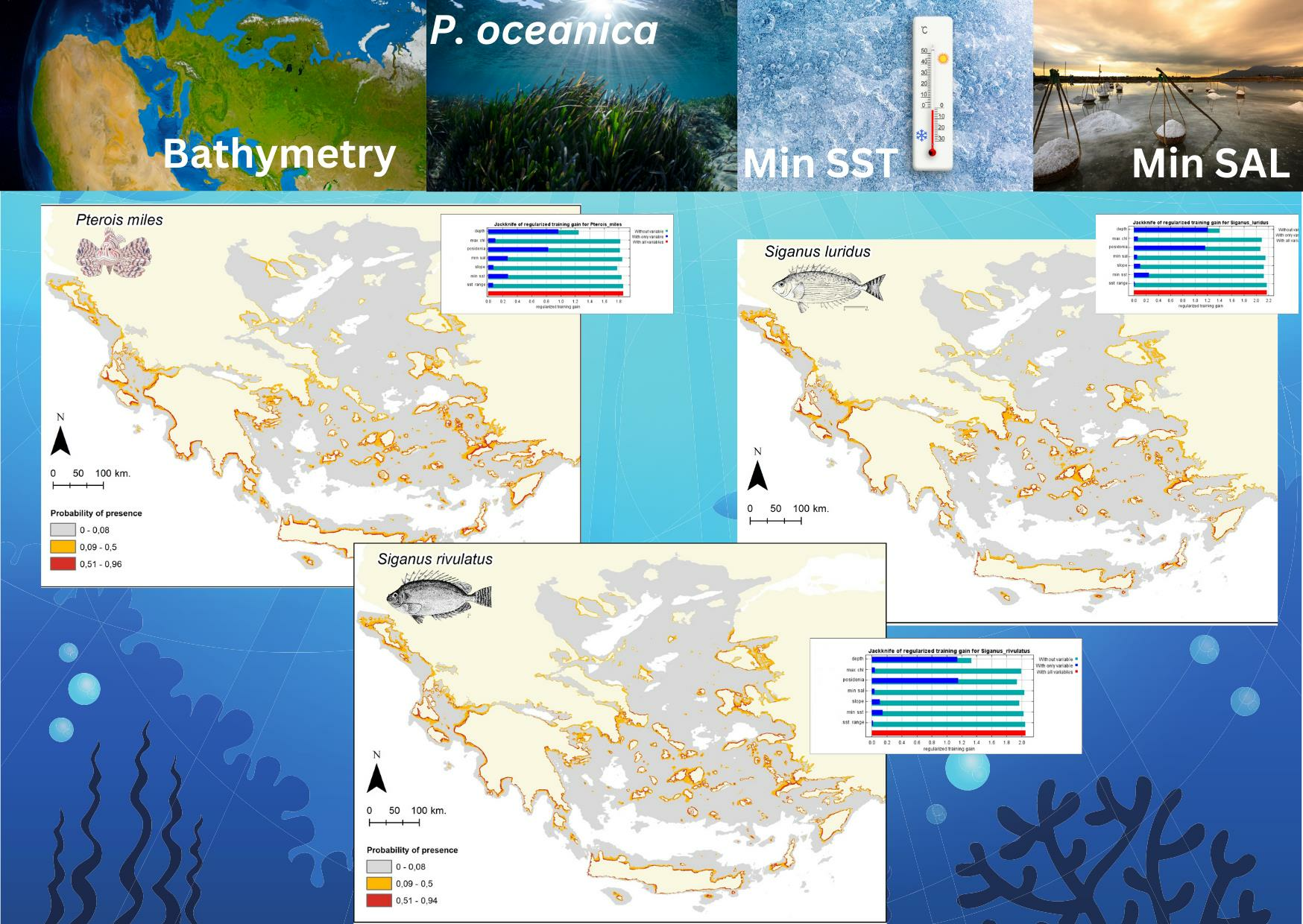{kind=link}
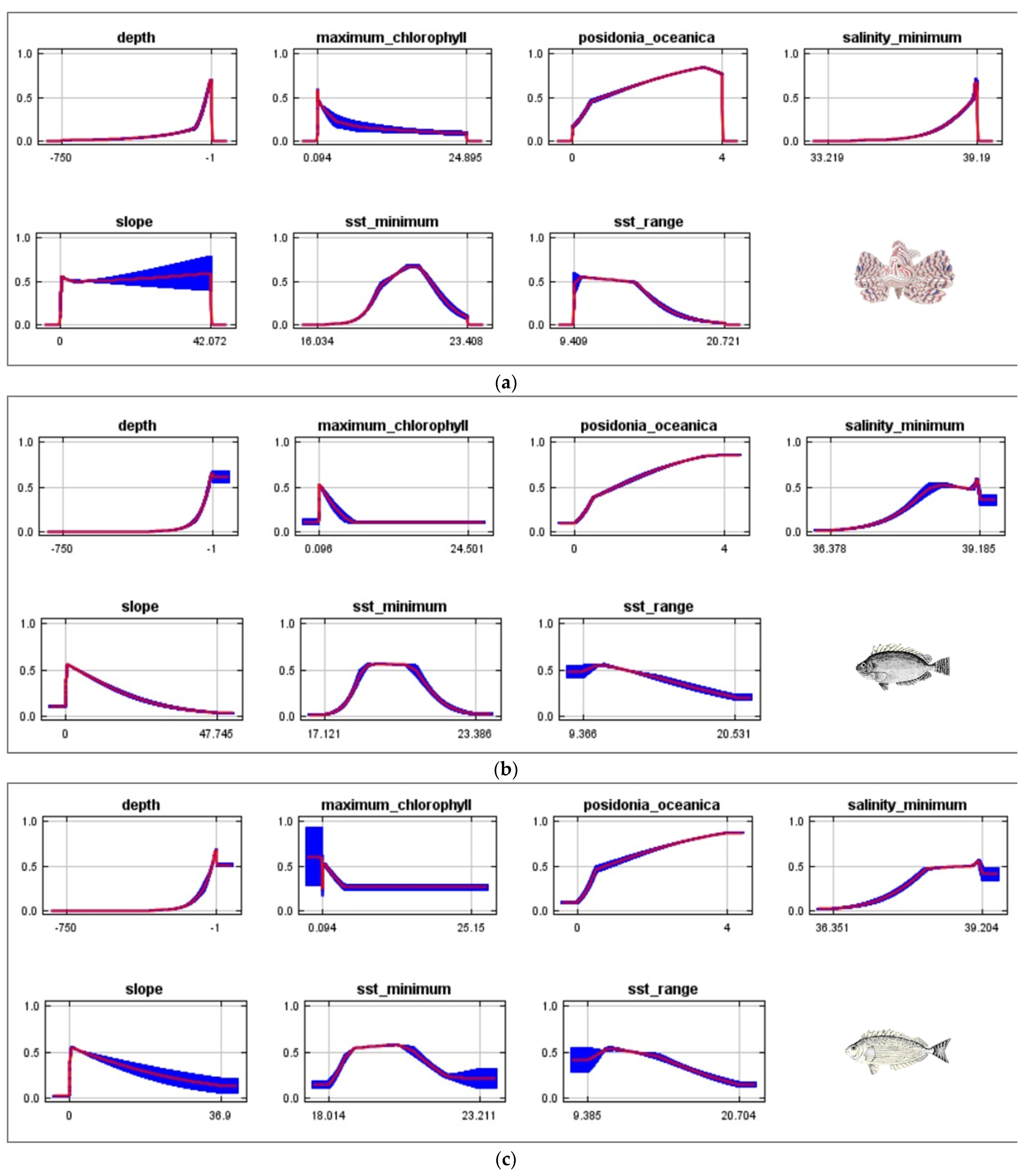{kind=link}
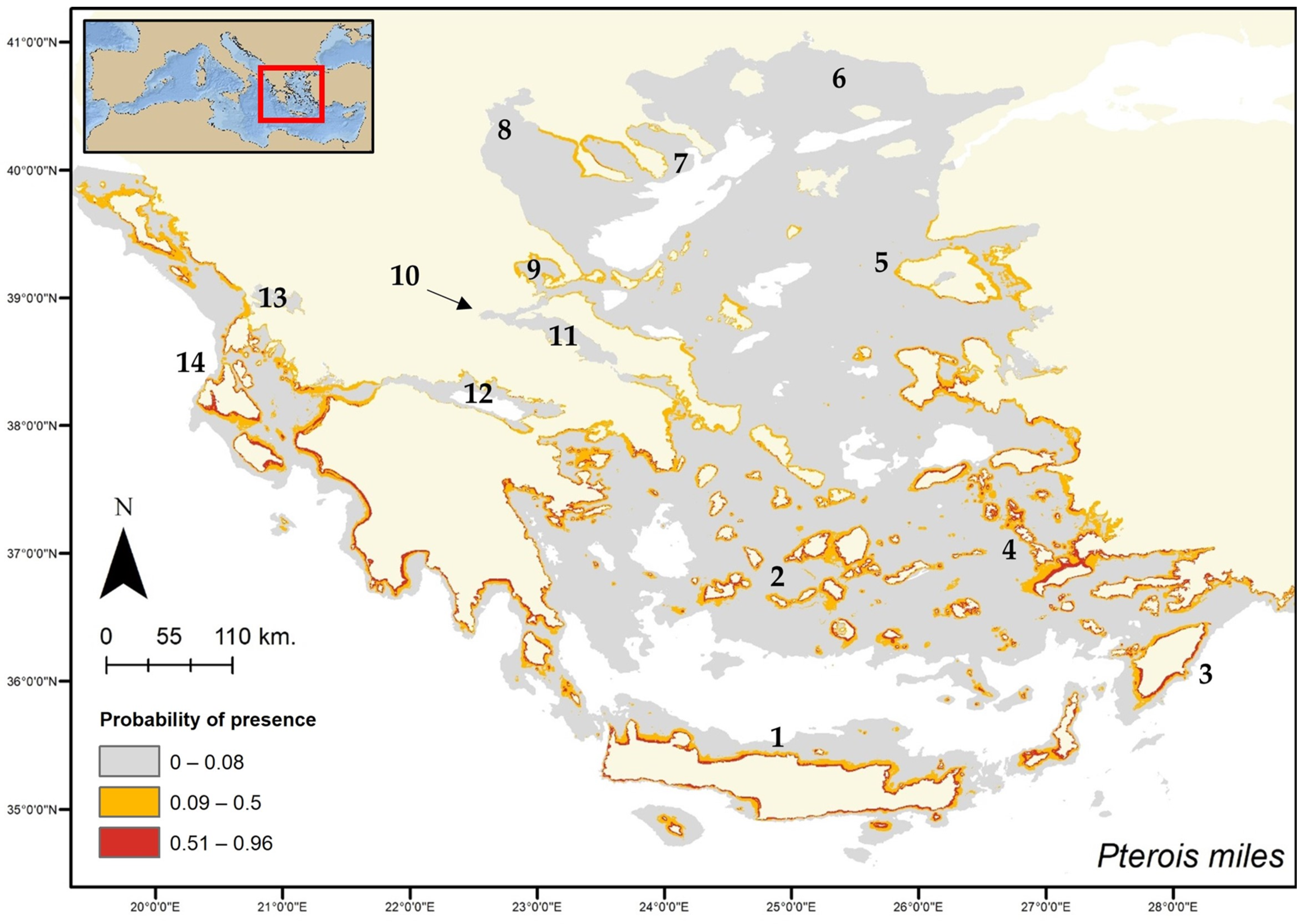{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
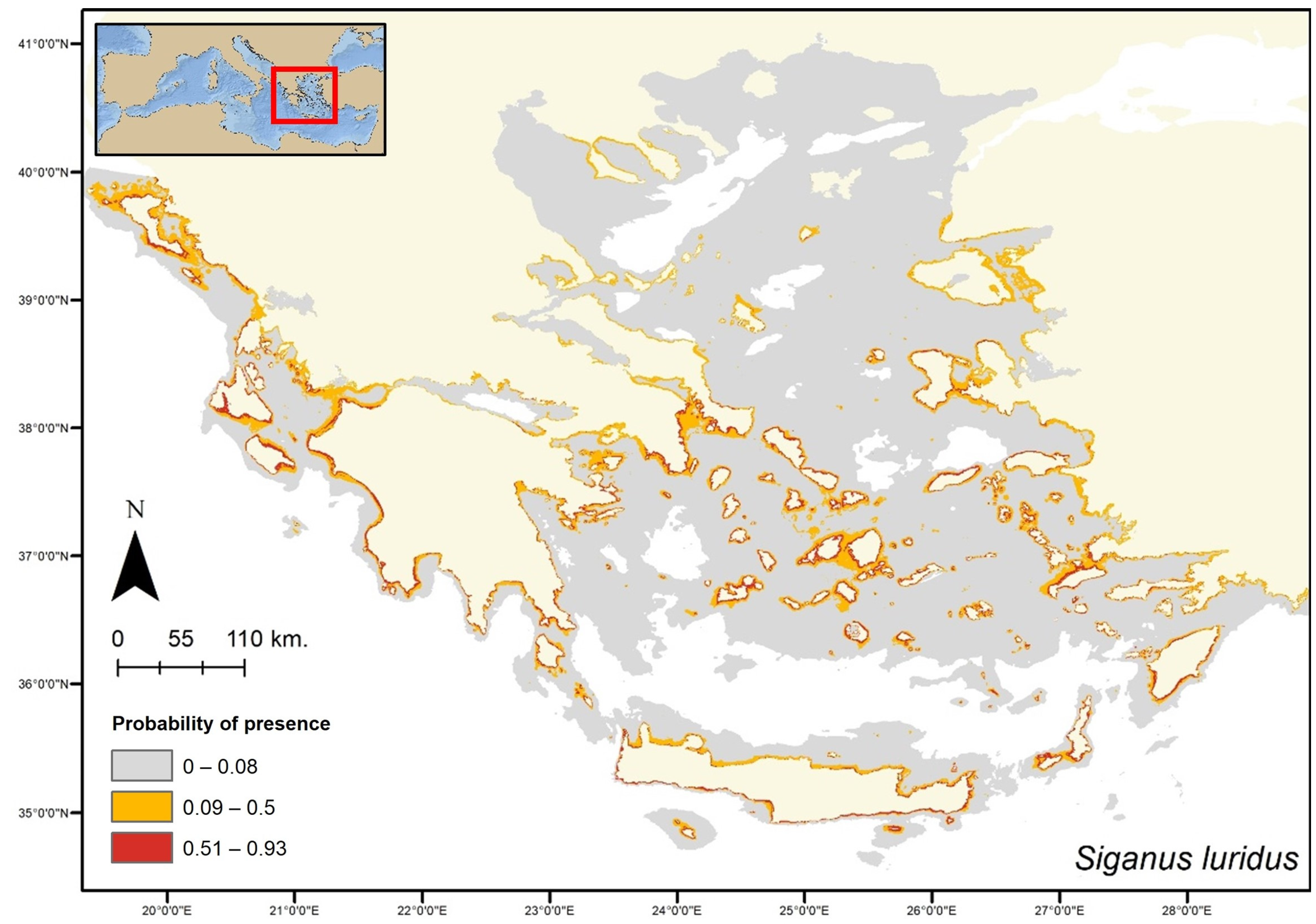
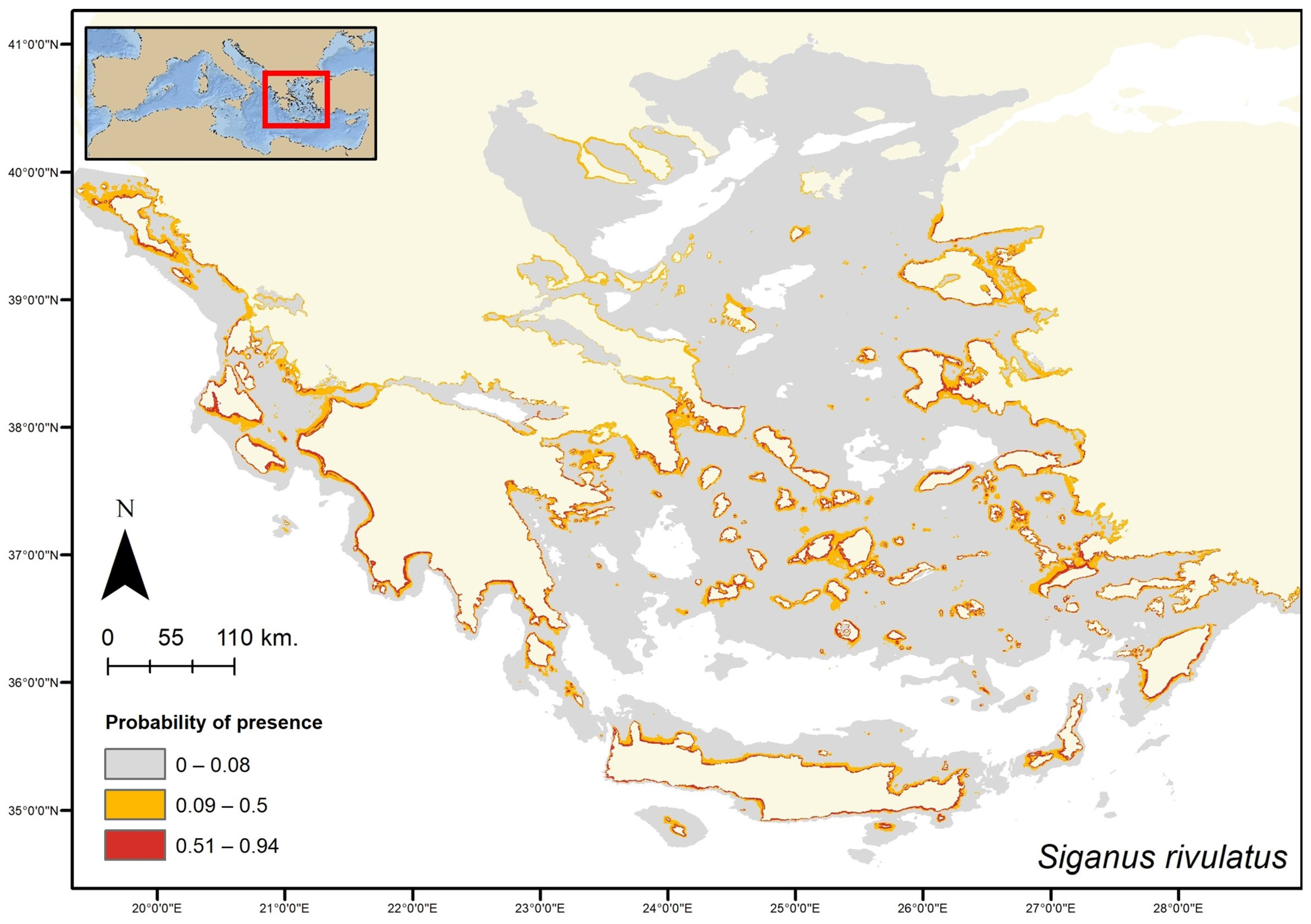
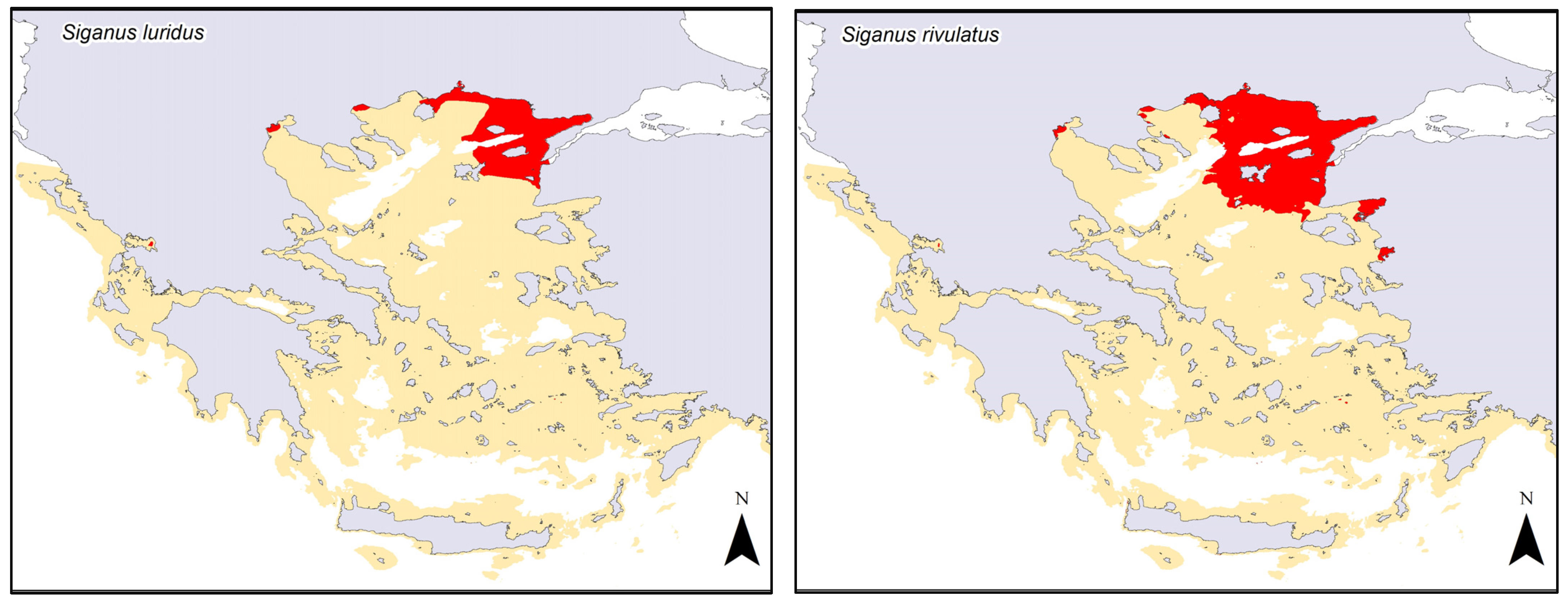
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Golani, D. Impact of Red Sea fish migrants through the Suez Canal on the aquatic environment of the Eastern Mediterranean. Bull. Ser. Yale Sch. For. Environ. Stud. 1998, 103, 375–387. [Google Scholar]
- Edelist, D.; Rilov, G.; Golani, D.; Carlton, J.T.; Spanier, E. Restructuring the Sea: Profound shifts in the world’s most invaded marine ecosystem. Divers. Distrib. 2013, 19, 69–77. [Google Scholar] [CrossRef]
- Karachle, P.K.; Oikonomou, A.; Pantazi, M.; Stergiou, K.I.; Zenetos, A. Can Biological Traits Serve as Predictors for Fishes’ Introductions, Establishment, and Interactions? The Mediterranean Sea as a Case Study. Biology 2022, 11, 1625. [Google Scholar] [CrossRef] [PubMed]
- Por, F.D. Lessepsian Migration: The Influx of Red Sea Biota into the Mediterranean by Way of the Suez Canal; Ecological Studies 23; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1978. [Google Scholar]
- Galanidi, M.; Zenetos, A. Data-Driven Recommendations for Establishing Threshold Values for the NIS Trend Indicator in the Mediterranean Sea. Diversity 2022, 14, 57. [Google Scholar] [CrossRef]
- Galil, B.S.; Danovaro, R.; Rothman, S.B.S.; Gevili, R.; Goren, M. Invasive biota in the deep-sea Mediterranean: An emerging issue in marine conservation and management. Biol. Invasions 2019, 21, 281–288. [Google Scholar] [CrossRef]
- Kalogirou, S.; Corsini-Foka, M.; Sioulas, A.; Wennhage, H.; Pihl, L. Diversity, structure and function of fish assemblages associated with Posidonia oceanica beds in an area of the eastern Mediterranean Sea and the role of non-indigenous species. J. Fish Biol. 2010, 77, 2338–2357. [Google Scholar] [CrossRef]
- Öndes, F.; Ünal, V. The dominance of non-indigenous species in the catch composition of small-scale fisheries: A case study from the Kaş–Kekova Special Environmental Protection Area, Türkiye, Eastern Mediterranean. Acta Ichthyol. Et Piscat. 2023, 53, 27–35. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; Garcia, E.L.; Stern, N.; Tsiamis, K.; Galanidi, M. Established non-indigenous species increased by 40% in 11 years in the Mediterranean Sea. Mediterr. Mar. Sci. 2022, 23, 196–212. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Bailey, S.A. An overview of thirty years of research on ballast water as a vector for aquatic invasive species to freshwater and marine environments. Aquat. Ecosyst. Health Manag. 2015, 18, 261–268. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Givan, O.; Parravicini, V.; Kulbicki, M.; Belmaker, J. Trait structure reveals the processes underlying fish establishment in the Mediterranean. Glob. Ecol. Biogeogr. 2017, 26, 142–153. [Google Scholar] [CrossRef]
- Savva, I.; Chartosia, N.; Antoniou, C.; Kleitou, P.; Georgiou, A.; Stern, N.; Hadjioannou, L.; Jimenez, C.; Andreou, V.; Hall-Spencer, J.M.; et al. They are here to stay: The biology and ecology of lionfish (Pterois miles) in the Mediterranean Sea. J. Fish Biol. 2020, 97, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Ulman, A.; Harris, H.E.; Doumpas, N.; Deniz Akbora, H.; Mabruk, A.; Azzurro, E.; Bariche, M.; Çiçek, B.A.; Deidun, A.; Demirel; et al. Low pufferfish and lionfish predation in their native and invaded ranges suggests human control mechanisms may be necessary to control their Mediterranean abundances. Front. Mar. Sci. 2021, 8, 670413. [Google Scholar] [CrossRef]
- Poore, A.G.; Campbell, A.H.; Coleman, R.A.; Edgar, G.J.; Jormalainen, V.; Reynolds, P.L.; Sotka, E.E.; Stachowicz, J.J.; Taylor, R.B.; Vanderklift, M.A.; et al. Global patterns in the impact of marine herbivores on benthic primary producers. Ecol. Lett. 2012, 15, 912–922. [Google Scholar] [CrossRef]
- Sala, E.; Kizilkaya, Z.; Yildirim, D.; Ballesteros, E. Alien marine fishes deplete algal biomass in the eastern Mediterranean. PLoS ONE 2011, 6, e17356. [Google Scholar] [CrossRef]
- Giakoumi, S. Distribution patterns of the invasive herbivore Siganus luridus (Rüppell, 1829) and its relation to native benthic communities in the central Aegean Sea, Northeastern Mediterranean. Mar. Ecol. 2014, 35, 96–105. [Google Scholar] [CrossRef]
- Dimitriadis, C.; Galanidi, M.; Zenetos, A.; Corsini-Foka, M.; Giovos, I.; Karachle, P.K.; Fournari–Konstantinidou, I.; Kytinou, E.; Issaris, Y.; Azzurro, E.; et al. Updating the occurrences of Pterois miles in the Mediterranean Sea, with considerations on thermal boundaries and future range expansion. Mediterr. Mar. Sci. 2020, 21, 62–69. [Google Scholar] [CrossRef]
- Poursanidis, D.; Kougioumoutzis, K.; Minasidis, V.; Chartosia, N.; Kletou, D.; Kalogirou, S. Uncertainty in Marine Species Distribution Modelling: Trying to Locate Invasion Hotspots for Pterois miles in the Eastern Mediterranean Sea. J. Mar. Sci. Eng. 2022, 10, 729. [Google Scholar] [CrossRef]
- Dragičević, B.; Ugarković, P.; Krželj, M.; Zurub, D.; Dulčić, J. New record of Pterois cf. miles (Actinopterygii: Scorpaeniformes: Scorpaenidae) from the eastern middle Adriatic Sea (Croatian waters): Northward expansion. Acta Ichthyol. Et Piscat. 2021, 51, 379–383. [Google Scholar] [CrossRef]
- Schofield, P.J. Update on geographic spread of invasive lionfishes (Pterois volitans [Linnaeus, 1758] and P. miles [Bennett, 1828]) in the Western North Atlantic Ocean, Caribbean Sea and Gulf of Mexico. Aquat. Invasions 2010, 5 (Suppl. 1), S117–S122. [Google Scholar] [CrossRef]
- Azzurro, E.; Stancanelli, B.; Di Martino, V.; Bariche, M. Range expansion of the common lionfish Pterois miles (Bennett, 1828) in the Mediterranean Sea: An unwanted new guest for Italian waters. BioInvasions Rec. 2017, 6, 95–98. [Google Scholar] [CrossRef]
- Côté, I.M.; Smith, N.S. The lionfish Pterois sp. invasion: Has the worst-case scenario come to pass? J. Fish Biol. 2018, 92, 660–689. [Google Scholar] [CrossRef]
- Muñoz, R.C.; Currin, C.A.; Whitfield, P.E. Diet of invasive lionfish on hard bottom reefs of the Southeast USA: Insights from stomach contents and stable isotopes. Mar. Ecol. Prog. Ser. 2011, 432, 181–193. [Google Scholar] [CrossRef]
- Corsini-Foka, M.; Mastis, S.; Kondylatos, G.; Batjakas, I.E. Alien and native fish in gill nets at Rhodes, eastern Mediterranean (2014–2015). J. Mar. Biol. Assoc. United Kingd. 2017, 97, 635–642. [Google Scholar] [CrossRef]
- Kleitou, P.; Moutopoulos, D.K.; Giovos, I.; Kletou, D.; Savva, I.; Cai, L.L.; Hall-Spencer, J.M.; Charitou, A.; Elia, M.; Katselis, G.; et al. Conflicting interests and growing importance of non-indigenous species in commercial and recreational fisheries of the Mediterranean Sea. Fish. Manag. Ecol. 2022, 29, 169–182. [Google Scholar] [CrossRef]
- Meadows, B.; Sims, C. Can We Love Invasive Species to Death? Creating Efficient Markets for Invasive Species Harvests. Environ. Resour. Econ. 2023, 85, 443–477. [Google Scholar] [CrossRef]
- Parravicini, V.; Azzurro, E.; Kulbicki, M.; Belmaker, J. Niche shift can impair the ability to predict invasion risk in the marine realm: An illustration using Mediterranean fish invaders. Ecol. Lett. 2015, 18, 246–253. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- D’Amen, M.; Azzurro, E. Lessepsian fish invasion in Mediterranean marine protected areas: A risk assessment under climate change scenarios. ICES J. Mar. Sci. 2020, 77, 388–397. [Google Scholar] [CrossRef]
- Turan, C. Species distribution modelling of invasive alien species; Pterois miles for current distribution and future suitable habitats. Glob. J. Environ. Sci. Manag. 2020, 6, 429–440. [Google Scholar] [CrossRef]
- D’Amen, M.; Azzurro, E. Integrating univariate niche dynamics in species distribution models: A step forward for marine research on biological invasions. J. Biogeogr. 2020, 47, 686–697. [Google Scholar] [CrossRef]
- Azzurro, E.; Soto, S.; Garofalo, G.; Maynou, F. Fistularia commersonii in the Mediterranean Sea: Invasion history and distribution modeling based on presence-only records. Biol. Invasions 2013, 15, 977–990. [Google Scholar] [CrossRef]
- Coro, G.; Vilas, L.G.; Magliozzi, C.; Ellenbroek, A.; Scarponi, P.; Pagano, P. Forecasting the ongoing invasion of Lagocephalus sceleratus in the Mediterranean Sea. Ecol. Model. 2018, 371, 37–49. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Gallagher, R.V.; Thuiller, W.; Downey, P.O.; Leishman, M.R.; Hughes, L. Different climatic envelopes among invasive populations may lead to underestimations of current and future biological invasions. Divers. Distrib. 2009, 15, 409–420. [Google Scholar] [CrossRef]
- Poursanidis, D.; Kalogirou, S.; Azzurro, E.; Parravicini, V.; Bariche, M.; Zu Dohna, H. Habitat suitability, niche unfilling and the potential spread of Pterois miles in the Mediterranean Sea. Mar. Pollut. Bull. 2020, 154, 111054. [Google Scholar] [CrossRef]
- Vale, C.G.; Tarroso, P.; Brito, J.C. Predicting species distribution at range margins: Testing the effects of study area extent, resolution and threshold selection in the Sahara–Sahel transition zone. Divers. Distrib. 2014, 20, 20–33. [Google Scholar] [CrossRef]
- Zenetos, A.; Arianoutsou, M.; Bazos, I.; Balopoulou, S.; Corsini-Foka, M.; Dimiza, M.; Drakopoulou, P.; Katsanevakis, S.; Kondylatos, G.; Koutsikos, N.; et al. ELNAIS: A collaborative network on aquatic alien species in Hellas (Greece). Manag. Biol. Invasions 2015, 6, 185–196. [Google Scholar] [CrossRef]
- Crocetta, F.; Agius, D.; Balistreri, P.; Bariche, M.; Bayhan, Y.; Çakir, M.; Ciriaco, S.; Corsini-Foka, M.; Deidun, A.; El Zrelli, R.; et al. New Mediterranean Biodiversity Records (October 2015). Mediterr. Mar. Sci. 2015, 16, 682–702. [Google Scholar] [CrossRef]
- Dragicevic, B.; Anadoli, O.; Angel, D.; Benabdi, M.; Bitar, G.; Castriota, L.; Crocetta, F.; Deidun, A.; Dulčić, J.; Edelist, D.; et al. New Mediterranean Biodiversity Records (December 2019). Mediterr. Mar. Sci. 2019, 20, 636–656. [Google Scholar] [CrossRef]
- Thessalou-Legaki, M.; Aydogan, O.; BEKAS, P.; Bilge, G.; Boyaci, Y.O.; Brunelli, E.; Circosta, V.; Crocetta, F.; Durucan, F.; Erdem, M.; et al. New Mediterranean Biodiversity Records (December 2012). Mediterr. Mar. Sci. 2012, 13, 312–327. [Google Scholar] [CrossRef]
- Margaritis, M.; Tsikliras, A.C.; Dogrammatzi, A.; Nalmpanti, M.; Bourtzis, T.; Maniati, M.; Karachle, P.K. Data collected from questionnaires to fishers in relation to four non-indigenous species. In Deliverable 2.4 in 4ALIEN: Biology and the Potential Economic Exploitation of Four Alien Species in the Hellenic Seas; MIS 5049511: Athens, Greece, 2021. (In Greek) [Google Scholar]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Kulbicki, M.; Beets, J.; Chabanet, P.; Cure, K.; Darling, E.; Floeter, S.R.; Galzin, R.; Green, A.; Harmelin-Vivien, M.; Hixon, M.; et al. Distributions of Indo-Pacific lionfishes Pterois spp. in their native ranges: Implications for the Atlantic invasion. Mar. Ecol. Prog. Ser. 2012, 446, 189–205. [Google Scholar] [CrossRef]
- Vergés, A.; Tomas, F.; Cebrian, E.; Ballesteros, E.; Kizilkaya, Z.; Dendrinos, P.; Karamanlidis, A.A.; Spiegel, D.; Sala, E. Tropical rabbitfish and the deforestation of a warming temperate sea. J. Ecol. 2014, 102, 1518–1527. [Google Scholar] [CrossRef]
- Scardi, M.; Martin, C.S.; Valavanis, V.; Fraschetti, S.; Belluscio, A.; Gristina, M.; Salomidi, M.; Punzo, E.; Panayotidis, P.; Giannoulaki, M. Modeling of protected habitats using predictor variables. In Task 1.3 in Mediterranean Sensitive Habitats (MEDISEH); Final Report, DG MARE Specific Contract SI2.600741; Hellenic Centre for Marine Research: Heraklion, Greece, 2013. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Vignali, S.; Barras, A.G.; Arlettaz, R.; Braunisch, V. SDMtune: An R package to tune and evaluate species distribution models. Ecol. Evol. 2020, 10, 11488–11506. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Zenetos, A.; Belchior, C.; Cardoso, A.C. Invading European Seas: Assessing pathways of introduction of marine aliens. Ocean Coast. Manag. 2013, 76, 64–74. [Google Scholar] [CrossRef]
- Johnston, M.W.; Purkis, S.J. Are lionfish set for a Mediterranean invasion? Modelling explains why this is unlikely to occur. Mar. Pollut. Bull. 2014, 88, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Kimball, M.E.; Miller, J.M.; Whitfield, P.E.; Hare, J.A. Thermal tolerance and potential distribution of invasive lionfish (Pterois volitans/miles complex) on the east coast of the United States. Mar. Ecol. Prog. Ser. 2004, 283, 269–278. [Google Scholar] [CrossRef]
- Marras, S.; Cucco, A.; Antognarelli, F.; Azzurro, E.; Milazzo, M.; Bariche, M.; Butenschön, M.; Kay, S.; Di Bitetto, M.; Quattrocchi, G.; et al. Predicting future thermal habitat suitability of competing native and invasive fish species: From metabolic scope to oceanographic modelling. Conserv. Physiol. 2015, 3, cou059. [Google Scholar] [CrossRef] [PubMed]
- Hiddink, J.G.; Ben Rais Lasram, F.; Cantrill, J.; Davies, A.J. Keeping pace with climate change: What can we learn from the spread of Lessepsian migrants? Glob. Change Biol. 2012, 18, 2161–2172. [Google Scholar] [CrossRef]
- Gavriel, T.; Pickholtz, R.; Belmaker, J. Large individual-level variability in diel activity and depth use for the common Lionfish (Pterois miles). Front. Mar. Sci. 2021, 8, 790930. [Google Scholar] [CrossRef]
- Stergiou, K.I. Feeding habits of the Lessepsian migrant Siganus luridus in the eastern Mediterranean, its new environment. J. Fish Biol. 1988, 33, 531–543. [Google Scholar] [CrossRef]
- Ozvarol, Y.; Ertan, O.O.; Turna, I.I. The grazing effect of Siganus luridus Rüppell, 1828 on Posidonia oceanica (L.) Delile, 1813 meadows in Turkish Mediterranean coast (Gazipașa/Antalya). J. Food Agric. Environ. 2011, 9, 531–533. [Google Scholar]
- Rojas, D.C.; Monteagudo, P.C.; Schmitter-Soto, J.J.; Wong, R.I.C.; Torres, H.S.; Sansón, E.C.; Rodríguez, A.G.; Osorio, A.F.; Pantoja, L.E.; Guerra, D.C.; et al. Density, size, biomass, and diet of lionfish in Guanahacabibes National Park, western Cuba. Aquat. Biol. 2016, 24, 219–226. [Google Scholar] [CrossRef]
- Zannaki, K.; Corsini-Foka, M.; Kampouris, T.E.; Batjakas, I.E. First results on the diet of the invasive Pterois miles (Actinopterygii: Scorpaeniformes: Scorpaenidae) in the Hellenic waters. Acta Ichthyol. Et Piscat. 2019, 49, 311–317. [Google Scholar] [CrossRef]
- Dabruzzi, T.F.; Bennett, W.A.; Fangue, N.A. Thermal ecology of red lionfish Pterois volitans from southeast Sulawesi, Indonesia, with comparisons to other Scorpaenidae. Aquat. Biol. 2017, 26, 1–14. [Google Scholar] [CrossRef]
- Golani, D. Colonization of the Mediterranean by Red Sea fishes via the Suez Canal-Lessepsian migration. Fish Invasions Mediterr. Sea Change Renew. 2010, 145, 188. [Google Scholar]
- George, C.J.; Athanassiou, V. A two year study of the fishes appearing in the seine fishery of St George Bay, Lebanon. Ann. Del Mus. Civ. Di Stor. Nat. Di Genova 1967, 76, 237–294. [Google Scholar]
- Golani, D. Trophic adaptation of Red Sea fishes to the eastern Mediterranean environment—Review and new data. Isr. J. Zool. 1993, 39, 391–402. [Google Scholar]
- Galil, B.S. Alien species in the Mediterranean Sea—Which, when, where, why? Hydrobiologia 2008, 606, 105–116. [Google Scholar] [CrossRef]
- Koutsidi, M.; Moukas, C.; Tzanatos, E. Trait-based life strategies, ecological niches, and niche overlap in the nekton of the data-poor Mediterranean Sea. Ecol. Evol. 2020, 10, 7129–7144. [Google Scholar] [CrossRef]
- Azzurro, E.; Fanelli, E.; Mostarda, E.; Catra, M.; Andaloro, F. Resource partitioning among early colonizing Siganus luridus and native herbivorous fish in the Mediterranean: An integrated study based on gut-content analysis and stable isotope signatures. J. Mar. Biol. Assoc. United Kingd. 2007, 87, 991–998. [Google Scholar] [CrossRef]
- Bariche, M.; Letourneur, Y.; Harmelin-Vivien, M. Temporal fluctuations and settlement patterns of native and Lessepsian herbivorous fishes on the Lebanese coast (eastern Mediterranean). Environ. Biol. Fishes 2004, 70, 81–90. [Google Scholar] [CrossRef]
- Magneville, C.; Leréec Le Bricquir, M.L.; Dailianis, T.; Skouradakis, G.; Claverie, T.; Villéger, S. Long-duration remote underwater videos reveal that grazing by fishes is highly variable through time and dominated by non-indigenous species. Remote Sens. Ecol. Conserv. 2022. [Google Scholar] [CrossRef]
- Albouy, C.; Guilhaumon, F.; Leprieur, F.; Lasram, F.B.R.; Somot, S.; Aznar, R.; Velez, L.; Le Loc’h, F.; Mouillot, D. Projected climate change and the changing biogeography of coastal Mediterranean fishes. J. Biogeogr. 2013, 40, 534–547. [Google Scholar] [CrossRef]
- Vasilakopoulos, P.; Maravelias, C.D.; Tserpes, G. The alarming decline of Mediterranean fish stocks. Curr. Biol. 2014, 24, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Côté, I.M.; Akins, L.; Underwood, E.; Curtis-Quick, J.; Green, S.J. Setting the record straight on invasive lionfish control: Culling works. PeerJ PrePrints 2014, 2, e398v1. [Google Scholar]
- Ragkousis, M.; Zenetos, A.; Souissi, J.B.; Hoffman, R.; Ghanem, R.; Taşkın, E.; Muresan, M.; Kaprova, E.; Slynko, E.; Dağlı, E.; et al. Unpublished Mediterranean and Black Sea records of marine alien, cryptogenic, and neonative species. BioInvasions Rec. 2023; in press. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solanou, M.; Valavanis, V.D.; Karachle, P.K.; Giannoulaki, M. Looking at the Expansion of Three Demersal Lessepsian Fish Immigrants in the Greek Seas: What Can We Get from Spatial Distribution Modeling? Diversity 2023, 15, 776. https://doi.org/10.3390/d15060776
Solanou M, Valavanis VD, Karachle PK, Giannoulaki M. Looking at the Expansion of Three Demersal Lessepsian Fish Immigrants in the Greek Seas: What Can We Get from Spatial Distribution Modeling? Diversity. 2023; 15(6):776. https://doi.org/10.3390/d15060776
Chicago/Turabian StyleSolanou, Maria, Vasilis D. Valavanis, Paraskevi K. Karachle, and Marianna Giannoulaki. 2023. "Looking at the Expansion of Three Demersal Lessepsian Fish Immigrants in the Greek Seas: What Can We Get from Spatial Distribution Modeling?" Diversity 15, no. 6: 776. https://doi.org/10.3390/d15060776
APA StyleSolanou, M., Valavanis, V. D., Karachle, P. K., & Giannoulaki, M. (2023). Looking at the Expansion of Three Demersal Lessepsian Fish Immigrants in the Greek Seas: What Can We Get from Spatial Distribution Modeling? Diversity, 15(6), 776. https://doi.org/10.3390/d15060776