
The Acoustic Repertoire of Bottlenose Dolphins (Tursiops truncatus) in the Cres-Lošinj Archipelago (Croatia): Site Dependent Diel and Seasonal Changes





Abstract
:1. Introduction
2. Materials and Methods
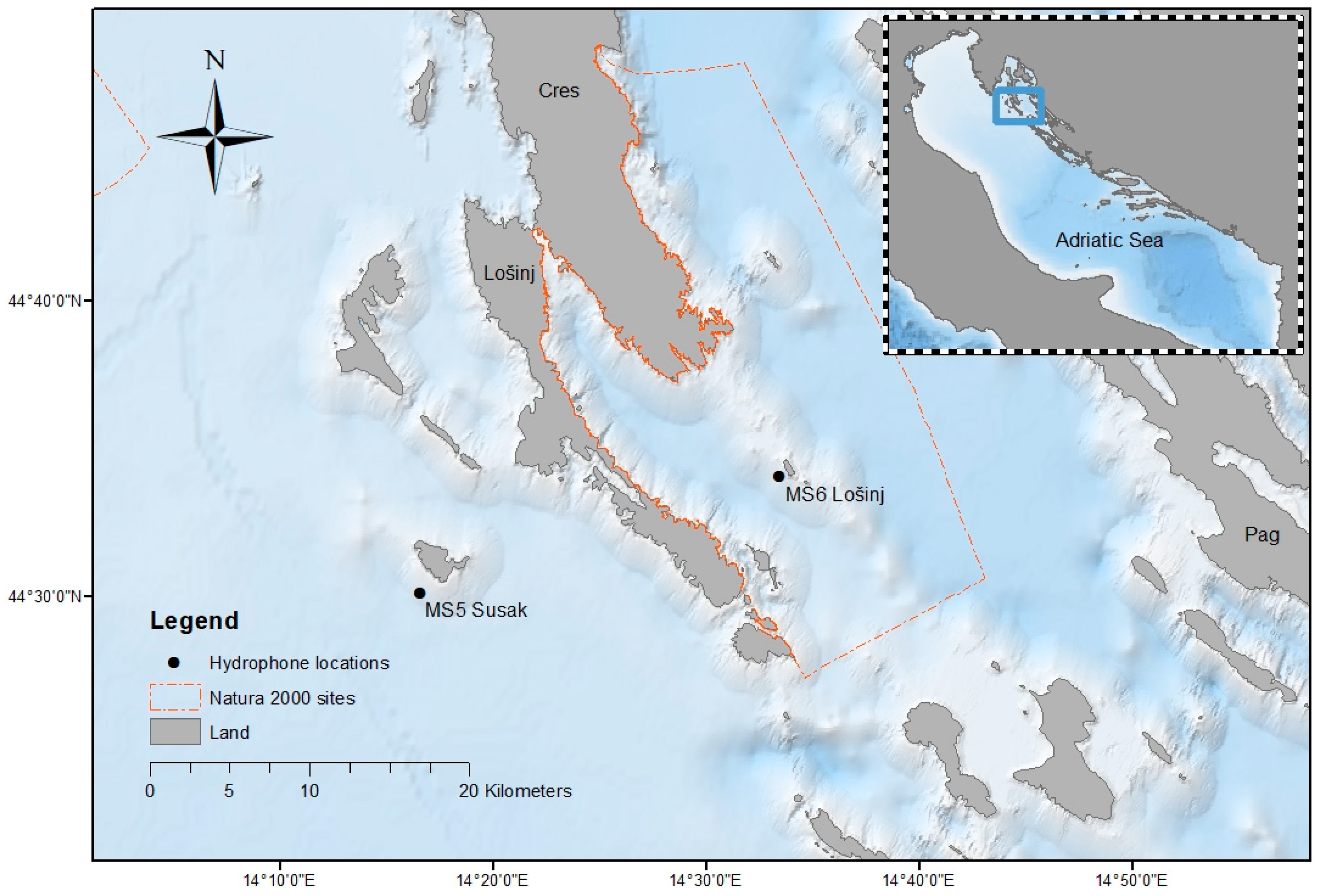
2.1. Study Area
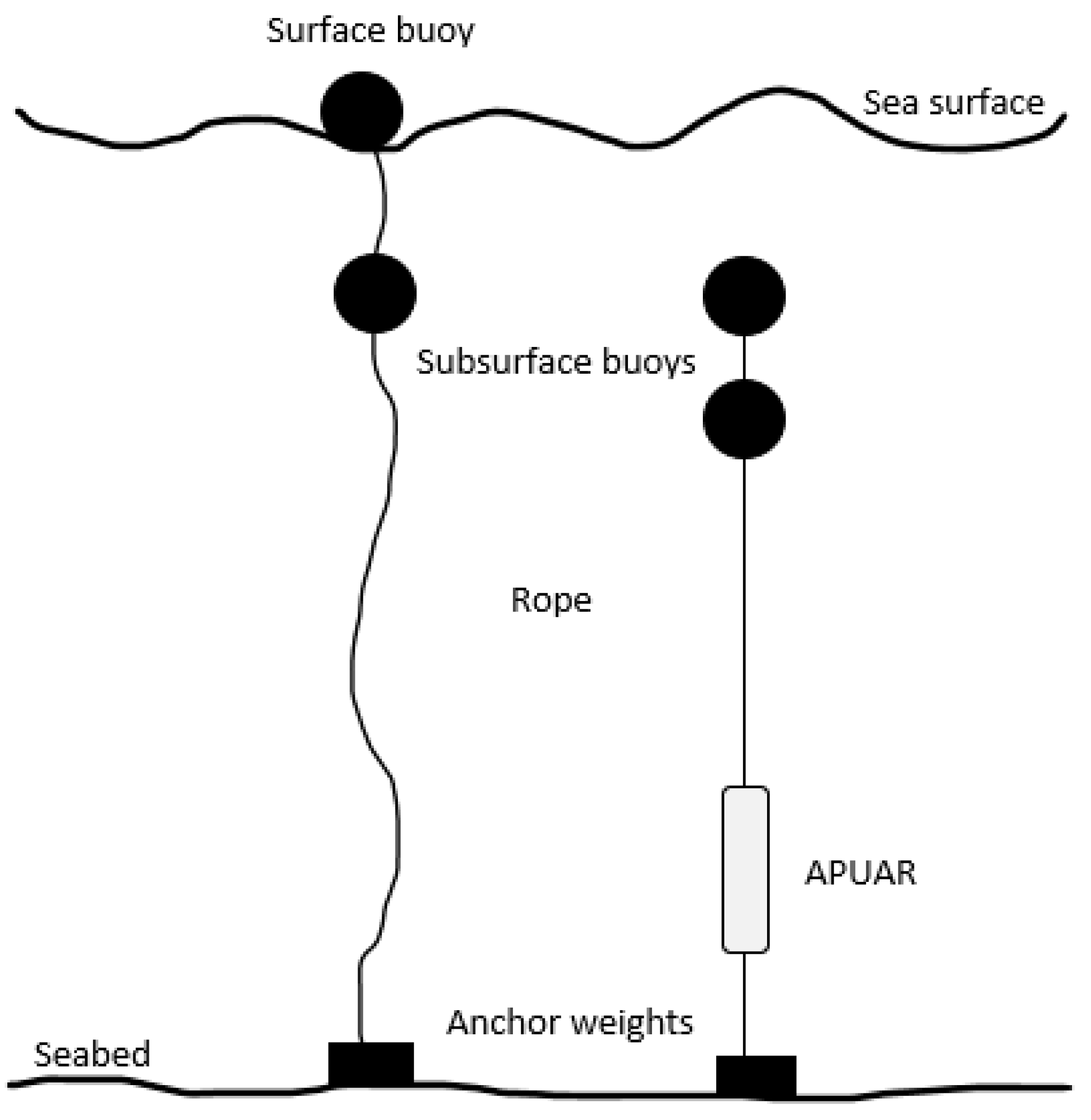
2.2. Data Collection
2.3. Data Analysis
2.3.1. Identifying Bottlenose Dolphin Vocalizations
- i.
- ii.
- iii.
- iv.
- v.
- Echolocation clicks were short and intense broadband sounds with ultrasonic peak frequencies that are usually produced in rapid succession [14].
- vi.
2.3.2. Characterizing Bottlenose Dolphin Vocalization Parameters
2.3.3. Sound Pressure Level Analysis
2.3.4. Statistical Analysis
3. Results
3.1. Distribution and Characterization of Bottlenose Dolphin Vocalizations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MS5 Susak | MS6 Lošinj | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Seasonal Period | Diel Period | n of Hours Analyzed | Whistle | Chirp | LFN | BP | Whistle | Chirp | LFN | BP |
| NTS | Day | 104 | 237 (2.3) | 91 (0.8) | 45 (0.4) | 13 (0.1) | 55 (0.5) | 9 (0.08) | 64 (0.6) | 2 (0.01) |
| NTS | Night | 88 | 13 (0.1) | 13 (0.1) | 12 (0.1) | 4 (0.04) | 207 (2.3) | 12 (0.1) | 83 (0.9) | 0 (0.0) |
| TS | Day | 208 | 27 (0.1) | 20 (0.09) | 173 (0.8) | 2 (0.01) | 72 (0.3) | 18 (0.08) | 200 (0.9) | 5 (0.02) |
| TS | Night | 104 | 8 (0.08) | 16 (0.1) | 233 (2.24) | 3 (0.02) | 46 (0.4) | 1 (0.01) | 10 (0.09) | 1 (0.01) |
3.2. Diel and Seasonal Differences in Bottlenose Dolphin Vocalization Parameters
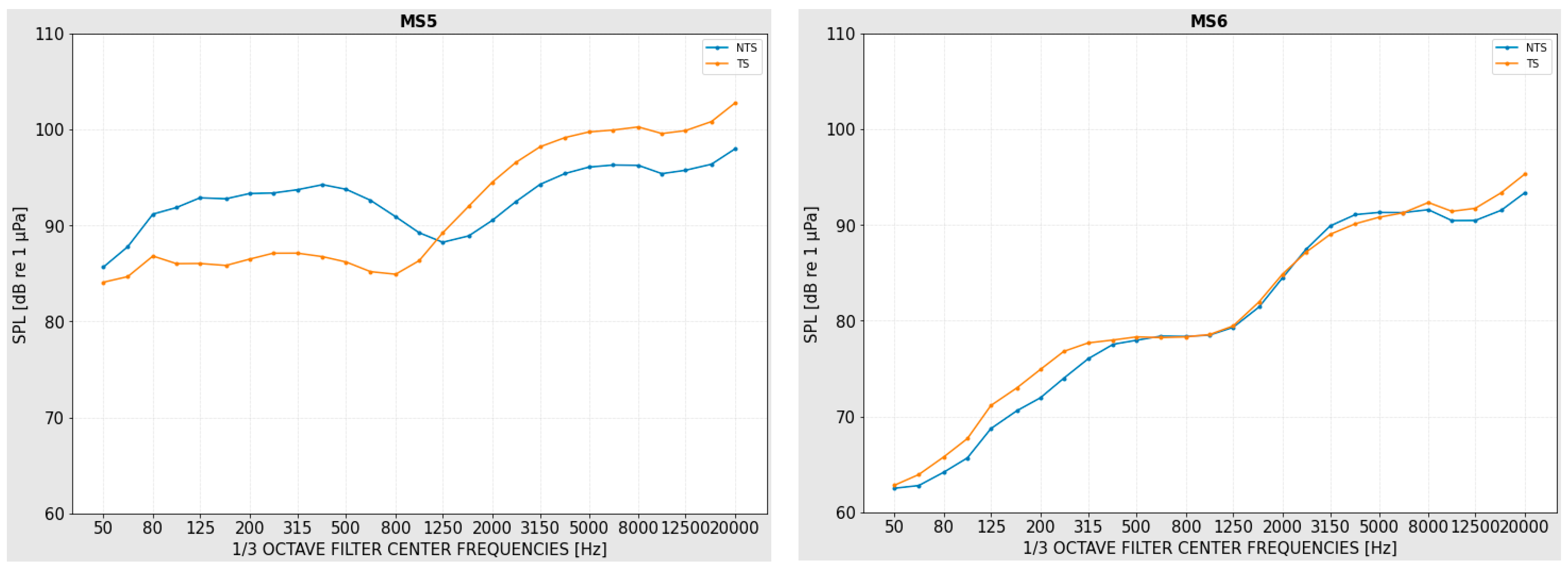
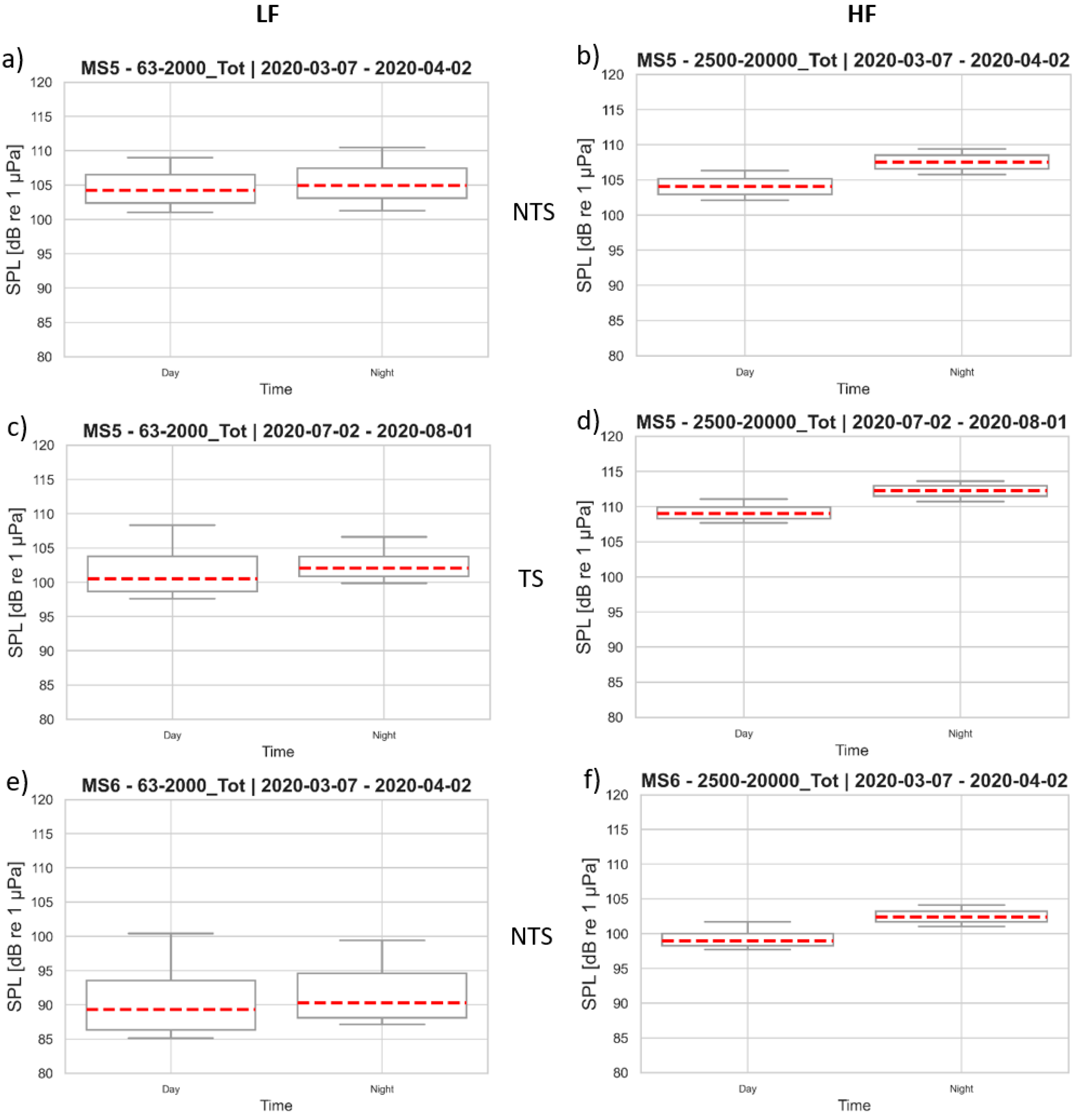
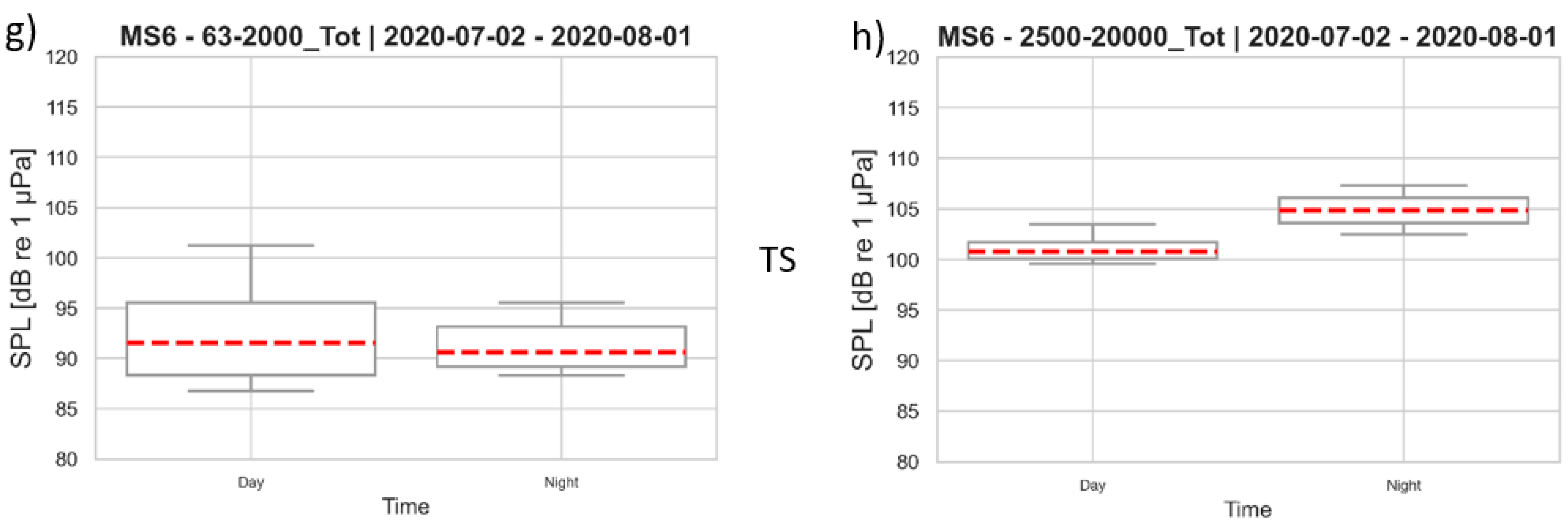
3.3. Sound Pressure Levels at the Monitoring Stations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bearzi, G.; Fortuna, C.M.; Reeves, R.R. Ecology and conservation of common bottlenose dolphins Tursiops truncates in the Mediterranean Sea. Mammal Rev. 2008, 39, 92–123. [Google Scholar] [CrossRef]
- Bearzi, G.; Holcer, D.; di Sciara, G.N. The role of historical dolphin takes and habitat degradation in shaping the present status of northern Adriatic cetaceans. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, 363–379. [Google Scholar] [CrossRef]
- Genov, T.; Kotnjek, P.; Lesjak, J.; Hace, A.; Fortuna, C.M. Bottlenose dolphins (Tursiops truncatus) in Slovenian and adjacent waters (northern Adriatic Sea). Ann. Ser. Hist. Nat. 2008, 18, 227–244. [Google Scholar]
- Gaspari, S.; Dooley, C.; Shreves, K.; Silva, C.S.; Chapman, N.; Genov, T.; Gonzalvo, J.; Holcer, D.; Moura, A.E. Connectivity patterns of bottlenose dolphins (Tursiops truncatus) in the north-east Mediterranean: Implications for local conservation. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Bearzi, G.; Politi, E.; di Sciara, G.N. Diurnal behavior of free-ranging bottlenose dolphins in the Kvarnerić (northern Adriatic Sea). Mar. Mammal Sci. 1999, 15, 1065–1097. [Google Scholar] [CrossRef]
- Pleslić, G.; Gospić, N.R.; Mackelworth, P.; Wiemann, A.; Holcer, D.; Fortuna, C. The abundance of common bottlenose dolphins (Tursiops truncatus) in the former special marine reserve of the Cres-Lošinj Archipelago, Croatia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 25, 125–137. [Google Scholar] [CrossRef]
- Bearzi, G.; di Sciara, G.N.; Politi, E. Social ecology of bottlenose dolphins in the Kvarnerić (northern Adriatic Sea). Mar. Mammal Sci. 1997, 13, 650–668. [Google Scholar] [CrossRef]
- Hermannsen, L.; Mikkelsen, L.; Tougaard, J.; Beedholm, K.; Johnson, M.; Madsen, P.T. Recreational vessels without Automatic Identification System (AIS) dominate anthropogenic noise contributions to a shallow water soundscape. Sci. Rep. 2019, 9, 15477. [Google Scholar] [CrossRef] [Green Version]
- Wilson, L.; Pine, M.K.; Radford, C.A. Small recreational boats: A ubiquitous source of sound pollution in shallow coastal habitats. Mar. Pollut. Bull. 2022, 174, 113295. [Google Scholar] [CrossRef]
- Bearzi, G.; Bonizzoni, S.; Riley, M.A.; Santostasi, N.L. Bottlenose dolphins in the north-western Adriatic Sea: Abundance and management implications. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 651–664. [Google Scholar] [CrossRef]
- Pleslić, G.; Rako-Gospić, N.; Holcer, D. Bottlenose dolphins (Tursiops truncatus) in North Dalmatia, Croatia: Occurrence and demographic parameters. Mar. Mammal Sci. 2021, 37, 142–161. [Google Scholar] [CrossRef]
- Picciulin, M.; Armelloni, E.; Falkner, R.; Rako-Gospić, N.; Radulović, M.; Pleslić, G.; Muslim, S.; Mihanović, H.; Gaggero, T. Characterization of the underwater noise produced by recreational and small fishing boats (<14 m) in the shallow-water of the Cres-Lošinj Natura 2000 SCI. Mar. Pollut. Bull. 2022, 183, 114050. [Google Scholar] [CrossRef] [PubMed]
- Janik, V.M.; Sayigh, L.S. Communication in bottlenose dolphins: 50 years of signature whistle research. J. Comp. Physiol. A 2013, 199, 479–489. [Google Scholar] [CrossRef]
- Au, W.W.L. The Sonar of Dolphins; Springer: New York, NY, USA, 1993. [Google Scholar]
- Allen, M.C.; Read, A.J. Habitat selection of foraging bottlenose dolphins in relation to boat density near clearwater, Florida. Mar. Mammal Sci. 2000, 16, 815–824. [Google Scholar] [CrossRef]
- Bejder, L.; Samuels, A.; Whitehead, H.; Gales, N.; Mann, J.; Connor, R.; Heithaus, M.; Watson-Capps, J.; Flaherty, C.; Krützen, M. Decline in relative abundance of bottlenose dolphins exposed to long-term disturbance. Conserv. Biol. 2006, 20, 1791–1798. [Google Scholar] [CrossRef] [Green Version]
- van Ginkel, C.; Becker, D.M.; Gowans, S.; Simard, P. Whistling in a noisy ocean: Bottlenose dolphins adjust whistle frequencies in response to real-time ambient noise levels. Bioacoustics 2017, 27, 391–405. [Google Scholar] [CrossRef]
- Fouda, L.; Wingfield, J.E.; Fandel, A.D.; Garrod, A.; Hodge, K.B.; Rice, A.N.; Bailey, H. Dolphins simplify their vocal calls in response to increased ambient noise. Biol. Lett. 2018, 14, 20180484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckstaff, K.C. Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mammal Sci. 2004, 20, 709–725. [Google Scholar] [CrossRef]
- May-Collado, L.J.; Wartzok, D. A comparison of bottlenose dolphin whistles in the Atlantic ocean: Factors promoting whistle variation. J. Mammal. 2008, 89, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Rako, N.; Fortuna, C.M.; Holcer, D.; Mackelworth, P.; Nimak-Wood, M.; Pleslić, G.; Sebastianutto, L.; Vilibić, I.; Wiemann, A.; Picciulin, M. Leisure boating noise as a trigger for the displacement of the bottlenose dolphins of the Cres-Lošinj archipelago (northern Adriatic Sea, Croatia). Mar. Pollut. Bull. 2013, 68, 77–84. [Google Scholar] [CrossRef]
- Gospić, N.R.; Picciulin, M. Changes in whistle structure of resident bottlenose dolphins in relation to underwater noise and boat traffic. Mar. Pollut. Bull. 2016, 105, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Rako, N.; Picciulin, M.; Vilibić, I.; Fortuna, C.M. Spatial and temporal variability of sea ambient noise as an anthropogenic pressure index: The case of the Cres-Lošinj archipelago, Croatia. J. Mar. Biol. Assoc. U. K. 2013, 93, 27–36. [Google Scholar] [CrossRef]
- Petrizzo, A.; Barbanti, A.; Barfucci, G.; Bastianini, M.; Biagiotti, I.; Bosi, S.; Centurelli, M.; Chavanne, R.; Codarin, A.; Costantini, I.; et al. First assessment of underwater sound levels in the Northern Adriatic Sea at the basin scale. Sci. Data 2023, 10, 137. [Google Scholar] [CrossRef]
- Picciulin, M.; Bolgan, M.; Rako-Gospić, N.; Petrizzo, A.; Radulović, M.; Falkner, R. A Fish and Dolphin Biophony in the Boat Noise-Dominated Soundscape of the Cres-Lošinj Archipelago (Croatia). J. Mar. Sci. Eng. 2022, 10, 300. [Google Scholar] [CrossRef]
- Herzing, D.L. Acoustics and Social Behavior of Wild Dolphins: Implications for a Sound Society. In Hearing by Whales and Dolphins; Springer: New York, NY, USA, 2000; pp. 225–272. [Google Scholar] [CrossRef]
- Boisseau, O. Quantifying the acoustic repertoire of a population: The vocalizations of free-ranging bottlenose dolphins in Fiordland, New Zealand. J. Acoust. Soc. Am. 2005, 117, 2318–2329. [Google Scholar] [CrossRef] [PubMed]
- López, B.D.; Shirai, J.A.B. Mediterranean Common Bottlenose Dolphin’s Repertoire and Communication Use. In Dolphins: Anatomy, Behavior, and Threats; Nova Science Publishers: New York, NY, USA, 2009; pp. 129–148. [Google Scholar]
- Gridley, T.; Nastasi, A.; Kriesell, H.; Elwen, S. The acoustic repertoire of wild common bottlenose dolphins (Tursiops truncatus) in Walvis Bay, Namibia. Bioacoustics 2015, 24, 153–174. [Google Scholar] [CrossRef] [Green Version]
- Wahlberg, M.; Jensen, F.H.; Soto, N.A.; Beedholm, K.; Bejder, L.; Oliveira, C.; Rasmussen, M.; Simon, M.; Villadsgaard, A.; Madsen, P.T. Source parameters of echolocation clicks from wild bottlenose dolphins (Tursiops aduncus and Tursiops truncatus). J. Acoust. Soc. Am. 2011, 130, 2263–2274. [Google Scholar] [CrossRef] [Green Version]
- La Manna, G.; Rako-Gospić, N.; Sarà, G.; Gatti, F.; Bonizzoni, S.; Ceccherelli, G. Whistle variation in Mediterranean common bottlenose dolphin: The role of geographical, anthropogenic, social, and behavioral factors. Ecol. Evol. 2020, 10, 1971–1987. [Google Scholar] [CrossRef]
- Pace, D.S.; Tumino, C.; Silvestri, M.; Giacomini, G.; Pedrazzi, G.; Pavan, G.; Papale, E.; Ceraulo, M.; Buscaino, G.; Ardizzone, G. Bray-Call Sequences in the Mediterranean Common Bottlenose Dolphin (Tursiops truncatus) Acoustic Repertoire. Biology 2022, 11, 367. [Google Scholar] [CrossRef]
- Rako-Gospić, N.; Radulović, M.; Vučur, T.; Pleslić, G.; Holcer, D.; Mackelworth, P. Factor associated variations in the home range of a resident Adriatic common bottlenose dolphin population. Mar. Pollut. Bull. 2017, 124, 234–244. [Google Scholar] [CrossRef]
- Gridley, T.; Berggren, P.; Cockcroft, V.G.; Janik, V.M. Whistle vocalizations of Indo-Pacific bottlenose dolphins (Tursiops aduncus) inhabiting the south-west Indian Ocean. J. Acoust. Soc. Am. 2012, 132, 4032–4040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilly, J.C.; Miller, A.M. Sounds emitted by the bottlenose dolphin: The audible emissions of captive dolphins under water or in air are remarkably complex and varied. Science 1961, 133, 1689–1693. [Google Scholar] [CrossRef]
- Janik, V.M.; King, S.L.; Sayigh, L.S.; Wells, R.S. Identifying signature whistles from recordings of groups of unrestrained bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 2013, 29, 109–122. [Google Scholar] [CrossRef]
- Griffin, D.R. Echoes of Bats and Men; Double-Day and, Co.: Garden City, NY, USA, 1959. [Google Scholar]
- Schultz, K.W.; Cato, D.H.; Corkeron, P.J.; Bryden, M.M. Low frequency narrow-band sounds produced by bottlenose dolphins. Mar. Mammal Sci. 1995, 11, 503–509. [Google Scholar] [CrossRef]
- Watkins, W.A. The Harmonic Interval: Fact or Artifact in Spectral Analysis of Pulse Trains. In Marine Bio-Acoustics; Tavolga, W.N., Ed.; Pergamon Press: Oxford, UK, 1967; pp. 15–42. [Google Scholar]
- Lammers, M.O.; Au, W.W.L.; Aubauer, R.; Nachtigall, P.E. A Comparative Analysis of the Pulsed Emissions of Free-Ranging Hawaiian Spinner Dolphins (Stenella longirostris). In Echolocation in Bats and Dolphins; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 414–419. [Google Scholar]
- Janik, V.M. Food-related bray calls in wild bottlenose dolphins (Tursiops truncatus). Proc. R. Soc. B Boil. Sci. 2000, 267, 923–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascão, I.; Lammers, M.O.; Prieto, R.; Santos, R.S.; Silva, M.A. Temporal patterns in acoustic presence and foraging activity of oceanic dolphins at seamounts in the Azores. Sci. Rep. 2020, 10, 3610. [Google Scholar] [CrossRef] [Green Version]
- Heiler, J.; Elwen, S.; Kriesell, H.; Gridley, T. Changes in bottlenose dolphin whistle parameters related to vessel presence, surface behaviour and group composition. Anim. Behav. 2016, 117, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Betke, K.; Folegot, T.; Matuschek, R.; Pajala, J.; Persson, L.; Tegowski, J.; Tougaard, J.; Wahlberg, M. BIAS Standards for Signal Processing. Aims, Processes and Recommendations, Amended version; Verfus, U.K., Sigray, P., Eds.; BIAS, 2015; Available online: https://biasproject.files.wordpress.com/2016/01/bias_sigproc_standards_v5_final.pdf (accessed on 15 November 2021).
- Jones, G.J.; Sayigh, L.S. Geographic variation in rates of vocal production of free-ranging bottlenose dolphins. Mar. Mammal Sci. 2002, 18, 374–393. [Google Scholar] [CrossRef]
- Quick, N.J.; Janik, V.M. Whistle rates of wild bottlenose dolphins (Tursiops truncatus): Influences of group size and behavior. J. Comp. Psychol. 2008, 122, 305–311. [Google Scholar] [CrossRef]
- Janik, V.M.; Sayigh, L.S.; Wells, R.S. Signature whistle shape conveys identity information to bottlenose dolphins. Proc. Natl. Acad. Sci. USA 2006, 103, 8293–8297. [Google Scholar] [CrossRef] [Green Version]
- King, S.L.; Janik, V.M. Bottlenose dolphins can use learned vocal labels to address each other. Proc. Natl. Acad. Sci. USA 2013, 110, 13216–13221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, M.C.; Caldwell, D.K.; Tyack, P.L. Review of the Signature-Whistle Hypothesis for the Atlantic Bottlenose Dolphin. In The Bottlenose Dolphin; Leatherwood, S., Reeves, R.R., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 199–234. [Google Scholar]
- Simard, P.; Lace, N.; Gowans, S.; Quintana-Rizzo, E.; Kuczaj, S.A.; Wells, R.S.; Mann, D.A. Low frequency narrow-band calls in bottlenose dolphins (Tursiops truncatus): Signal properties, function, and conservation implications. J. Acoust. Soc. Am. 2011, 130, 3068–3076. [Google Scholar] [CrossRef] [PubMed]
- Lammers, M.O.; Au, W.W.L.; Herzing, D.L. The broadband social acoustic signaling behavior of spinner and spotted dolphins. J. Acoust. Soc. Am. 2003, 114, 1629–1639. [Google Scholar] [CrossRef] [Green Version]
- Janik, V.M. Acoustic Communication in Delphinids. Adv. Study Behav. 2009, 40, 123–157. [Google Scholar] [CrossRef]
- Falkner, R.; Pokupec, D.; Granziol, L.; Frleta-Valić, M.; Rako-Gospić, N.; Vučur Blazinić, T.; Constaratas, A.; Radulović, M.; Cukrov Car, M.; Hofs, J. Report on the Distribution of Target Species. SOUNDSCAPE Project, WP4. 2021; p. 49. Available online: https://www.italy-croatia.eu/web/soundscape (accessed on 19 March 2023).
- Guerra, M.; Dawson, S.M.; Brough, T.E.; Rayment, W.J. Effects of boats on the surface and acoustic behaviour of an endangered population of bottlenose dolphins. Endanger. Species Res. 2014, 24, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Kastelein, R.; Vaughan, N.; Walton, S.; Wiepkema, P. Food intake and body measurements of Atlantic bottlenose dolphins (Tursiops truncates) in captivity. Mar. Environ. Res. 2002, 53, 199–218. [Google Scholar] [CrossRef]
- Luís, A.R.; May-Collado, L.J.; Rako-Gospić, N.; Gridley, T.; Papale, E.; Azevedo, A.; Silva, M.A.; Buscaino, G.; Herzing, D.; dos Santos, M.E. Vocal universals and geographic variations in the acoustic repertoire of the common bottlenose dolphin. Sci. Rep. 2021, 11, 11847. [Google Scholar] [CrossRef]





| Statistics | Start Freq (kHz) | End Freq (kHz) | Min Freq (kHz) | Max Freq (kHz) | Peak Freq (kHz) | Freq Range (kHz) | Duration (s) | IP (n) | Harmonics (n) | Pulse Rep. Rate (p/s) |
|---|---|---|---|---|---|---|---|---|---|---|
| Whistles | ||||||||||
| n | 665 | 665 | 665 | 665 | 665 | 665 | 665 | 665 | - | - |
| Mean ± SD | 7.05 2.56 | 9.10 3.99 | 6.03 1.92 | 10.99 4.04 | 7.73 2.86 | 4.97 3.18 | 0.79 0.43 | 1.01 1.35 | - - | - - |
| Min | 0.87 | 3.65 | 0.87 | 4.59 | 1.13 | 0.29 | 0.14 | 0.00 | - | - |
| Max | 18.17 | 22.46 | 14.84 | 22.46 | 17.34 | 15.09 | 6.10 | 10.00 | - | - |
| CV | 36.31 | 43.84 | 31.83 | 36.77 | 36.95 | 64.05 | 55.01 | 134.11 | - | - |
| Chirps | ||||||||||
| n | 180 | 180 | 180 | 180 | 180 | 180 | 180 | - | - | - |
| Mean ± SD | 1.93 0.90 | 2.40 0.77 | 1.78 0.81 | 2.51 0.78 | 2.11 0.79 | 0.73 3.35 | 0.07 0.02 | - - | - - | - - |
| Min Max | 0.26 8.75 | 0.51 8.14 | 0.51 8.14 | 1.35 9.02 | 1.13 8.72 | 0.25 1.91 | 0.02 0.10 | - - | - - | - - |
| CV | 46.50 | 31.95 | 45.42 | 31.12 | 37.24 | 48.04 | 29.51 | - | - | - |
| LFN sounds | ||||||||||
| n | - | - | 820 | 820 | 820 | 820 | 820 | - | 820 | - |
| Mean ± SD | - - | - - | 0.33 0.10 | 0.70 0.14 | 0.50 0.10 | 0.37 0.12 | 0.04 0.02 | - - | 0.07 0.29 | - - |
| Min Max | - - | - - | 0.11 0.72 | 0.41 1.58 | 0.28 1.03 | 0.18 1.22 | 0.01 0.19 | - - | 0.00 4.00 | - - |
| CV | - | - | 29.70 | 20.06 | 20.90 | 33.60 | 61.18 | - | 402.69 | - |
| BP sounds | ||||||||||
| n | - | - | - | - | - | - | 30 | - | - | 30 |
| Mean ± SD | - - | - - | - - | - - | - - | - - | 0.71 0.19 | - - | - - | 358.87 104.51 |
| Min Max | - - | - - | - - | - - | - - | - - | 0.03 0.69 | - - | - - | 102.88 571.07 |
| CV | - | - | - | - | - | - | 107.54 | - | - | 29.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falkner, R.; Picciulin, M.; Pleslić, G.; Rako-Gospić, N. The Acoustic Repertoire of Bottlenose Dolphins (Tursiops truncatus) in the Cres-Lošinj Archipelago (Croatia): Site Dependent Diel and Seasonal Changes. Diversity 2023, 15, 787. https://doi.org/10.3390/d15060787
Falkner R, Picciulin M, Pleslić G, Rako-Gospić N. The Acoustic Repertoire of Bottlenose Dolphins (Tursiops truncatus) in the Cres-Lošinj Archipelago (Croatia): Site Dependent Diel and Seasonal Changes. Diversity. 2023; 15(6):787. https://doi.org/10.3390/d15060787
Chicago/Turabian StyleFalkner, Raffaela, Marta Picciulin, Grgur Pleslić, and Nikolina Rako-Gospić. 2023. "The Acoustic Repertoire of Bottlenose Dolphins (Tursiops truncatus) in the Cres-Lošinj Archipelago (Croatia): Site Dependent Diel and Seasonal Changes" Diversity 15, no. 6: 787. https://doi.org/10.3390/d15060787
APA StyleFalkner, R., Picciulin, M., Pleslić, G., & Rako-Gospić, N. (2023). The Acoustic Repertoire of Bottlenose Dolphins (Tursiops truncatus) in the Cres-Lošinj Archipelago (Croatia): Site Dependent Diel and Seasonal Changes. Diversity, 15(6), 787. https://doi.org/10.3390/d15060787