Abstract
In order to investigate the effects of various nitrogen (ammonium) concentrations on the epibiotic bacterial community associated with Oocystis borgei, a metabarcoding sequencing method was employed. The 16S rDNA sequencing and bioinformatic analysis were conducted on cultures of O. borgei that were grown on four different nitrogen element concentrations (2.5, 10, 50, and 100 mg/L), and the differences in the epibiotic bacterial community and functions of O. borgei among different nitrogen concentrations were compared. The results showed that the chlorophyll a content of O. borgei increased with increasing nitrogen concentrations. A total of 43 operational taxonomic units (OTUs) were obtained from the four groups, which were categorized into 7 phyla, 9 classes, 18 orders, 21 families, and 26 genera. Winogradskyella was the dominant genus in the groups with 2.5 and 100 mg/L nitrogen, while Marinobacter and Winogradskyella were relatively abundant in the groups with 50 and 100 mg/L nitrogen. Functional analysis using PICRUST2 showed that the three most abundant gene functions were carbohydrate metabolism, amino acid transport and metabolism, ribosomal structure and biogenesis, and energy production and conversion. BugBase phenotypic analysis revealed that there were no significant differences in phenotypes between the groups with 2.5 and 10 mg/L nitrogen, while the group with 50 mg/L nitrogen exhibited higher abundance in aerobicity, biofilm formation, mobile genetic elements, and stress tolerance phenotypes. Most of the bacteria in this work belonged to the aerobic types. A redundancy analysis (RDA) of environmental factors demonstrated that nitrogen concentration showed a positive correlation with species changes in the groups with 10 and 50 mg/L nitrogen; chlorophyll a exhibited a positive correlation with species changes in the groups with 50 and 100 mg/L nitrogen. Nitrogen concentration significantly influenced the epibiotic bacterial community associated with O. borgei, leading to changes in dominant species and community structure. This study provides important references for understanding the functional characteristics of the epiphytic microbial community of O. borgei and the exploration of specific microorganisms.
1. Introduction
The area surrounding algae is known as the microenvironment of algal interplay, also referred to as the phycosphere [1]. Florez [2] defines the phycosphere as a holistic biofilm, elucidating the symbiotic interactions between algae and associated bacteria as epiphytic interactions. The diversity of epiphytic bacteria directly determines the functionality of algal interplay communities. The interactions between algae and bacteria can influence and shape microbial diversity, primarily through physical contact, substrate exchange, information exchange, and horizontal gene transfer [3]. Bacteria provide essential growth factors, vitamins, iron carriers, and surfactants that inhibit pathogenic agents on the surface of algae [4]. Aerobic bacteria consume oxygen, converting large organic compounds into small molecules such as ammonium and nitrite that microalgae can utilize, while also supplying carbon dioxide and other growth-promoting molecules [5,6]. Meanwhile, microalgae continuously secrete metabolites, including peptides, and lipids which provide carbon, oxygen, nutrients, and energy for aerobic heterotrophic bacteria. The interactions between algae and bacteria are mutually selective and evolutionary [7]. The microenvironment formed by different algae fosters specific epiphytic bacteria [8], exhibiting host-specific characteristics. In other words, the microbial community structure in the phycosphere of the same algae species from different environments is more similar than the structure between the phycospheres of different seaweed species in the same environment. For instance, the similarity in microbial community structures within the phycosphere of Codium fragile in different environments is as high as 70% [9]. Lachnit et al. [10] analyzed the composition of attached bacterial communities on brown alga (Fucus vesiculosus), red alga (Gracilaria vermiculophylla), and green alga (Ulva intestinalis) in Kiel Fjord, and found that the surfaces of marine macroalgae possess species-specific and temporally adaptive biofilms which play a crucial role in algal survival. The synergistic relationships between algae and bacteria have also been applied in the popular use of algae such as Chlorella and Scenedesmus for organic wastewater treatment, resulting in significant improvements in nitrogen and phosphorus removal rates [11,12]. Moreover, the biofilm, a complex matrix composed of numerous bacteria and algae, is capable of modifying the pH through algal secretions and respiration, promoting the formation of different epiphytic bacterial communities, and stimulating heterotrophic microbial activities. This, in turn, influences community functionality. The algae–bacteria interactions have had significant importance for the balance of aquatic ecosystems and biogeochemical cycles [13]. Algae and bacteria have coexisted in the ocean for over two hundred million years. As we know, the oceans represent the largest active carbon reservoir, with micro-sized planktonic organisms playing a crucial role in the oceanic carbon sink process. It is precisely this mutually beneficial relationship between algae and bacteria that promotes the continuous regeneration and transformation of dissolved organic carbon (DOC) [14]. Studying the diversity of bacteria attached to algae can enhance the predictability of biomass production, especially under limiting conditions such as nitrogen availability in summer, as well as defense responses against bacterial pathogens and epiphytes [15]. Furthermore, certain attached bacteria are closely associated with the morphogenesis of algae. Under sterile conditions, Monostroma oxyspermum, for example, fails to develop into a normal gametophyte [16]. While there have been numerous studies on the diversity of attached bacteria and specific algal–bacterial relationships in large seaweeds, research on the specific diversity of attached bacteria in microalgae under various environmental conditions remains limited.
Oocystis borgei, a member of the phylum Chlorophyta and order Chlorococcales, is widely distributed in organic-rich water bodies such as estuaries and aquaculture ponds. It becomes a dominant microalga during and after aquaculture due to its preference for such environments. O. borgei thrives at optimal concentrations of NO3-N and PO4-P, which are 11.32 mg·L−1 and 1.29 mg·L−1, respectively. It exhibits characteristics such as prolonged community persistence and stable population growth, effectively reducing nitrogen pollution [17], increasing dissolved oxygen, inhibiting the growth of Vibrio and harmful blue-green algae [18], and demonstrating good adsorption efficiency for heavy metals like Cu2+ and Zn2+ [19]. Therefore, O. borgei is widely utilized in aquaculture for regulating water quality [20]. Xie [21] found that the quantity and variety of metabolites produced by O. borgei were significantly influenced by different nitrogen concentrations. Liu et al. [22] found that O. borgei exhibits high absorption efficiency for ammonium nitrogen. Building a benign microalgal community mainly composed of O. borgei in water pools can effectively remove ammonia nitrogen and nitrite nitrogen and improve water quality. Yang et al. [23] revealed that under nitrogen limitations and lack of light, O. borgei altered its reproductive mode, resulting in insufficient cellular material synthesis, primarily expanding the population through the production of two or three autospores.
It is necessary to maintain the balance of algae and bacteria in a water environment to regulate water quality by using O. borgei in shrimp culture. This organism can team up with Rhodopseudomonas palustris to establish a synergistic algae–bacteria consortia that is adept at taking up various nitrogen compounds, thereby enhancing water purification [24]. It has been indicated that the dominance of O. borgei in microalgae communities can contribute to enhancing water quality and managing the proliferation of bacteria associated with L. vannamei diseases [25]. As nitrogen is a pivotal nutrient for both microalgae and bacteria, fluctuations in its levels can sway their proliferation and, in turn, impact the equilibrium of the O. borgei-associated bacterial community. Investigating how different nitrogen levels affect these microbial populations is of paramount importance for managing water quality and preventing disease in shrimp culture. The mutualistic exchanges between algae and bacteria are garnering interest; they are engaged in a dynamic exchange of metabolites that bolster bacterial population growth, while the activity of advantageous bacteria can, in turn, promote microalgal growth [26]. If the number of beneficial bacteria in the epiphytic bacteria of O. borgei can be controlled by adjusting the concentration of nitrogen, thus inhibiting the growth of pathogenic bacteria, it will be more feasible to construct an algae–bacteria system to maintain the stability of the pond ecosystem.
This study aims to investigate the changes in the attached bacterial community of O. borgei under different nitrogen concentrations using metabarcoding sequencing. By analyzing and predicting the functions of the attached bacterial community, we aim to explore the role of the attached bacterial community in the nitrogen adaptation process of O. borgei and uncover the relationship between specific bacterial groups and O. borgei. These findings will provide valuable references for establishing an algal–bacterial system using O. borgei in aquaculture production and serve as an important reference for further research into the functional elucidation and exploration of specific microorganisms in the interfacial microbial communities associated with O. borgei.
2. Materials and Methods
2.1. Culture and Maintenance
The culture of O. borgei used in this study was isolated from an aquaculture pond and maintained in the Laboratory of Algal Development and Aquaculture Environmental Ecological Restoration, Guangdong Ocean University. The O. borgei culture was maintained in artificial seawater (with a salinity of 31) enriched with f/2-Si medium (the nitrogen source was replaced with ammonium) at 25 °C, and 24 h light at approximately 50 μmol quanta m−2·s−1 supplied by white fluorescent lights. The culture was transferred every other two weeks to ensure that the cells were in the exponential growth stage.
2.2. Experiment Design
Cells of O. borgei at exponential growth stage were harvested, and washed several times using sterilized artificial seawater. Subsequently, the cells were inoculated into the f/2 modified medium supplemented with NH4Cl, resulting in nitrogen element mass concentrations of 2.5, 10, 50, and 100 mg L−1, respectively (groups were labeled as N2.5, N10, N50, and N100, respectively). The initial volume was 400 mL, with a cell density of 1.5 × 106 cells/L. The cultures were maintained at a temperature of 25 °C, under a 24 h light cycle with an illumination intensity of 50 μmol quanta m−2·s−1. The culture flasks were shaken four times per day, and each experimental condition was performed in triplicate for a duration of seven days.
2.3. Chlorophyll a Content Measurement
An aliquot of 5 mL algal culture of each treatment was centrifuged at 5000 rpm for 10 min. The supernatant was discarded, and 95% ethanol was added to a final volume of 5 mL. After mixing, the samples were stored in darkness for 24 h. Then, the samples were centrifuged again, and the absorbance values of the supernatant were measured at 630 nm, 647 nm, 664 nm, and 750 nm using a UV spectrophotometer (UV-2450, Shimadzu, Japan). The calculation formula for chlorophyll a content (mg/L) was as follows [27]:
where A630, A647, and A664 represent the absorbance values at wavelengths of 630 nm, 647 nm, and 664 nm, respectively. V1 is the volume (mL) after the extraction solution has been made up to the final volume, and V2 is the volume (mL) of the algal solution.
2.4. Sample Processing and Sequencing of Microbial Diversity
According to the manufacturer, the total DNA of the algal–bacteria mixed samples was extracted with a soil DNA extraction kit (Biomarker Soil Genomic DNA Kit, Beijing, China). Then, specific primers with barcodes were synthesized using the 16S full-length primers 27F and 1492R [28]. PCR amplification (95 °C for 5 min; 30 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 90 s, with a final extension of 72 °C for 7 min) was performed using these primers, and the resulting products were purified, quantified, and normalized to create a library. The qualified library was then sequenced using PacBio Sequel. The sequencing was carried out by Biomarker Biotechnology Co., Ltd., Beijing, China.
2.5. Data Analyses
After obtaining the raw data, Circular Consensus Sequencing (CCS) sequences were exported and the Effective-CCS sequences were obtained through the following steps: Firstly, using lima v1.7.0 software, CCS sequences are identified by their barcodes to obtain the Raw-CCS sequence data. Then, the primer sequences are recognized by cutadapt 1.9.1 software, and data quality is evaluated by analyzing parameters such as sequence count and length. The data are filtered by removing primer sequences and applying length filtering, resulting in Clean-CCS sequences without primer sequences. Furthermore, chimeric sequences are identified and removed, resulting in the Effective-CCS sequences produced by UCHIME v4.2 software. After filtering, Usearch software v5.64 [29] is used to cluster reads at a 97.0% similarity level, obtain operational taxonomic units (OTUs), and perform species annotation and taxonomic analysis using the NCBI reference database and a naïve Bayesian classifier combined with alignment-based methods. In addition, QIIME1 software is employed to assess alpha and beta diversity indices and compare the similarity of species diversity between different samples. Finally, functional gene composition is inferred using the Integrated Microbial Genomes (IMG) database to analyze functional differences between different samples or groups. Redundancy analysis (RDA) is a linear model-based ranking method developed based on correspondence analysis, which is mainly used to reflect the relationship between bacteria communities or samples and environmental factors.
The Shannon and Simpson indices are used in alpha diversity analysis to measure species diversity, taking into account the abundance and evenness of species in a sample community. In cases where species have the same abundance, a greater evenness among the species within a community is associated with higher diversity. Higher values of the Shannon and Simpson indices indicate greater species diversity in the sample. The calculation formulas are as follows [30]:
The calculation of the Simpson index (DSimpon) and Shannon index (H):
Here, Sobs represents the number of observed OTUs; ni is the number of OTUs containing i sequences; and N is the number of all sequences.
The calculation of the ACE index (Sace):
Among them, , , , and ni represents the number of OUTs containing i sequences; Srare represents the number of OTUs containing “abund” sequences; Sabund represents the number of OTUs with more than “abund” sequences; and “abund” represents the threshold of “advantage” OTU, which defaults to 10.
The computation of the Chao1 index (Schao1) is as follows:
where Schao1 represents the estimated OTU number; Sobs represents the observed OTU number; n1 represents the OTU number with only one number; and n2 represents the OUT number with only two sequences.
The calculation of the goods coverage (C) rate is as follows:
where C represents the coverage rate, F1 is the number of species with a sample size of only 1, and N is the total number of individuals in the sample.
3. Results
3.1. The Changes in Chlorophyll a Concentration of O. borgei under Different Nitrogen Concentrations
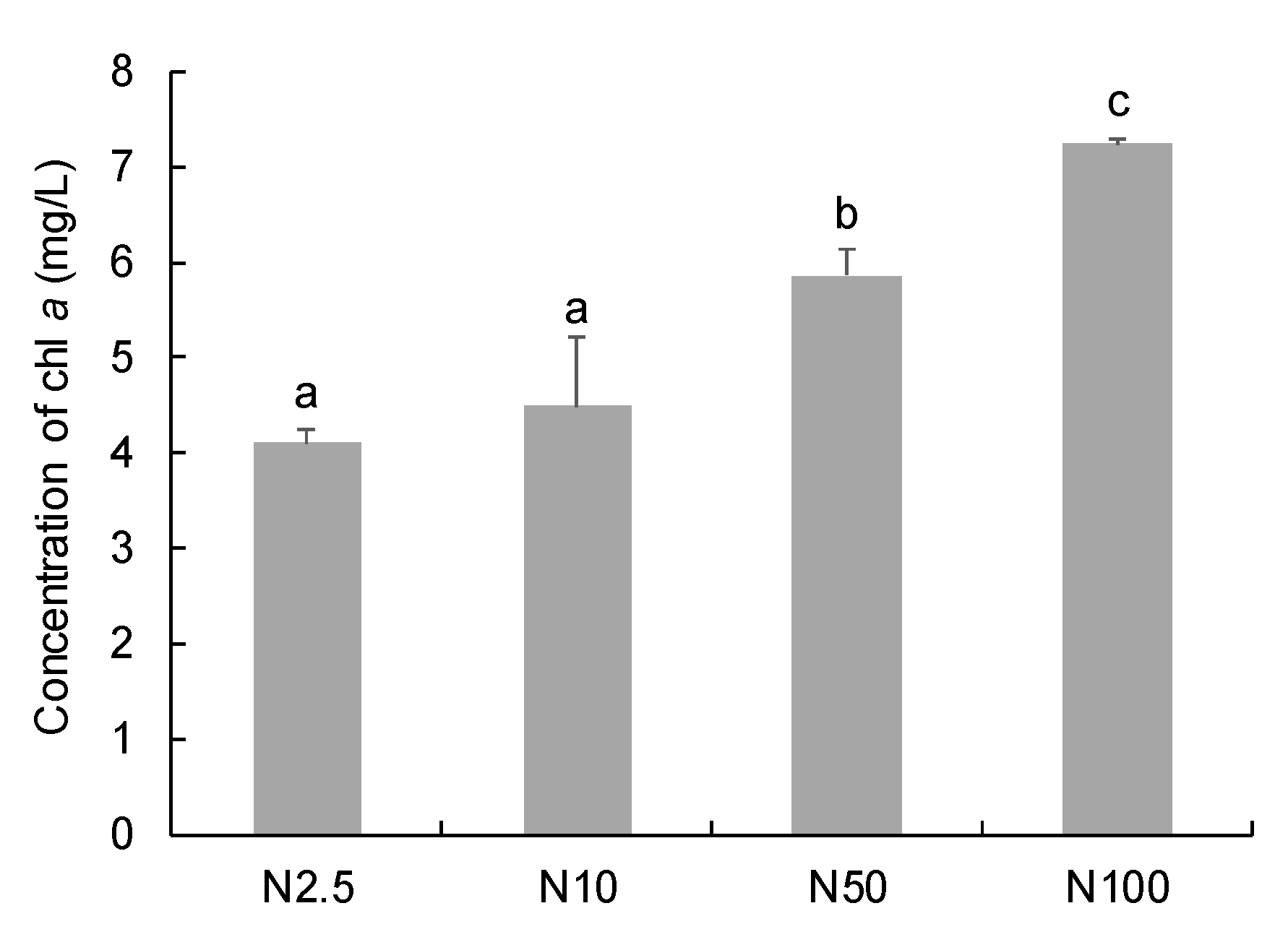
Nitrogen concentrations (2.5–100 mg/L) had a positive effect on the chlorophyll a content of O. borgei (Figure 1). The highest Chl a concentration (7.23 mg/L) was observed in the group with 100 mg/L nitrogen, and was significantly higher than that in other groups with 2.5–50 mg/L nitrogen (p < 0.05). The lowest Chl a concentration (4.10 mg/L) was observed in the group with 2.5 mg/L nitrogen. There was no significant difference in chlorophyll a concentration between the groups with 2.5 mg/L and 10 mg/L nitrogen (p > 0.05).
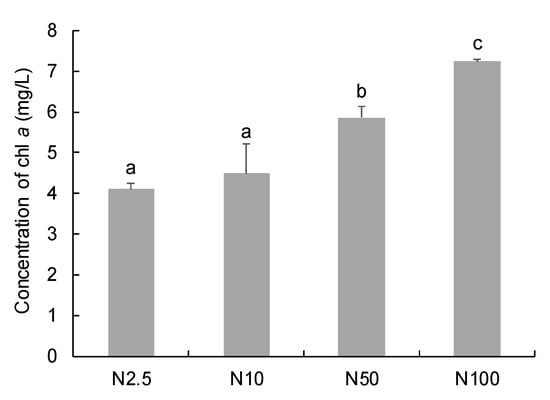
Figure 1.
Chlorophyll a concentration (mg/L) of O. borgei under different nitrogen concentrations. The bars with different letters are statistically different (p < 0.05).
3.2. Changes in Nitrogen Concentrations during the Experiments
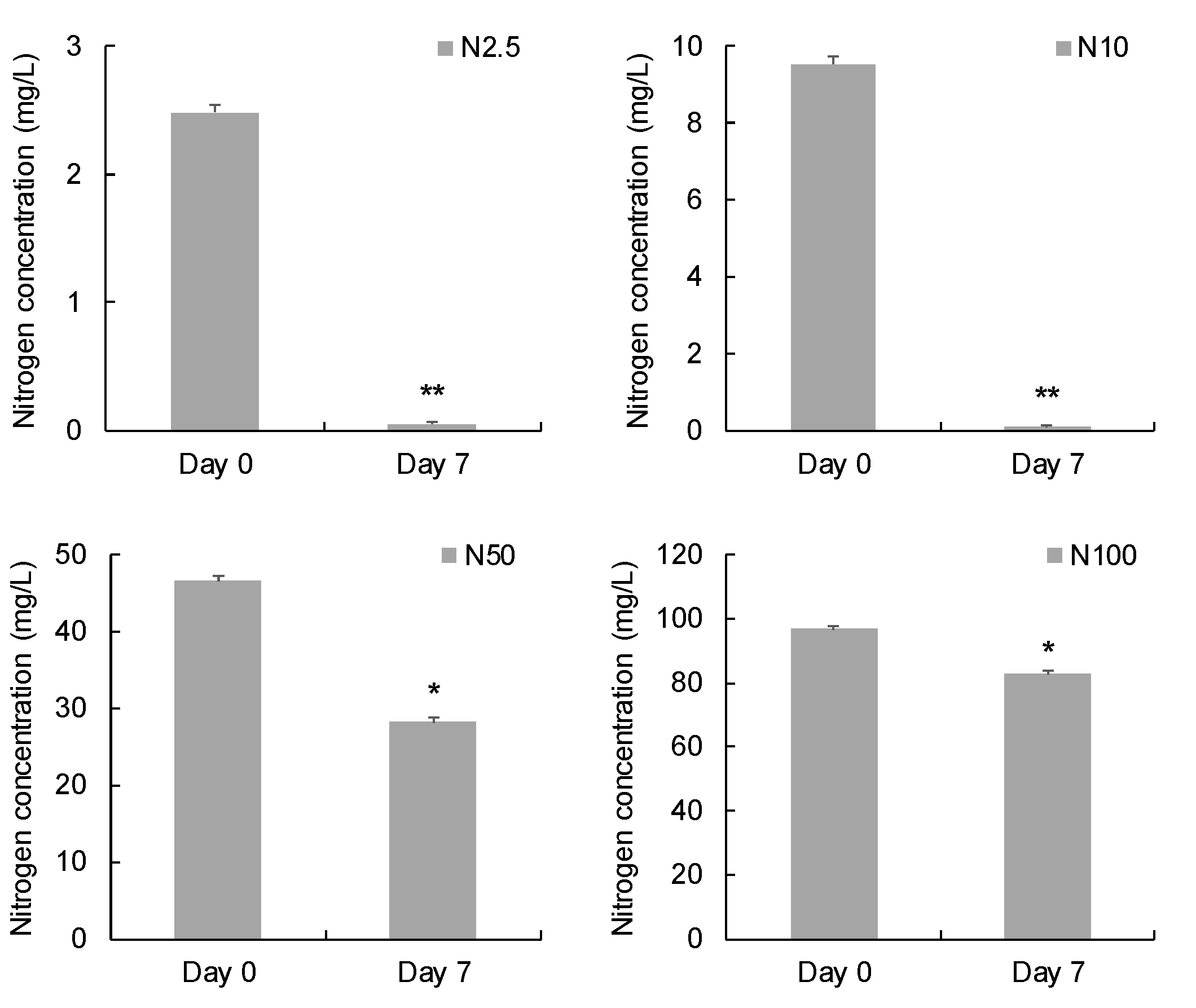
After 7 days’ cultivation, 2.44 mg/L, 9.43 mg/L, 18.47 mg/L, and 13.93 mg/L nitrogen were removed from the groups supplied with 2.5, 10, 50, and 100 mg/L nitrogen, respectively (Figure 2). The nitrogen removal efficiencies in the groups supplied with 2.5 and 10 mg/L nitrogen were significantly higher than the groups supplied with 50 and 100 mg/L nitrogen (p < 0.05).
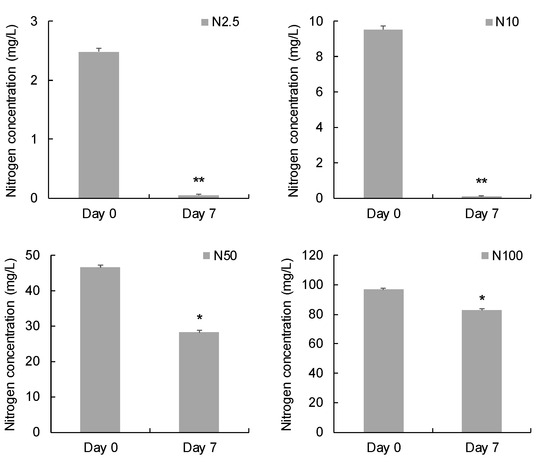
Figure 2.
Comparison of nitrogen concentration before and after 7 days’ incubation. (* indicates significant difference, p < 0.05; ** indicates extremely significant difference, p < 0.01.).
3.3. OTU and Beta Diversity Analyses of the Epiphytic Microbial Community of O. borgei
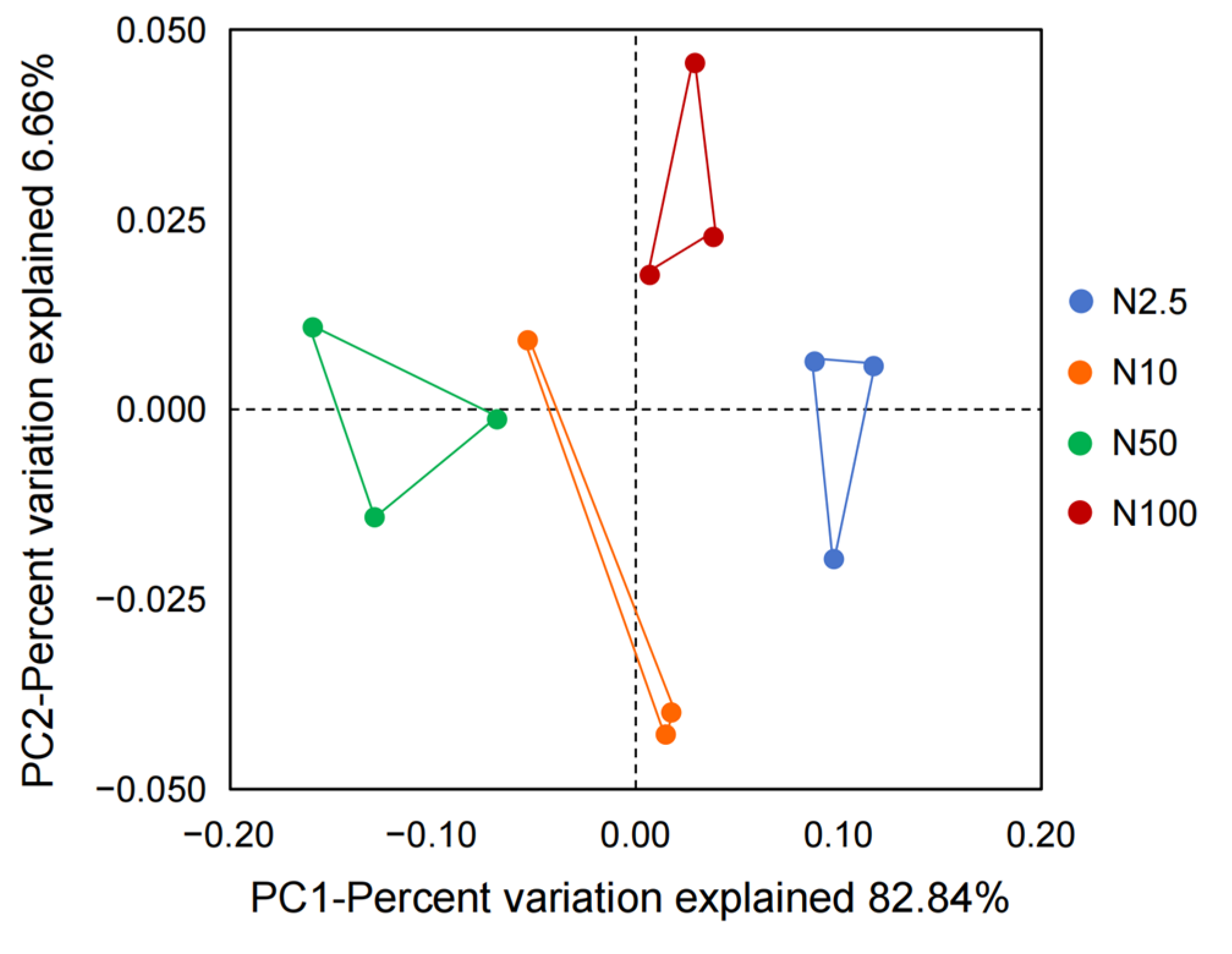
A total of 90,335 CCS sequences from 12 samples were generated through 16S rRNA gene metabarcoding sequencing, with from 7366 to 7654 CCS sequences in each sample, with an average of 7528 CCS sequences. A total of 43 OTUs were annotated at a 97.0% similarity level. Based on the division of OTUs, the similarity and dissimilarity of the epiphytic microbial community structure of O. borgei supplied with different nitrogen concentrations were compared. The principal coordinate analysis (PCoA) revealed that the groups supplied with 2.5, 10, 50, and 100 mg/L nitrogen were distributed in four different positions: right, bottom, left, and top, respectively (Figure 3). Those groups that were grouped together on the plot indicated a greater similarity in species diversity. The sample points from each group on the plot appeared to cluster together. The first principal component explained 82.84% of the variability, suggesting significant differences in the composition of the epiphytic microbial community of O. borgei under different nitrogen concentrations. Obviously, different nitrogen concentrations had a considerable impact on the composition of the epiphytic microbial community.
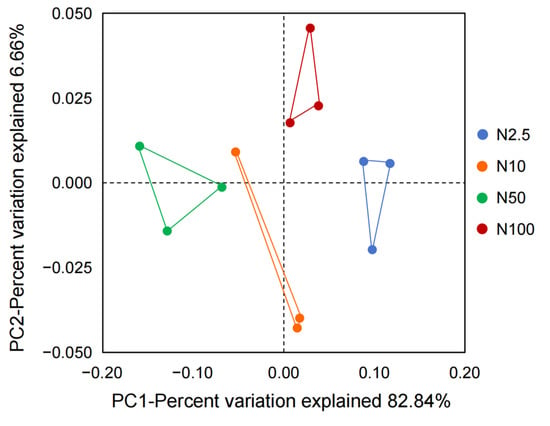
Figure 3.
PCoA plot of the microbial community structure in cultures of O. borgei supplied with different concentrations of nitrogen.
3.4. Analysis of the Species Composition of the Epiphytic Microbial Community of O. borgei
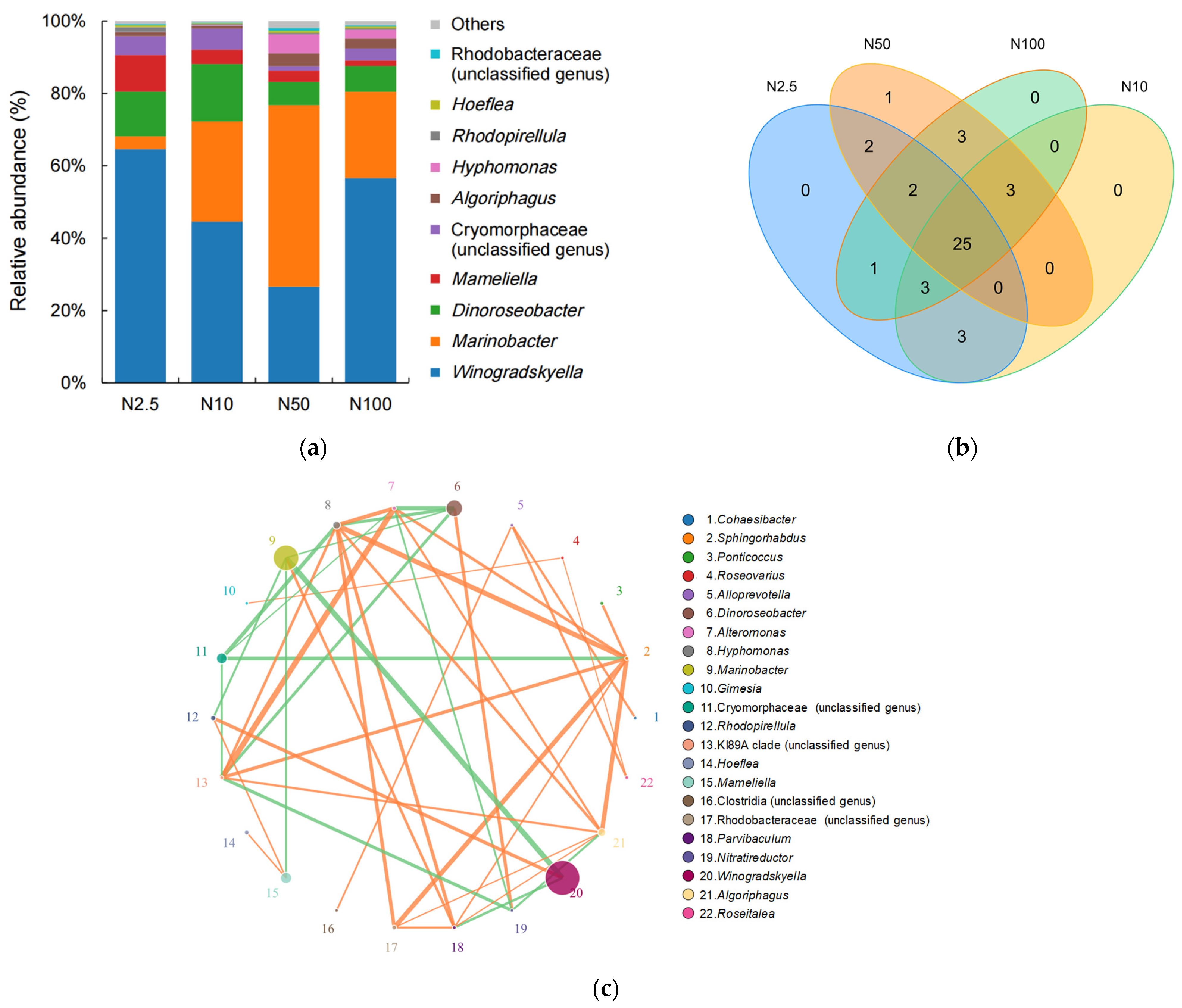
A total of 26 genera belonging to 21 families, 18 orders, 9 classes, and 7 phyla were annotated in the four groups supplied with 2.5, 10, 50, and 100 mg/L nitrogen (Figure 4). The four genera with the highest relative abundance in the epiphytic microbial community of O. borgei were Winogradskyella, Marinobacter, Dinoroseobacter, and Mameliella (Figure 4). The abundances of Dinoroseobacter were relatively higher in the lower nitrogen concentration groups (2.5 and 10 mg/L; Figure 4a). The abundances of Marinobacter were highest in the group supplied with 50 mg/L nitrogen, accounting for 50% of the total community, but abundance was much lower in the groups supplied with 2.5, 10, and 100 mg/L nitrogen (Figure 4a). Winogradskyella showed relatively higher abundance in the groups with 2.5 and 100 mg/L nitrogen (Figure 4a). The four groups shared a total of 25 genera, indicating a high degree of similarity in the composition of the epiphytic microbial community associated with O. borgei at different nitrogen concentrations (Figure 4b). The most abundant genera, Vibrio and Wingradskaia, exhibited the strongest correlation, and following them were the genera Roseobacter and Marinosulfonomonas, which were negatively correlated (Figure 4c). However, there was no correlation observed among Wingradskaia and Roseobacter or Marinosulfonomonas.
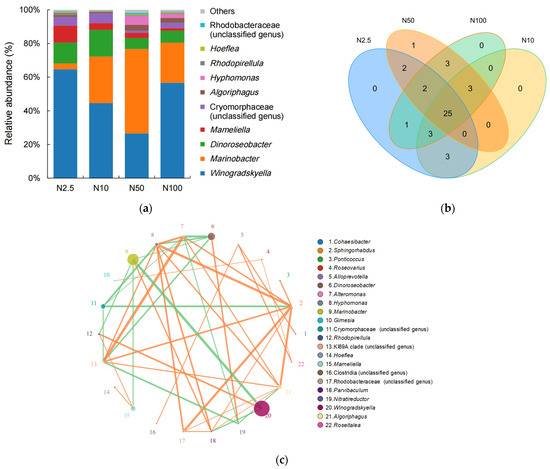
Figure 4.
(a) Stacked bar chart of the genus-level epiphytic microbial community associated with O. borgei. (b) Genus-level Venn diagram of the epiphytic microbial community associated with O. borgei. (c) A network diagram of the genus-level epiphytic microbial community associated with O. borgei. The lines in the diagram represent the correlations between different species, with the thickness of the lines indicating the strength of the correlation. The color of the lines represent the nature of the correlation, with orange lines indicating positive correlations and green lines indicating negative correlations.
3.5. Analysis of Species Diversity in the Epiphytic Microbial Community of O. borgei
There were no significant differences in the Shannon index and Simpson index among the four groups supplied with different nitrogen concentrations (Table 1). Furthermore, the Chao1 and Ace indices indicated that the species richness was ranked from high to low as follows: N10, N2.5, N50, and N100 groups. These differences were not significant, suggesting that the nitrogen concentration did not have a significant impact on the species diversity and species richness of O. borgei.

Table 1.
The alpha diversity index of the epiphytic bacterial community in O. borgei.
3.6. Functional Analysis and Prediction of the Epiphytic Bacterial Community in O. borgei
Using PICRUSt2 functional prediction analysis of the epiphytic microbial community of O. borgei, the differences and variations in functional genes involved in metabolic pathways among different groups were observed (Table 2). Combining the results from the comparison with the KEGG database, the top 20 pathway functions, as refined, were presented in Table 2. The top 10 pathways included carbohydrate metabolism, amino acid metabolism, energy metabolism, the metabolism of cofactors and vitamins, membrane transport, nucleotide metabolism, translation, signal transduction, replication and repair, and lipid metabolism (Table 2). There were significant variations in cellular motility, which might be the abundance of nitrogen resources advantageous for cellular activities. Moreover, there were no significant differences in the relative abundance of the same biological pathways among the groups (p > 0.05), indicating that nitrogen concentration did not have a significant impact on the metabolic function of the epiphytic microbial community of O. borgei.
Table 2.
The relative abundance values of KEGG metabolic pathway functions in the bacterial community associated with O. borgei.
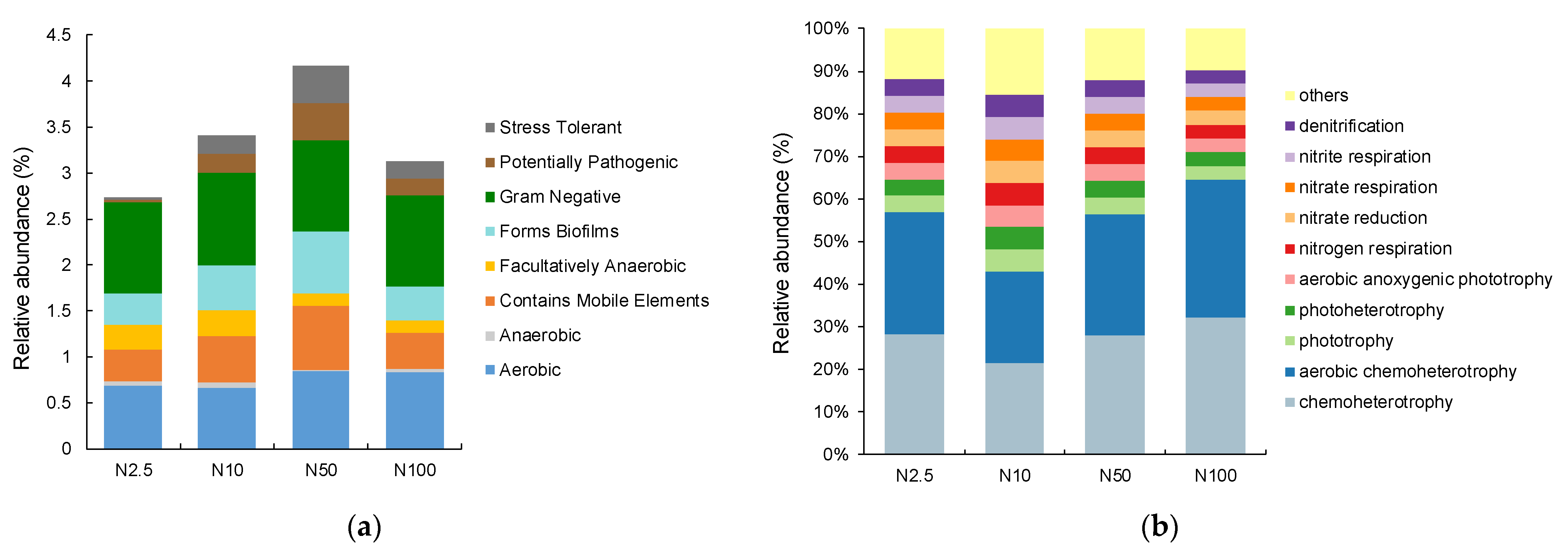
Bugbase phenotype analyses are commonly used to analyze the phenotypes of microbial community samples, including oxidative stress tolerance, pathogenic potential, Gram-positivity, Gram-negativity, biofilm formation, and mobile genetic elements. All identified bacteria belonged to the Gram-negative category, with the majority being aerobic (Figure 5a). There were no significant differences in the phenotypes of the epiphytic bacteria among the groups, while in the group with 50 mg/L nitrogen, the bacterial stress tolerance and potential pathogenicity were prominent.
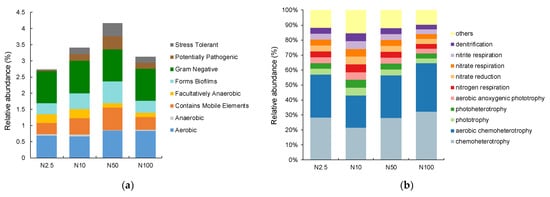
Figure 5.
(a) The BugBase phenotype analysis of bacterial communities associated with O. borgei with different nitrogen concentrations. (b) The FAPROTAX functional predictions of bacterial communities associated with O. borgei with different nitrogen concentrations.
The FAPROTAX database is a commonly used functional annotation database for analyzing microbial diversity, particularly in the context of marine or lake biochemical processes. Combining the FAPROTAX database with the functional annotation of the epiphytic bacterial community in O. borgei, the relative abundance of nitrogen element metabolic pathways was determined. The three highest relative abundances were chemoheterotrophy, aerobic chemoheterotrophy, and phototrophy (Figure 5b). The relative abundances of chemoheterotrophy and aerobic chemoheterotrophy in the epiphytic bacterial community of the groups with 50 and 100 mg/L nitrogen were higher than those in the groups with 2.5 and 10 mg/L nitrogen. The relative abundances of photoautotrophy and oxygenic photoautotrophy in the epiphytic bacterial community of the groups with 2.5 and 10 mg/L nitrogen were higher than those in the groups with 50 and 100 mg/L nitrogen.
3.7. Analyses of the Correlation between Bacterial Communities Associated with O. borgei and Environmental Factors
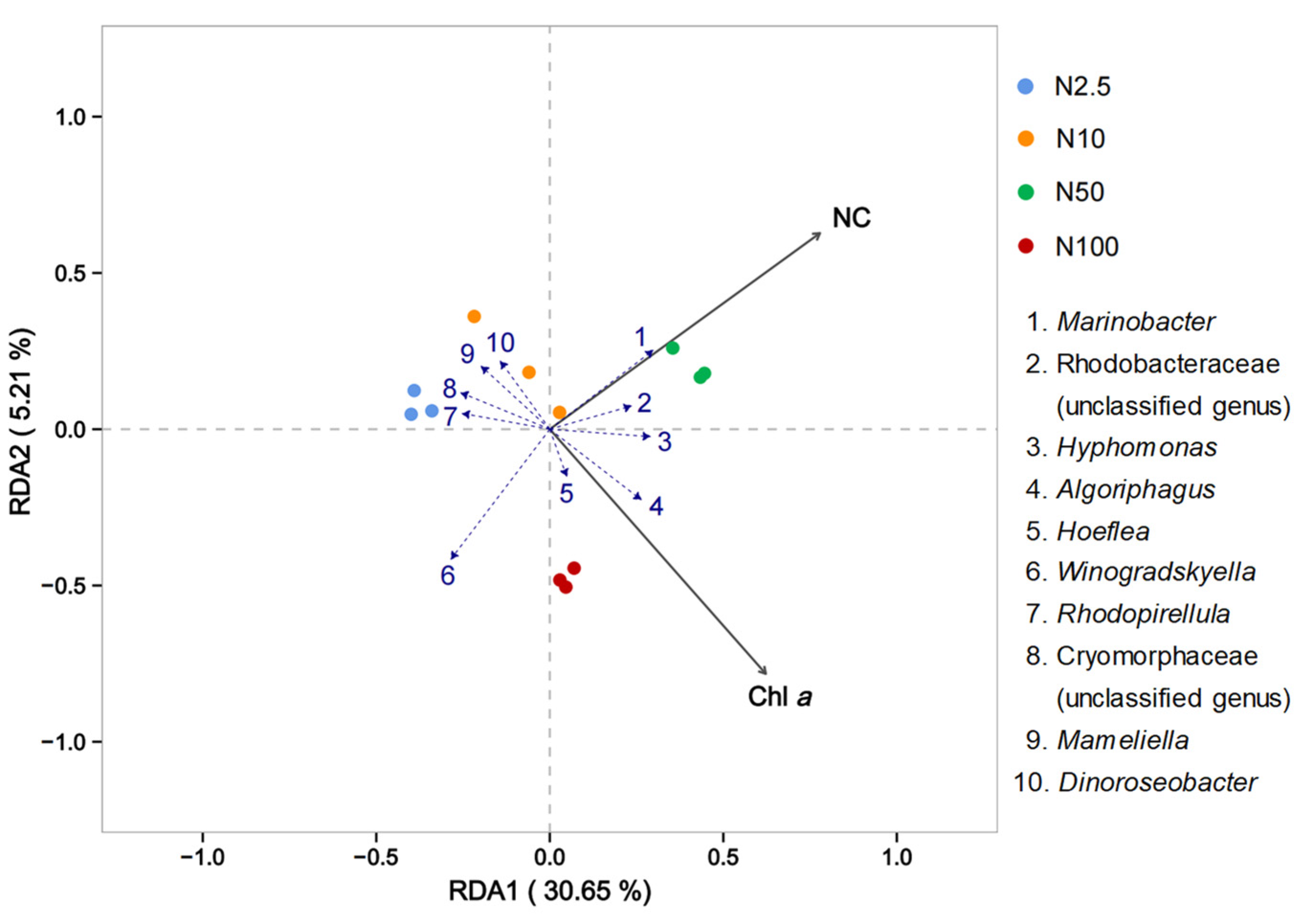
The RDA analysis of the attached bacterial community with O. borgei revealed the influential environmental factors affecting the community (Figure 6). The black arrows represented nitrogen concentration (NC) and chlorophyll a concentration (Chl a) had similar length, suggesting their substantial impact on the attached bacterial community (Figure 6). The vertical distance from the samples to the black arrows represented the intensity of the environmental factor’s effect on the samples, with closer distances indicating a stronger influence. When the sample points were connected to the origin and formed an angle with the environmental factor arrows, the acute angles represented a positive correlation, while the obtuse angles indicated a negative correlation. The blue dashed lines represented various bacterial species, and the smaller the angle formed with the environmental factors, the higher the correlation. Significantly positive correlations were observed between chlorophyll a content, nitrogen concentration, and bacterial species such as Marinobacter, Rhodobacteraceae (unclassified genus), Hyphomonas, Algoriphagus, and Hoeflea. Conversely, there were significant negative correlations with bacterial species such as small pear-shaped bacteria, Cryomorphaceae, Mameliella, and Dinoroseobacters. The nitrogen concentration factor showed a positive correlation with species changes in the groups with 10 and 50 mg/L nitrogen, and a negative correlation with the groups with 2.5 and 100 mg/L nitrogen. On the other hand, chlorophyll a exhibited a positive correlation with species changes in the groups with 50 and 100 mg/L nitrogen and a negative correlation with the groups supplied with 2.5 and 10 mg/L nitrogen.
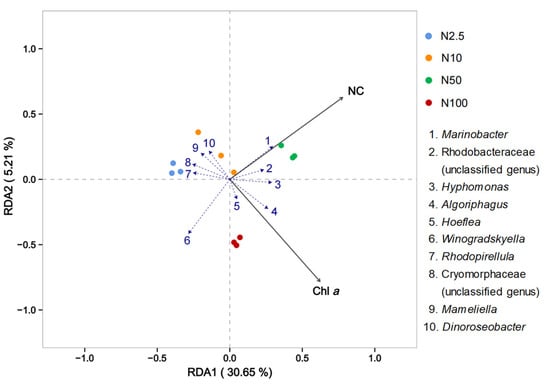
Figure 6.
RDA analysis at the genus level of the bacterial communities associated with O. borgei. Black arrows originating from the origin represent different environmental factors (NC = nitrogen concentration, Chl a = chlorophyll a concentration), with the length of the arrow indicating the strength of their impact on community changes. Longer arrows indicate greater influence.
4. Discussion
In this study, the chlorophyll a content of O. borgei showed a significant increase with increasing nitrogen concentrations. Similar results have been reported in related studies, suggesting that nitrogen limitation may directly influence the growth and chlorophyll a synthesis of O. borgei [21]. Nitrogen is closely associated with the dark reaction phase of microalgal photosynthesis and cell division, and nitrogen limitation can lead to an extended G1 phase [23,31].
The relative abundance of dominant bacterial populations associated with O. borgei at different nitrogen concentrations revealed that a low level of nitrogen favored the predominance of the genus Winogradskyella, while the medium and high nitrogen groups showed the community dominated by the genera Marinobacter and Winogradskyella. Although the relative abundance of Algoriphagus in the epiphytic microbial with O. borgei was low, it significantly increased under high nitrogen concentrations. Tujula et al. [9] found that bacteria belonging to the family Rhodobacteraceae and the genus Dinoroseobacter belonged to the predominant groups in the epibiont community of Ulvacean algae, whereas their abundance was relatively low in O. borgei at different nitrogen concentrations. The bacterial population of the family Rhodobacteraceae is known to be a key player in the initial formation of biofilms and may promote denitrification in nitrogen-polluted environments, leading to the formation of biofilms on leaf surfaces and an increase in community diversity. It is speculated that the above interaction between Rhodobacteraceae (unclassified genus) and microalgae may be the reason why its relative abundance is higher at high nitrogen concentrations. Furthermore, the bacterial density in the biofilm is much higher than that in the surrounding water.
Winogradskyella, a Gram-negative bacterial genus, was found to be the most abundant group in the O. borgei, and members of this genus have been previously reported in the epibionts of Ulva fenestrate [32]. Similar to the findings of this study, a negative correlation was observed between nitrogen concentrations and the presence of Winogradskyella [33]. Zhu et al. [34] found that Mameliella were widespread in marine dinoflagellates and suggested that these bacteria provide essential nutrients, such as vitamin B12, to promote algal growth. This study also identified the presence of Mameliella, which showed a significant negative correlation with nitrogen concentration. These results indicated that the dominant bacterial groups associated with microalgae cultured under different nitrogen concentrations varied accordingly.
The functional interactions within epibiont communities have a significant impact on algal growth [13,35,36]. Competition is very common between microalgae and bacteria, while some bacteria can also benefit from microalgae. Aerobic bacteria and chemoheterotrophic bacteria compete with microalgae for oxygen and nutrients [37]. For example, due to the preference of bacteria for utilizing ammonium nitrogen as a nitrogen source [38], bacteria from the family Rhodobacteraceae play a crucial role in nitrogen removal and denitrification processes, and this leads to more prominent denitrification and nitrogen metabolism function in the epibiotic environment, which is beneficial for the rapid uptake of ammonium nitrogen and the decomposition of nitrate nitrogen [39], which is in accordance with the results of O. borgei to effectively absorb ammonium. In contrast to competitive interactions, Algoriphagus can form a synergistic relationship with O. borgei and promote the accumulation of chlorophyll a [40]. The genus Marinobacter has the ability to secrete ammonium nitrogen, thereby promoting algal growth and chlorophyll a synthesis [41,42]. Ling et al. found that Marinobacter can promote the growth of seven microalgal species [43]. The exchange of nutrients between these algae and bacteria may be closely related to the significant nitrogen metabolism observed in this study.
Environmental factors also have a significant impact on the growth of microalgae and the composition of their epibiont communities [44,45,46]. Pei et al. [47] compared the environmental factors, such as nitrogen, temperature, pH, salinity, and dissolved oxygen, and found that nitrogen had a key influence on the epibiont community of Gracilariopsis lemaneiformis. Meanwhile, Ma et al. [24] investigated the symbiotic system of O. borgei and bacteria and found that temperature had the greatest impact on nitrite uptake rate, followed by salinity, while light intensity had the smallest impact. This study found that at high nitrogen concentrations, both ammonium nitrogen and chlorophyll a were positively correlated with changes in the epibiont community and diversity of O. borgei. The results show that both nitrogen concentration and chlorophyll a had significant effects on the bacteria with high relative abundance. Specifically, there was a positive correlation between species changes in the groups with 10 and 50 mg/L nitrogen, while a negative correlation was found with the groups supplied with 2.5 and 100 mg/L nitrogen. On the other hand, chlorophyll a exhibited a positive correlation with species changes in the groups with 50 and 100 mg/L nitrogen and a negative correlation with the groups with 2.5 and 10 mg/L nitrogen. It should be noted that the concentration of nitrogen showed a positive correlation with Marinobacter and Rhodobacteraceae. Therefore, increasing the addition of ammonia nitrogen in the process of regulating water quality for L. vannamei culture using O. borgei may be more beneficial for the development of the microalgal community.
Although the interactions between phytoplankton and bacteria occur on a small scale, they have a large-scale impact on primary production and carbon cycling in aquatic ecosystems. A more comprehensive understanding of these interactions, including different phytoplankton groups, will enhance our ability to model and predict marine environmental changes. This study demonstrated that the bacterial community associated with O. borgei included dominant genera such as Marinobacter and Winogradskyella under nitrogen-limited and nitrogen-replete conditions. However, the relationship between microalgae and bacteria is a co-evolutionary one, and further research is needed to explore their more intimate connections, such as information exchange, nutritional complementarity, and gene transfer, as well as the functional role of the epibiotic bacterial community associated with O. borgei. Considering the significant role of O. borgei in aquaculture for water quality regulation and its widespread application, understanding its bacterial community and functions becomes an important aspect for applied research.
5. Conclusions
The species composition of the attached bacterial community of O. borgei remained largely consistent under different nitrogen concentrations, but the relative abundance exhibited significant variations. The attached bacteria of O. borgei were all Gram-negative, with the majority being aerobic bacteria. Moreover, the attached bacterial community exhibited significant nitrogen cycling and denitrification functions, which were closely related to the characteristics of O. borgei, effectively assimilating ammonium nitrogen and nitrate nitrogen. There was a significant correlation among O. borgei, attached bacteria, and environmental factors. In particular, bacteria that may be beneficial to O. borgei, such as Rhodobacteraceae, Marinobacter, were positively correlated with nitrogen concentration, Winogradskyella was significantly negatively correlated with nitrogen concentration, while other bacteria were not significantly correlated with nitrogen concentration. This study elucidates specific bacteria with important functions in the attached bacterial community of O. borgei under different nitrogen concentration conditions, as well as the functional diversity of inter-algal bacterial communities, which provides valuable insights for further understanding the functions of O. borgei and utilizing O. borgei–algal consortia to maintain aquatic ecosystems.
Author Contributions
Conceptualization, Y.Z. and X.H.; methodology, B.L.; investigation, B.L.; data curation, B.L. and Y.Z.; writing—original draft preparation, B.L.; writing—review and editing, Z.H. and Y.Z.; supervision, C.L.; funding acquisition, Z.H. and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Modern Seed Industry Park for Whiteleg Shrimp of Guangdong Province (No. K22226), the Program for Scientific Research Start-up Funds of Guangdong Ocean University (Nos. 060302022102, 060302022201), and the Program of Shrimp Aquaculture Talent Development (No. B23312).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors greatly appreciate the constructive advice of anonymous reviewers.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Seymour, J.R.; Amin, S.A.; Raina, J.-B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef]
- Florez, J.Z.; Camus, C.; Hengst, M.B.; Marchant, F.; Buschmann, A.H. Structure of the epiphytic bacterial communities of macrocystis pyrifera in localities with contrasting nitrogen concentrations and temperature. Algal Res. 2019, 44, 101706. [Google Scholar] [CrossRef]
- You, X.; Xu, N.; Yang, X.; Sun, W. Pollutants affect algae-bacteria interactions: A critical review. Environ. Pollut. 2021, 276, 116723. [Google Scholar] [CrossRef] [PubMed]
- Dogs, M.; Wemheuer, B.; Wolter, L.; Bergen, N.; Daniel, R.; Simon, M.; Brinkhoff, T. Rhodobacteraceae on the marine brown alga fucus spiralis are abundant and show physiological adaptation to an epiphytic lifestyle. Syst. Appl. Microbiol. 2017, 40, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Klotz, M.G. Diversity and evolution of bioenergetic systems involved in microbial nitrogen compound transformations. Biochim. Biophys. Acta BBA-Bioenerg. 2013, 1827, 114–135. [Google Scholar] [CrossRef] [PubMed]
- Mouget, J.-L.; Dakhama, A.; Lavoie, M.C.; de la Noüe, J. Algal growth enhancement by bacteria: Is consumption of photosynthetic oxygen involved? FEMS Microbiol. Ecol. 1995, 18, 35–43. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Piampiano, E.; Pini, F.; Biondi, N.; Pastorelli, R.; Giovannetti, L.; Viti, C. Analysis of microbiota in cultures of the green microalga Tetraselmis suecica. Eur. J. Phycol. 2019, 54, 497–508. [Google Scholar] [CrossRef]
- Tujula, N.A.; Crocetti, G.R.; Burke, C.; Thomas, T.; Holmström, C.; Kjelleberg, S. Variability and abundance of the epiphytic bacterial community associated with a green marine Ulvacean alga. ISME J. 2010, 4, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Lachnit, T.; Meske, D.; Wahl, M.; Harder, T.; Schmitz, R. Epibacterial community patterns on marine macroalgae are host-specific but temporally variable: Epiphytic communities on macroalgae. Environ. Microbiol. 2011, 13, 655–665. [Google Scholar] [CrossRef]
- Tam, N.F.Y.; Wong, Y.S. Wastewater nutrient removal by chlorella Pyrenoidosa and Pcenedesmus sp. Environ. Pollut. 1989, 58, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Apandi, N.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Gani, P.; Ibrahim, A.; Kassim, A.H.M. Scenedesmus biomass productivity and nutrient removal from wet market wastewater, a bio-kinetic study. Waste Biomass Valorization 2019, 10, 2783–2800. [Google Scholar] [CrossRef]
- Xia, P.; Yan, D.; Sun, R.; Song, X.; Lin, T.; Yi, Y. Community composition and correlations between bacteria and algae within epiphytic biofilms on submerged macrophytes in a plateau lake, southwest China. Sci. Total Environ. 2020, 727, 138398. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kan, J.; He, C.; Shi, Q.; Liu, Y.-X.; Fan, Z.-C.; Sun, J. Epiphytic bacteria are essential for the production and transformation of algae-derived carboxyl-rich alicyclic molecule (CRAM)-like DOM. Microbiol. Spectr. 2021, 9, e01531-21. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-del-Valle, M.; Vílchez, C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Suzuki, M.; Kasai, H.; Shizuri, Y.; Harayama, S. Isolation and phylogenetic characterization of bacteria capable of inducing differentiation in the green alga Monostroma oxyspermum. Environ. Microbiol. 2003, 5, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, X.-H.; Li, C.-L.; Gu, B. Study on the uptake of dissolved nitrogen by Oocystis borgei in prawn (Litopenaeus vannamei) aquaculture ponds and establishment of uptake model. Aquac. Int. 2020, 28, 1445–1458. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Li, C.; Huang, X.; Li, F.; Wang, X.; Li, G. Allelopathic effect of Oocystis borgei culture on Microcystis aeruginosa. Environ. Technol. 2022, 43, 1662–1671. [Google Scholar] [CrossRef]
- Huang, X.; Wei, S.; Zhou, M.; Jiang, D. Study on the tolerance and adsorption of Cu2+ and Zn2+ by Oocystis borgei. J. Shanghai Ocean Univ. 2012, 21, 374–381. [Google Scholar]
- Salim, A. The Complete Plastome of the Green Alga Oocystis borgei. Master’s Dissertation, The University of Texas at Austin, Austin, TX, USA, 2023. [Google Scholar]
- Xie, L. The Impact of Carbon and Nitrogen on the Biochemical Composition, Sedimentation, and Metabolic Groups of Oocystis borgei. Master’s Dissertation, Guangdong Ocean University, Zhanjiang, China, 2020. [Google Scholar] [CrossRef]
- Liu, M.; Huang, X.; Zhang, R.; Li, C.; Gu, B. Uptake of urea nitrogen by Oocystis borgei in prawn (litopenaeus vannamei) aquaculture ponds. Bull. Environ. Contam. Toxicol. 2018, 101, 586–591. [Google Scholar] [CrossRef]
- Yang, M.; Li, C.; Huang, X.; Wang, Q.; Chen, A. The influence of nitrogen concentration, temperature, and light intensity on the reproductive patterns of Oocystis borgei. J. Guangdong Ocean Univ. 2019, 39, 44–49. [Google Scholar]
- Ma, Y.; Luo, Z.; Zhong, J.; Zhang, H.; Huang, X.; Li, C.; Zhang, Y. Effect of environmental factors on nitrite nitrogen absorption in microalgae–bacteria consortia of Oocystis borgei and Rhodopseudomonas palustris. Water 2023, 15, 1722. [Google Scholar] [CrossRef]
- Huang, X.; Li, X.; Wang, Y.; Zhou, M. Effects of environmental factors on the uptake rates of dissolved nitrogen by a salt-water green alga (Oocystis Borgei Snow). Bull. Environ. Contam. Toxicol. 2012, 89, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.Y.; Honda, K.; Derek, C.J.C. A review on microalgal-bacterial co-culture: The multifaceted role of beneficial bacteria towards enhancement of microalgal metabolite production. Environ. Res. 2023, 228, 115872. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Guo, L.; Wang, X.; Lin, Y.; Yang, X.; Ni, K.; Yang, F. Microorganisms that are critical for the fermentation quality of paper mulberry silage. Food Energy Secur. 2021, 10, e304. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. The Impact of Temperature, Light Intensity, and Nitrogen on the Reproductive Cycle of Oocystis borgei. Master’s Dissertation, Guangdong Ocean University, Zhanjiang, China, 2019. [Google Scholar]
- Nedashkovskaya, O.I.; Kukhlevskiy, A.D.; Zhukova, N.V. Winogradskyella Ulvae sp. nov., an epiphyte of a pacific seaweed, and emended descriptions of the genus Winogradskyella and Winogradskyella thalassocola, Winogradskyella echinorum, Winogradskyella exilis and Winogradskyella eximia. Int. J. Syst. Evol. Microbiol. 2012, 62, 1450–1456. [Google Scholar] [CrossRef]
- Cai, Z. Study on Microbial Community Characteristics of Main Seagrass Beds in Wenchang and the Impact of Two Types of Land-Based Pollutants and Vibrio on the Bacterial Community of Thalassia. Doctoral Dissertation, Hainan University, Haikou, China, 2021. [Google Scholar]
- Zhu, W.; Xu, F.; Ye, Y.; Yang, Q.; Zhang, X. Comparative genomics reveals insights into phylogenomic taxonomy and potential algae-bacteria interactions of novel versatile Mameliella alba strain LZ-28 Isolated from highly-toxic marine phycosphere. bioRxiv 2021. [Google Scholar] [CrossRef]
- Florez, J.Z.; Camus, C.; Hengst, M.B.; Buschmann, A.H. A functional perspective analysis of macroalgae and epiphytic bacterial community interaction. Front. Microbiol. 2017, 8, 2561. [Google Scholar] [CrossRef]
- Lian, J.; Wijffels, R.H.; Smidt, H.; Sipkema, D. The effect of the algal microbiome on industrial production of microalgae. Microb. Biotechnol. 2018, 11, 806–818. [Google Scholar] [CrossRef]
- González-Camejo, J.; Barat, R.; Pachés, M.; Murgui, M.; Seco, A.; Ferrer, J. Wastewater nutrient removal in a mixed microalgae–bacteria culture: Effect of light and temperature on the microalgae–bacteria competition. Environ. Technol. 2018, 39, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Benno, Y.; Mitsuoka, T. Utilization of ammonia nitrogen by intestinal bacteria isolated from pigs. Appl. Environ. Microbiol. 1980, 39, 30–35. [Google Scholar] [CrossRef]
- Qi, Y.; Zhong, Y.; Luo, L.; He, J.; Feng, B.; Wei, Q.; Zhang, K.; Ren, H. Subsurface constructed wetlands with modified biochar added for advanced treatment of tailwater: Performance and microbial communities. Sci. Total Environ. 2024, 906, 167533. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, X.; Zhu, C.; Mo, F.; Wen, Q.; Zhang, Y. Isolation and identification of a microalgal symbiotic bacterium strain and nitrogen uptake characteristics of the algal-bacterial consortium. J. Guangdong Ocean Univ. 2022, 42, 49–63. [Google Scholar] [CrossRef]
- Wei, X.; Shi, F.; Chen, Z.; Feng, J.; Zhu, L. Response of the triangle brown alga and its algal-bacterial consortia to different inorganic nitrogen sources. Mar. Sci. 2022, 46, 10–21. [Google Scholar] [CrossRef]
- Lupette, J.; Lami, R.; Krasovec, M.; Grimsley, N.; Moreau, H.; Piganeau, G.; Sanchez-Ferandin, S. Marinobacter dominates the bacterial community of the Ostreococcus tauri phycosphere in culture. Front. Microbiol. 2016, 7, 1414. [Google Scholar] [CrossRef]
- Ling, T.; Zhang, Y.F.; Cao, J.Y.; Xu, J.L.; Kong, Z.Y.; Zhang, L.; Liao, K.; Zhou, C.-X.; Yan, X.-J. Analysis of bacterial community diversity within seven bait-microalgae. Algal Res. 2020, 51, 102033. [Google Scholar] [CrossRef]
- Morales, M.; Sánchez, L.; Revah, S. The impact of environmental factors on carbon dioxide fixation by microalgae. FEMS Microbiol. Lett. 2018, 365, fnx262. [Google Scholar] [CrossRef]
- Moreno-Garcia, L.; Gariépy, Y.; Barnabé, S.; Raghavan, G.S.V. Effect of environmental factors on the biomass and lipid production of microalgae grown in wastewaters. Algal Res. 2019, 41, 101521. [Google Scholar] [CrossRef]
- Voolstra, C.R.; Ziegler, M. Adapting with microbial help: Microbiome flexibility facilitates rapid responses to environmental change. BioEssays 2020, 42, 2000004. [Google Scholar] [CrossRef] [PubMed]
- Pei, P.; Aslam, M.; Du, H.; Liang, H.; Wang, H.; Liu, X.; Chen, W. Environmental factors shape the epiphytic bacterial communities of Gracilariopsis lemaneiformis. Sci. Rep. 2021, 11, 8671. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).