
Soil Horizons Harbor Differing Fungal Communities

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Site Description
2.2. Experimental Design and Sampling
2.3. Soil Characteristics
2.4. DNA Isolation and Analysis
2.5. Bioinformatics and Statistical Analyses
2.6. α-Diversity
2.7. β-Diversity
2.8. Fungal Lifestyle
3. Results
3.1. Soil Characteristics
3.2. DNA Isolation and Analysis
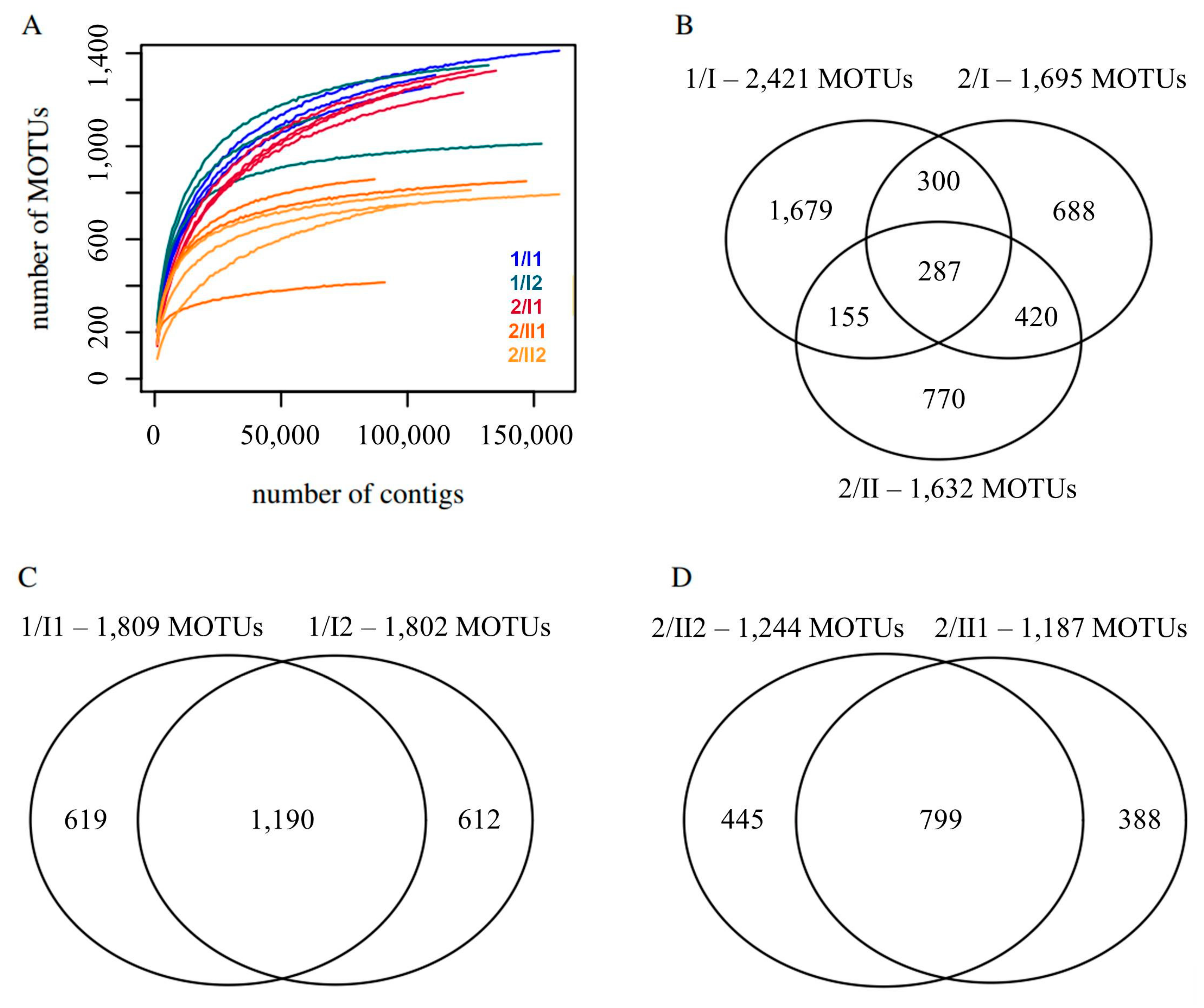
3.3. α-Diversity
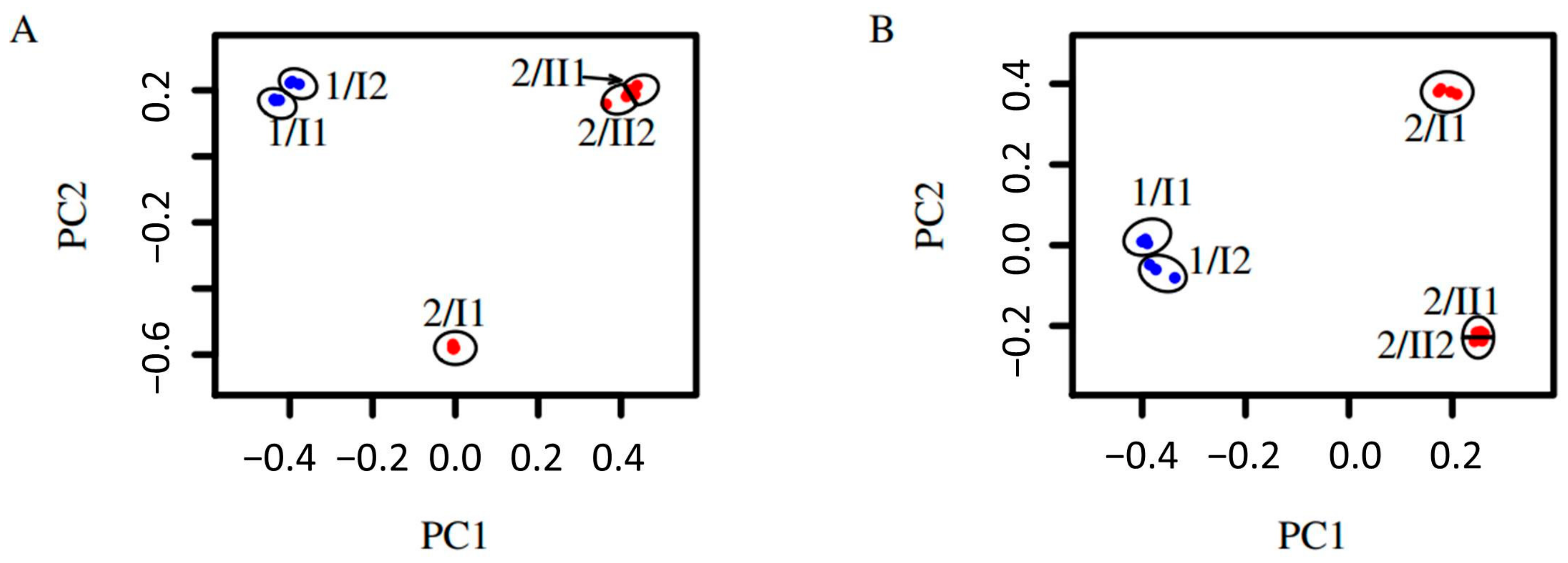
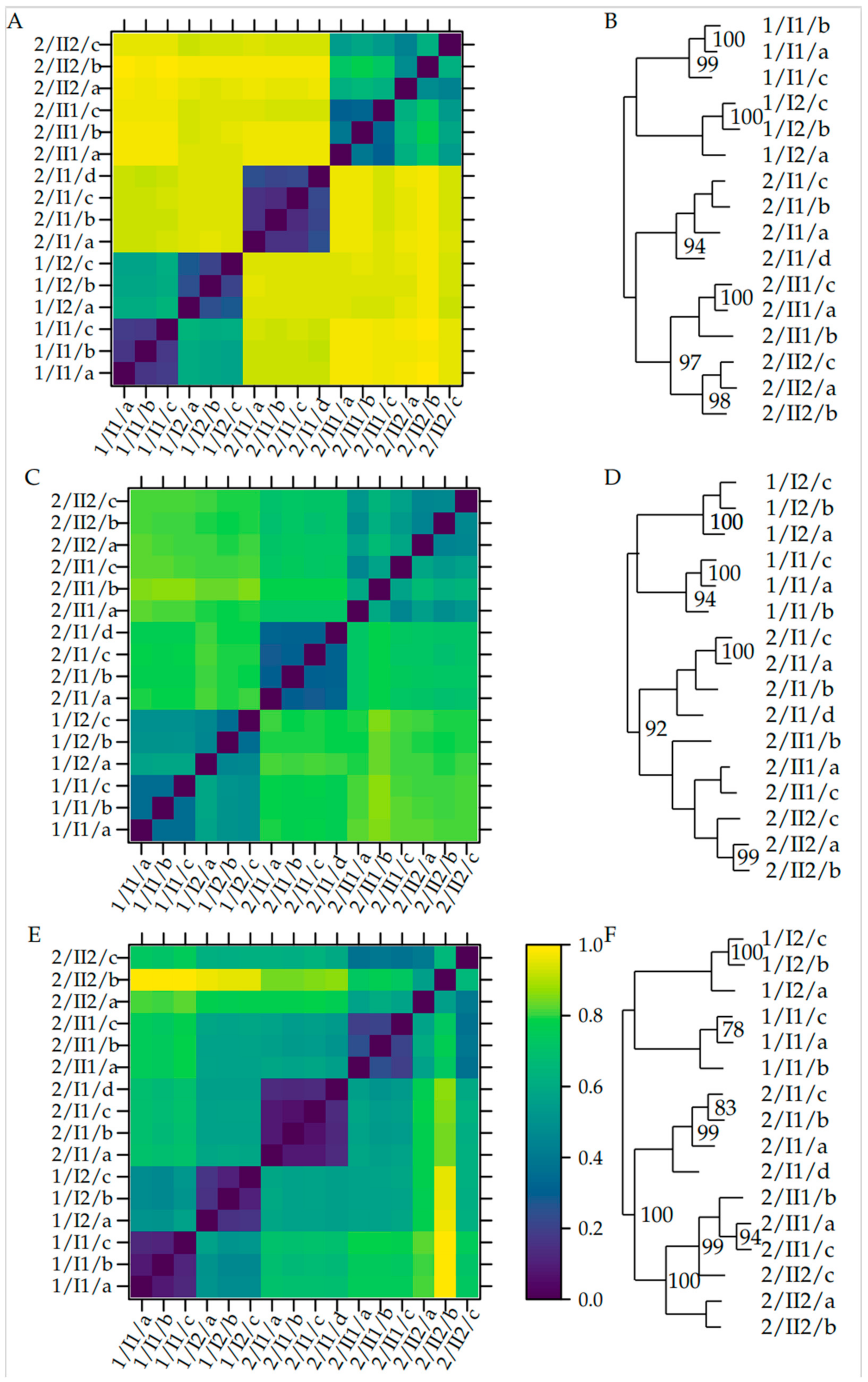
3.4. β-Diversity
3.5. Fungal Lifestyle
4. Discussion
4.1. Soil Characteristics
4.2. DNA Isolation and Analysis
4.3. α-Diversity
4.4. β-Diversity
4.5. Fungal Lifestyle
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Bosso, L.; Scelza, R.; Testa, A.; Cristinzio, G.; Rao, M.A. Depletion of pentachlorophenol contamination in an agricultural soil treated with Byssochlamys nivea, Scopulariopsis brumptii and urban waste compost: A laboratory microcosm study. Water Air Soil Pollut. 2015, 226, 183. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Zanne, A.E.; Abarenkov, K.; Afkhami, M.E.; Aguilar-Trigueros, C.A.; Bates, S.; Bhatnagar, J.M.; Busby, P.E.; Christian, N.; Cornwell, W.K.; Crowther, T.W.; et al. Fungal functional ecology: Bringing a trait-based approach to plant-associated fungi. Biol. Rev. 2020, 95, 409–433. [Google Scholar] [CrossRef]
- Jongmans, A.G.; van Breemen, N.; Lundström, U.; van Hees, P.A.W.; Finlay, R.D.; Srinivasan, M.; Unestam, T.; Giesler, R.; Melkerud, P.; Olsson, M. Rock-eating fungi. Nature 1997, 389, 682–683. [Google Scholar] [CrossRef]
- Ad-Hoc-AG Boden. Bodenkundliche Kartieranleitung 5 (KA5), 5th ed.; Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 2005. [Google Scholar]
- Zech, W.; Schad, P.; Hintermaier-Erhard, G. Böden der Welt: Ein Bildatlas, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- IUSS Working Group. World Reference Base (WRB) for Soil Resources 2014, Update 2015, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Food and Agriculture Organization: Rome, Italy, 2015; Available online: https://www.fao.org/3/i3794en/I3794en.pdf (accessed on 24 December 2023).
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Sharma, S.; Khare, P.K.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers. Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Warcup, J. The ecology of soil fungi. Trans. Br. Mycol. Soc. 1951, 34, 376–399. [Google Scholar] [CrossRef]
- Hamilton, A.C.; Taylor, D. History of climate and forests in tropical Africa during the last 8 million years. Clim. Change 1991, 19, 65–78. [Google Scholar] [CrossRef]
- Michel, P. Reliefgenerationen in Westafrika. In Beiträge zur Reliefgenese in Verschiedenen Klimazonen: Erweiterte Vorträge der Gordon-Konferenz 20–23.02.1975; Institut für Geographie der Universität Würzburg: Würzburg, Germany, 1977; pp. 111–130. [Google Scholar]
- Louis, H. Allgemeine Geomorphologie: Textteil und Gesonderter Bilderteil, 4th ed.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 1979. [Google Scholar]
- Porembski, S.; Barthlott, W. Granitic and gneissic outcrops (inselbergs) as centers of diversity for desiccation-tolerant vascular plants. Plant Ecol. 2000, 151, 19–28. [Google Scholar] [CrossRef]
- Porembski, S.; Barthlott, W. Inselbergs: Biotic Diversity of Isolated Rock Outcrops in Tropical and Temperate Regions; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2000; Volume 146, pp. 1–524. [Google Scholar] [CrossRef]
- Martin, J.P. Use of acid, rose bengal, and streptomycin in the plate method for estimating soil fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Bridge, P.; Spooner, B. Soil fungi: Diversity and detection. Plant Soil 2001, 232, 147–154. [Google Scholar] [CrossRef]
- Baldrian, P.; Kolařík, M.; Štursová, M.; Kopecký, J.; Valášková, V.; Větrovský, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, Č.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Hollingsworth, T.N.; McFarland, J.W.; Lennon, N.J.; Nusbaum, C.; Ruess, R.W. A first comprehensive census of fungi in soil reveals both hyperdiversity and fine-scale niche partitioning. Ecol. Monogr. 2014, 84, 3–20. [Google Scholar] [CrossRef]
- Khokon, A.M.; Schneider, D.; Daniel, R.; Polle, A. Soil layers matter: Vertical stratification of root-associated fungal assemblages in temperate forests reveals differences in habitat colonization. Microorganisms 2021, 9, 2131. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Liu, K.; Shen, Y.; Yao, G.; Yang, W.; Mortimer, P.E.; Gui, H. Fungal community composition and diversity vary with soil horizons in a subtropical forest. Front. Microbiol. 2021, 12, 650440. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Drenkhan, R.; Pritsch, K.; Buegger, F.; Padari, A.; Hagh-Doust, N.; Mikryukov, V.; Gohar, D.; et al. Regional-scale in-depth analysis of soil fungal diversity reveals strong pH and plant species effects in Northern Europe. Front. Microbiol. 2020, 11, 1953. [Google Scholar] [CrossRef]
- Voříšková, J.; Brabcová, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef]
- Yang, T.; Adams, J.M.; Shi, Y.; Sun, H.; Cheng, L.; Zhang, Y.; Chu, H. Fungal community assemblages in a high elevation desert environment: Absence of dispersal limitation and edaphic effects in surface soil. Soil Biol. Biochem. 2017, 115, 393–402. [Google Scholar] [CrossRef]
- Soderstrom, B.E.; Baath, E. Soil microfungi in three Swedish coniferous forests. Ecography 1978, 1, 62–72. Available online: https://www.jstor.org/stable/3682101 (accessed on 12 December 2023). [CrossRef]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Rudolph, S.; Maciá-Vicente, J.; Lotz-Winter, H.; Schleuning, M.; Piepenbring, M. Temporal variation of fungal diversity in a mosaic landscape in Germany. Stud. Mycol. 2018, 89, 95–104. [Google Scholar] [CrossRef]
- van der Linde, S.; Suz, L.M.; Orme, C.D.L.; Cox, F.; Andreae, H.; Asi, E.; Atkinson, B.; Benham, S.; Carroll, C.; Cools, N.; et al. Environment and host as large-scale controls of ectomycorrhizal fungi. Nature 2018, 558, 243–248. [Google Scholar] [CrossRef]
- Guerra, C.A.; Heintz-Buschart, A.; Sikorski, J.; Chatzinotas, A.; Guerrero-Ramírez, N.; Cesarz, S.; Beaumelle, L.; Rillig, M.C.; Maestre, F.T.; Delgado-Baquerizo, M.; et al. Blind spots in global soil biodiversity and ecosystem function research. Nat. Commun. 2020, 11, 3870. [Google Scholar] [CrossRef]
- Nguyen, D.Q.; Schneider, D.; Brinkmann, N.; Song, B.; Janz, D.; Schöning, I.; Daniel, R.; Pena, R.; Polle, A. Soil and root nutrient chemistry structure root-associated fungal assemblages in temperate forests. Environ. Microbiol. 2020, 22, 3081–3095. [Google Scholar] [CrossRef] [PubMed]
- Piepenbring, M.; Maciá-Vicente, J.G.; Codjia, J.E.I.; Glatthorn, C.; Kirk, P.; Meswaet, Y.; Minter, D.; Olou, B.A.; Reschke, K.; Schmidt, M.; et al. Mapping mycological ignorance—Checklists and diversity patterns of fungi known for West Africa. IMA Fungus 2020, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Meidl, P.; Furneaux, B.; Tchan, K.I.; Kluting, K.; Ryberg, M.; Guissou, M.-L.; Soro, B.; Traoré, A.; Konomou, G.; Yorou, N.S.; et al. Soil fungal communities of ectomycorrhizal dominated woodlands across West Africa. MycoKeys 2021, 81, 45–68. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22/21, 5271–5277. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Sahraei, S.E.; Furneaux, B.; Kluting, K.; Zahieh, M.; Rydin, H.; Hytteborn, H.; Rosling, A. Soil eukaryote community shift but not composition is consistently recovered by different OTU inference methods applied to long read metabarcoding data. Authorea 2021. Preprints. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Köhn, M. Korngrößenbestimmung mittels Pipettanalyse. Tonindustrie-Zeitung 1929, 55, 729–731. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Masoudi, A.; Wang, M.; Zhang, X.; Wang, C.; Qiu, Z.; Wang, W.; Wang, H.; Liu, J. Meta-analysis and evaluation by insect-mediated baiting reveal different patterns of Hypocrealean entomopathogenic fungi in the soils from two regions of China. Front. Microbiol. 2020, 11, 1133. [Google Scholar] [CrossRef] [PubMed]
- Babraham Institute. Fastqc: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 4 March 2018).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.; Weber, N. Microbial community analysis using MEGAN. Methods Enzymol. 2013, 531, 465–485. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Junge, B. Die Böden des Oberen Ouémé-Einzugsgebietes in Benin/Westafrika. Ph.D. Thesis, University Bonn, Bonn, Germany, 2004. [Google Scholar]
- Junge, B.; Skowronek, A. Genesis, properties, classification and assessment of soils in Central Benin, West Africa. Geoderma 2007, 139, 357–370. [Google Scholar] [CrossRef]
- Hiepe, C. Soil Degradation by Water Erosion in a Sub-Humid West-African Catchment: A Modelling Approach Considering Land Use and Climate Change in Benin. Ph.D. Thesis, Rheinische Friedrich Wilhelms-Universität Bonn, Bonn, Germany, 2008. [Google Scholar]
- Piepenbring, M.; Hofmann, T.A.; Unterseher, M.; Kost, G. Species richness of plants and fungi in western Panama: Towards a fungal inventory in the tropics. Biodivers. Conserv. 2012, 21, 2181–2193. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Global species numbers of fungi: Are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers. Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Rossman, A.Y. Protocols for an All Taxa Biodiversity Inventory of Fungi in a Costa Rican Conservation Area; Parkway Publishers, Inc.: Boone, NC, USA, 1998. [Google Scholar] [CrossRef]
- Antonín, V. Supplements to the monograph of tropical african species of Marasmius (Basidiomycota, Marasmiaceae). Cryptogam. Mycol. 2013, 34, 113–135. [Google Scholar] [CrossRef]
- Parmentier, I.; Oumorou, M.; Porembski, S.; Lejoly, J.; Decocq, G. Ecology, distribution, and classification of xeric monocotyledonous mats on inselbergs in West Africa and Atlantic central Africa. Phytocoenologia 2006, 36, 547–564. [Google Scholar] [CrossRef]
- Oumorou, M.; Lejoly, J. Écologie, flore et végétation de l’inselberg Sobakpérou (nord-Bénin). Acta Bot. Gallica 2003, 150, 65–84. [Google Scholar] [CrossRef]
- Frahm, J.-P.; Porembski, S. Moose von Inselbergen in Benin. Trop. Bryol. 1998, 14, 3–10. [Google Scholar]
- Giertz, S. Analyse der Hydrologischen Prozesse in den Sub-Humiden Tropen Westafrikas unter Besonderer Berücksichtigung der Landnutzung am Beispiel des Aguima-Einzugsgebietes in Benin. Ph.D. Thesis, University Bonn, Bonn, Germany, 2004. [Google Scholar]
- Niu, B.; Fu, G. Response of plant diversity and soil microbial diversity to warming and increased precipitation in alpine grasslands on the Qinghai-Xizang Plateau—A review. Sci. Total Environ. 2023, 912, 168878. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.; Espelta, J.M.; Morillo, J.A.; Pino, J.; Rodríguez-Echeverría, S. Land-use history alters the diversity, community composition and interaction networks of ectomycorrhizal fungi in beech forests. J. Ecol. 2021, 109, 2856–2870. [Google Scholar] [CrossRef]
- Guerra, C.A.; Bardgett, R.D.; Caon, L.; Crowther, T.W.; Delgado-Baquerizo, M.; Montanarella, L.; Navarro, L.M.; Orgiazzi, A.; Singh, B.K.; Tedersoo, L.; et al. Tracking, targeting, and conserving soil biodiversity. Science 2021, 371, 239–241. [Google Scholar] [CrossRef]
- Bâ, A.M.; Duponnois, R.; Moyersoen, B.; Diédhiou, A.G. Ectomycorrhizal symbiosis of tropical African trees. Mycorrhiza 2012, 22, 1–29. [Google Scholar] [CrossRef]
- Sprent, J.I.; James, E.K. Legume evolution: Where do nodules and mycorrhizas fit in? Plant Physiol. 2007, 144, 575–581. [Google Scholar] [CrossRef]
- Davison, J.; de León, D.G.; Zobel, M.; Moora, M.; Bueno, C.G.; Barceló, M.; Gerz, M.; León, D.; Meng, Y.; Pillar, V.D.; et al. Plant functional groups associate with distinct arbuscular mycorrhizal fungal communities. New Phytol. 2020, 226, 1117–1128. [Google Scholar] [CrossRef]
- Davison, J.; Moora, M.; Semchenko, M.; Adenan, S.B.; Ahmed, T.; Akhmetzhanova, A.A.; Alatalo, J.M.; Al-Quraishy, S.; Andriyanova, E.; Anslan, S.; et al. Temperature and pH define the realised niche space of arbuscular mycorrhizal fungi. New Phytol. 2021, 231, 763–776. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Sample | pH (%) | Sand (%) | Silt (%) | Clay (%) |
|---|---|---|---|---|---|
| site 1 | 1/I1 | 6.3 | 66.64 | 24.68 | 8.68 |
| 1/I2 | 6.1 | 70.85 | 22.71 | 6.44 | |
| site 2 | 2/I1 | 4.9 | 44.76 | 30.51 | 24.73 |
| 2/II1 | 4.0 | 37.47 | 27.33 | 35.19 | |
| 2/II2 | 4.0 | 34.28 | 29.03 | 36.69 |
| Taxonomic Level | Number of mOTUs | Fraction of Total mOTUs [%] |
|---|---|---|
| species | 75 | 1.7 |
| genus | 713 | 27 |
| family | 1285 | 30 |
| order | 1789 | 42 |
| class | 2050 | 48 |
| division | 2072 | 48 |
| unassigned | 2227 | 52 |
| total | 4299 | 100 |
| Order | Species | 1/I1 | 1/I2 | 2/I1 | 2/II1 | 2/II2 |
|---|---|---|---|---|---|---|
| Agaricales | Marasmius ferruginoides | + | ||||
| Calcarisporiellales | Calcarisporiella thermophila | + | + | |||
| Cladochytriales | Nowakowskiella elegans | + | + | + | + | + |
| Eurotiales | Aspergillus tamarii | + | ||||
| Eurotiales | Penicillium ornatum | + | + | + | + | + |
| Glomerales | Claroideoglomus walkeri | + | + | + | + | + |
| Hypocreales | Beauveria felina | + | + | + | ||
| Mortierellales | Mortierella ambigua | + | + | |||
| Mucorales | Cunninghamella blakesleeana | + | + | + | ||
| Pleosporales | Falciformispora senegalensis | + | + | + | + | + |
| Pleosporales | Lepidosphaeria nicotiae | + | + | + | + | + |
| Pleosporales | Pseudoxylomyces elegans | + | + | + | + | |
| Ramicandelaberales | Ramicandelaber taiwanensis | + | + | |||
| Rhizophlyctidales | Rhizophlyctis rosea | + | + | + | + | |
| Rhytismatales | Lophodermium piceae | + | + | |||
| Sordariales | Humicola fuscoatra | + | + | + | + | + |
| Spizellomycetales | Spizellomyces dolichospermus | + | + | + | ||
| Tremellales | Saitozyma podzolica | + |
| Replicate | Total Number of mOTUs | Unassigned | Assigned | Saprotrophic | Animal Parasitic | Arbuscular Mycorrhizal | Lichenized | Ectomycorrhizal | Mycoparasitic | Plant Pathogenic |
|---|---|---|---|---|---|---|---|---|---|---|
| 1/I1/a | 1307 | 902 | 405 | 244 | 9 | 12 | 0 | 92 | 3 | 43 |
| 1/I1/b | 1415 | 982 | 433 | 264 | 6 | 11 | 0 | 99 | 2 | 46 |
| 1/I1/c | 1257 | 865 | 392 | 233 | 7 | 11 | 0 | 98 | 1 | 40 |
| 1/I2/a | 1011 | 783 | 228 | 171 | 6 | 5 | 0 | 18 | 0 | 25 |
| 1/I2/b | 1127 | 852 | 275 | 205 | 6 | 7 | 0 | 20 | 1 | 33 |
| 1/I2/c | 1349 | 1.029 | 320 | 227 | 7 | 8 | 0 | 30 | 1 | 42 |
| 2/I1/a | 1235 | 803 | 432 | 371 | 8 | 2 | 5 | 3 | 3 | 37 |
| 2/I1/b | 1327 | 869 | 458 | 397 | 7 | 1 | 5 | 4 | 2 | 39 |
| 2/I1/c | 1231 | 798 | 433 | 375 | 7 | 3 | 5 | 3 | 4 | 33 |
| 2/I1/d | 1327 | 862 | 465 | 396 | 7 | 2 | 5 | 6 | 3 | 43 |
| 2/II1/a | 858 | 655 | 203 | 168 | 4 | 6 | 0 | 5 | 1 | 17 |
| 2/II1/b | 415 | 332 | 83 | 68 | 0 | 3 | 0 | 0 | 1 | 11 |
| 2/II1/c | 850 | 659 | 191 | 163 | 4 | 4 | 0 | 3 | 1 | 14 |
| 2/II2/a | 796 | 594 | 202 | 163 | 5 | 3 | 0 | 4 | 1 | 24 |
| 2/II2/b | 752 | 538 | 214 | 176 | 4 | 3 | 0 | 4 | 1 | 24 |
| 2/II2/c | 812 | 627 | 185 | 150 | 5 | 5 | 1 | 2 | 1 | 19 |
| all | 4299 | 3221 | 1078 | 804 | 14 | 22 | 5 | 112 | 6 | 107 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mager, E.; Brockhage, R.; Piepenbring, M.; Segers, F.; Yorou, N.S.; Ebersberger, I.; Mangelsdorff, R.D. Soil Horizons Harbor Differing Fungal Communities. Diversity 2024, 16, 97. https://doi.org/10.3390/d16020097
Mager E, Brockhage R, Piepenbring M, Segers F, Yorou NS, Ebersberger I, Mangelsdorff RD. Soil Horizons Harbor Differing Fungal Communities. Diversity. 2024; 16(2):97. https://doi.org/10.3390/d16020097
Chicago/Turabian StyleMager, Enno, Ronja Brockhage, Meike Piepenbring, Francisca Segers, Nourou Soulemane Yorou, Ingo Ebersberger, and Ralph Daniel Mangelsdorff. 2024. "Soil Horizons Harbor Differing Fungal Communities" Diversity 16, no. 2: 97. https://doi.org/10.3390/d16020097
APA StyleMager, E., Brockhage, R., Piepenbring, M., Segers, F., Yorou, N. S., Ebersberger, I., & Mangelsdorff, R. D. (2024). Soil Horizons Harbor Differing Fungal Communities. Diversity, 16(2), 97. https://doi.org/10.3390/d16020097