
Investigation of Potential Effects of Ibuprofen on the Storage Cells and Anhydrobiosis Capacity of the Tardigrade Paramacrobiotus experimentalis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Experiment
2.2.2. Light and Transmission Electron Microscopy
2.2.3. Confocal Microscopy
TUNEL (Terminal Deoxynucleotidyl Transferase)—Detection of DNA Fragmentation during Apoptosis
LysoTracker Red—Detection of Acidic Organelles
Dihydroethidium (DHE) Analyzing Superoxide Levels (Differentiation of ROS+ and ROS− Cells)
Induction and Recovery from Anhydrobiosis
Statistical Analysis
3. Results
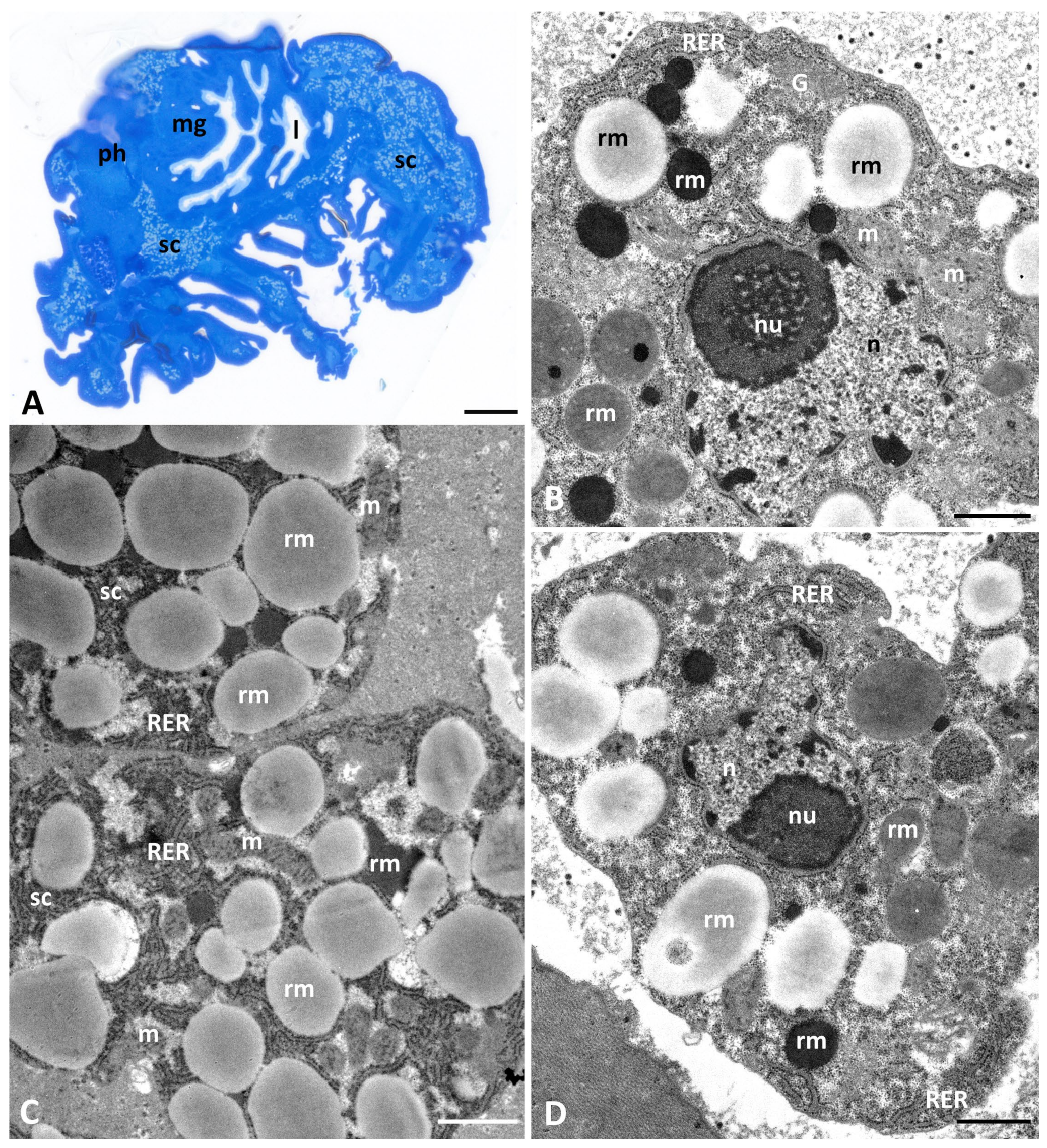
3.1. Ultrastructural Changes in Storage Cells of Pam. experimentalis Exposed to Ibuprofen
3.1.1. Control Groups (PeC, PeC0, PeC7, PeC28)
3.1.2. Experimental Groups after 7 Days of Exposure to Ibuprofen: Ib1a (Ibuprofen Concentration 0.1 μg/L), Ib1b (Ibuprofen Concentration 16.8 μg/L), Ib1c (Ibuprofen Concentration 1 mg/L)
3.1.3. Experimental Group Ib2a (Ibuprofen Concentration 0.1 μg/L, 28 Days)
3.1.4. Experimental Group Ib2b (Ibuprofen Concentration 16.8 μg/L, 28 Days)
3.1.5. Experimental Group Ib2c (Ibuprofen Concentration 1 mg/L, 28 Days)
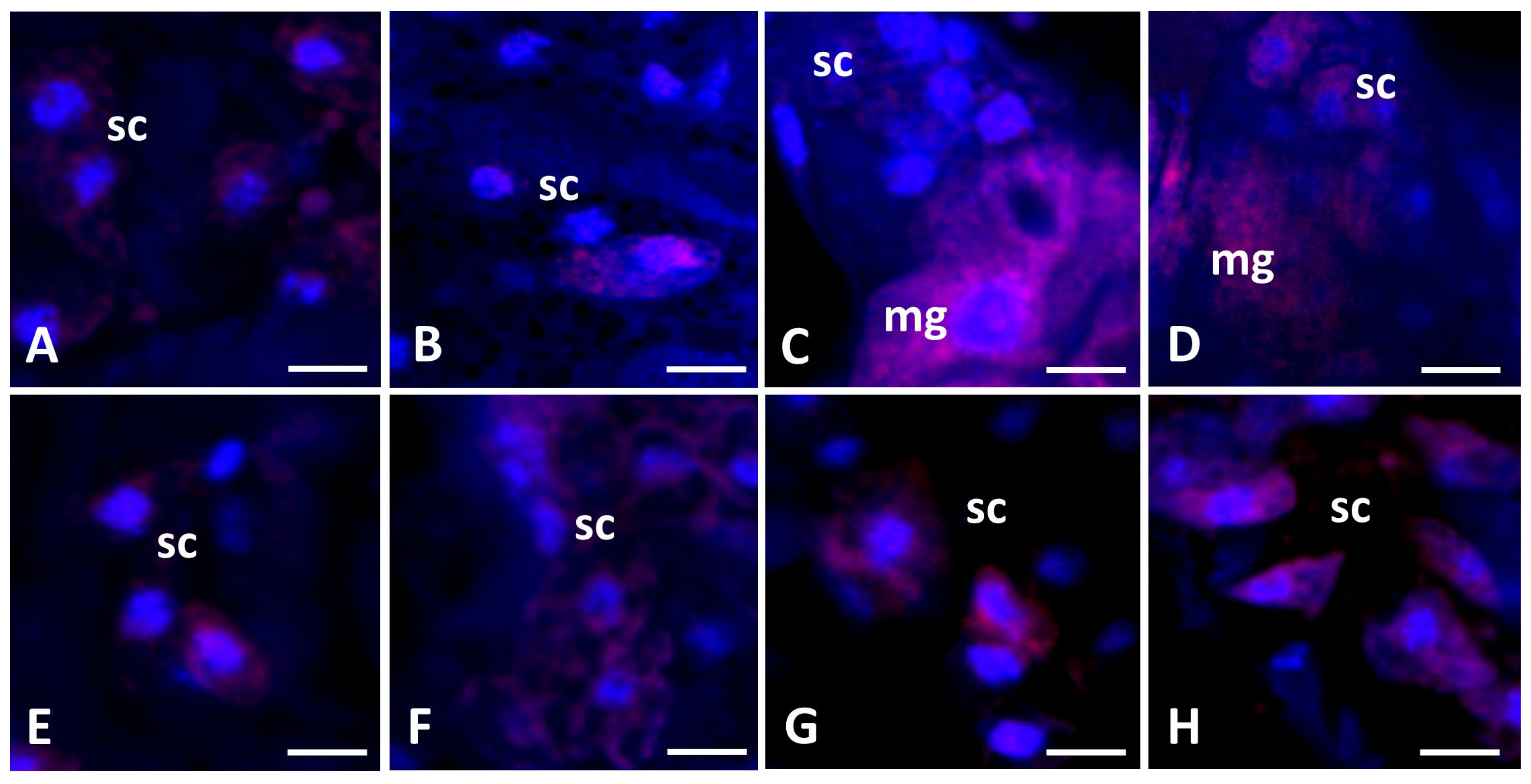
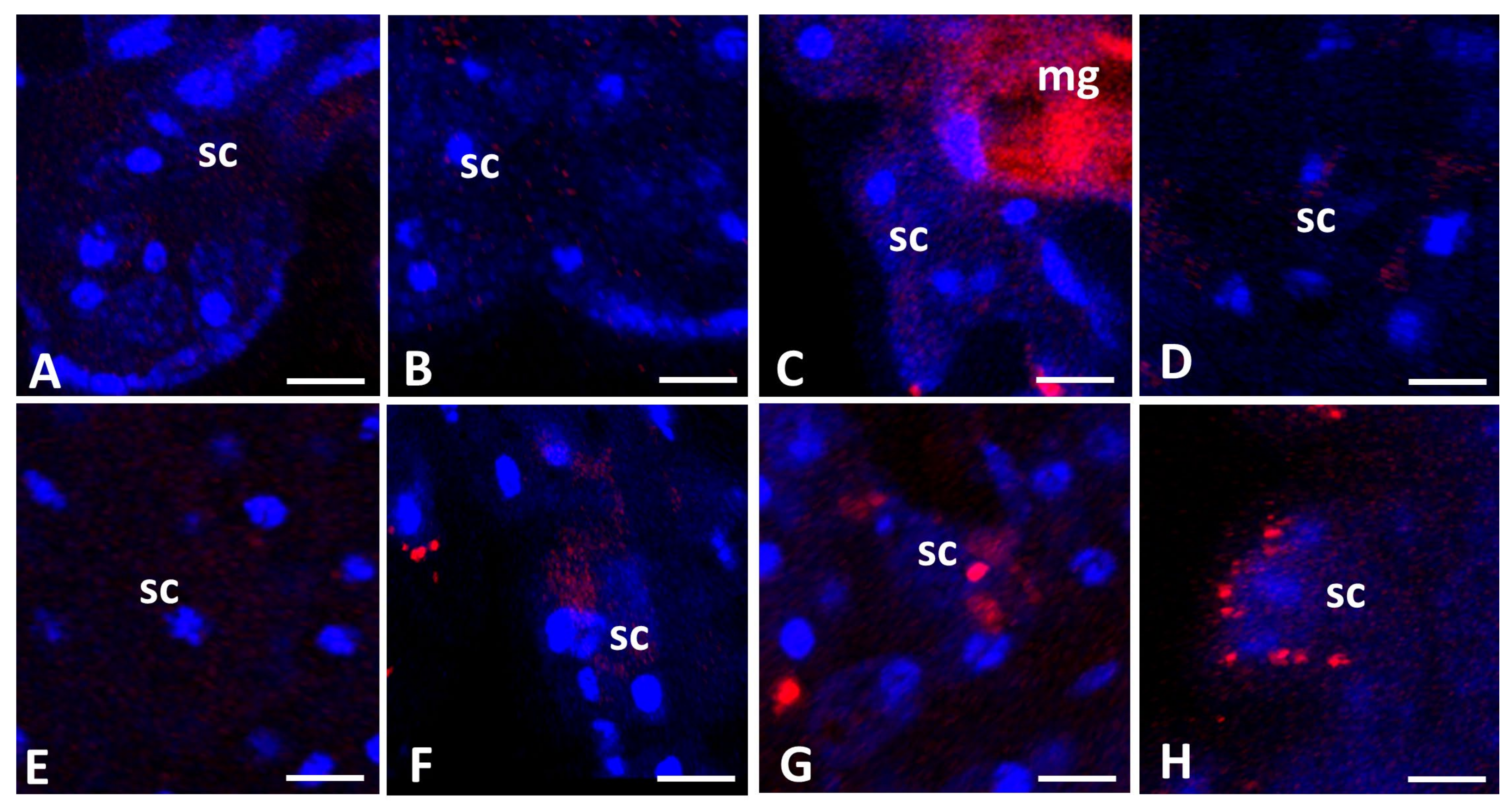
3.2. Confocal Microscopy Analysis
3.2.1. ROS+/ROS− Cells in Storage Cells of Pam. experimentalis Exposed to Ibuprofen
3.2.2. Apoptosis in Storage Cells of Pam. experimentalis Exposed to Ibuprofen
3.2.3. Autophagy in Storage Cells of Pam. experimentalis Exposed to Ibuprofen
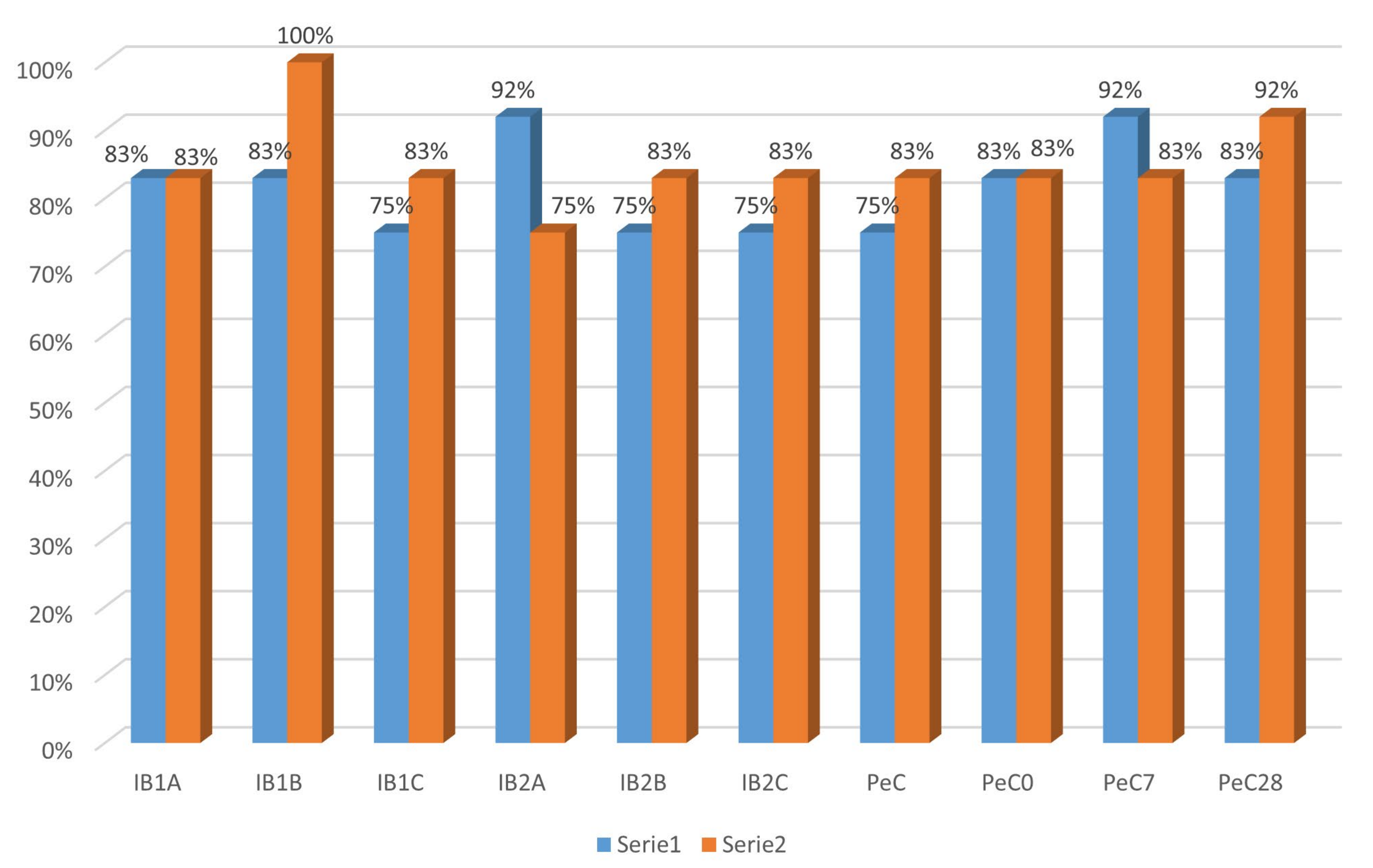
3.3. Recovery from Anhydrobiosis Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boxall, A.B.; Rudd, M.A.; Brooks, B.W.; Caldwell, D.J.; Choi, K.; Hickmann, S.; Innes, E.; Ostapyk, K.; Staveley, J.P.; Verslycke, T.; et al. Pharmaceuticals and personal care products in the environment: What are the big questions? Environ. Health Perspect. 2012, 120, 1221–1229. [Google Scholar] [CrossRef]
- Fent, K.; Weston, A.A.; Caminada, D. Ecotoxicology of human pharmaceuticals. Aquat. Toxicol. 2006, 76, 122–159. [Google Scholar] [CrossRef] [PubMed]
- Szymonik, A.; Lach, J. Zagrożenie środowiska wodnego obecnością środków farmaceutycznych. Inżynieria i Ochrona Środowiska 2012, 15, 249–263. [Google Scholar]
- Nikolaou, A.; Meric, S.; Fatta, D. Occurrence patterns of pharmaceuticals in water and wastewater environments. Anal. Bioanal. Chem. 2007, 387, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Harshkova, D.; Aksmann, A. Zanieczyszczenie środowiska niesteroidowymi lekami przeciwzapalnymi na przykładzie diklofenaku? Przyczyny, skutki, bioindykacja. Kosmos 2019, 68, 185–194. [Google Scholar] [CrossRef]
- Parolini, M. Toxicity of the Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) acetylsalicylic acid, paracetamol, diclofenac, ibuprofen and naproxen towards freshwater invertebrates: A review. Sci. Total Environ. 2020, 740, 140043. [Google Scholar] [CrossRef]
- Karaźniewicz, M. Czterdzieści lat doświadczeń stosowania ibuprofenu w lecznictwie. Nowiny Lekarskie 2007, 76, 195–197. [Google Scholar]
- Bushra, R.; Aslam, N. An overview of clinical pharmacology of Ibuprofen. Oman Med. J. 2010, 25, 155. [Google Scholar] [CrossRef]
- Stanley-Samuelson, D.W. Physiological roles of prostaglandins and other eicosanoids in invertebrates. Biol. Bull. 1987, 173, 92–109. [Google Scholar] [CrossRef]
- Batucan, N.S.P.; Tremblay, L.A.; Northcott, G.L.; Matthaei, C.D. Medicating the environment? A critical review on the risks of carbamazepine, diclofenac and ibuprofen to aquatic organisms. Environ. Adv. 2022, 7, 100164. [Google Scholar] [CrossRef]
- Nelson, D.R.; Guidetti, R.; Rebecchi, L. Phylum Tardigrada. In Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 347–380. [Google Scholar]
- Węglarska, B. Studies on the morphology of Macrobiotus richtersi Murray, 1911. Mem. Ist. Ital. Idrobiol. 1975, 32, 445–464. [Google Scholar]
- Poprawa, I. Ultrastructural changes of the storage cells during oogenesis in Dactylobiotus dispar (Murray, 1907) (Tardigrada: Eutardigrada). Zool. Pol. 2006, 51, 13–18. [Google Scholar]
- Reuner, A.; Hengherr, S.; Brümmer, F.; Schill, R.O. Comparative studies on storage cells in tardigrades during starvation and anhydrobiosis. Curr. Zool. 2010, 56, 259–263. [Google Scholar] [CrossRef]
- Czerneková, M.; Janelt, K.; Student, S.; Jönsson, K.I.; Poprawa, I. A comparative ultrastructure study of storage cells in the eutardigrade Richtersius coronifer in the hydrated state and after desiccation and heating stress. PLoS ONE 2018, 13, e0201430. [Google Scholar] [CrossRef]
- Crowe, J.H.; Madin, K.A. Anhydrobiosis in tardigrades and nematodes. Trans. Am. Micros. Soc. 1974, 93, 513–524. [Google Scholar] [CrossRef]
- Hengherr, S.; Brümmer, F.; Schill, R.O. Anhydrobiosis in tardigrades and its effects on longevity traits. J. Zool. 2008, 275, 216–220. [Google Scholar] [CrossRef]
- Wełnicz, W.; Grohme, M.A.; Kaczmarek, Ł.; Schill, R.O.; Frohme, M. Anhydrobiosis in tardigrades—The last decade. J. Insect Physiol. 2011, 57, 577–583. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Roszkowska, M.; Fontaneto, D.; Jezierska, M.; Pietrzak, B.; Wieczorek, R.; Poprawa, I.; Kosicki, J.Z.; Karachitos, A.; Kmita, H. Staying young and fit? Ontogenetic and phylogenetic consequences of animal anhydrobiosis. J. Zool. 2019, 309, 1–11. [Google Scholar] [CrossRef]
- Poprawa, I.; Chajec, Ł.; Chachulska-Żymełka, A.; Wilczek, G.; Student, S.; Leśniewska, M.; Rost-Roszkowska, M. Ovaries and testes of Lithobius forficatus (Myriapoda, Chilopoda) react differently to the presence of cadmium in the environment. Sci. Rep. 2022, 12, 6705. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Roszkowska, M.; Poprawa, I.; Janelt, K.; Kmita, H.; Gawlak, M.; Fiałkowska, E.; Mioduchowska, M. Integrative description of bisexual Paramacrobiotus experimentalis sp. nov. (Macrobiotidae) from republic of Madagascar (Africa) with microbiome analysis. Mol. Phylogenet. Evol. 2020, 145, 106730. [Google Scholar] [CrossRef]
- aus der Beek, T.; Weber, F.A.; Bergmann, A.; Hickmann, S.; Ebert, I.; Hein, A.; Küster, A. Pharmaceuticals in the environment—Global occurrences and perspectives. Environ. Toxicol. Chem. 2016, 35, 823–835. [Google Scholar] [CrossRef]
- Mrowiec, B. Farmaceutyki–zagrożenie środowiska wodnego. Edukacja Biologiczna i Środowiskowa 2015, 4, 25–33. [Google Scholar]
- Heckmann, L.H.; Connon, R.; Hutchinson, T.H.; Maund, S.J.; Sibly, R.M.; Callaghan, A. Expression of target and reference genes in Daphnia magna exposed to ibuprofen. BMC Genom. 2006, 7, 175. [Google Scholar] [CrossRef]
- Heckmann, L.H.; Callaghan, A.; Hooper, H.L.; Connon, R.; Hutchinson, T.H.; Maund, S.J.; Sibly, R.M. Chronic toxicity of ibuprofen to Daphnia magna: Effects on life history traits and population dynamics. Toxicol. Lett. 2007, 172, 137–145. [Google Scholar] [CrossRef]
- Hayashi, Y.; Heckmann, L.H.; Callaghan, A.; Sibly, R.M. Reproduction recovery of the crustacean Daphnia magna after chronic exposure to ibuprofen. Ecotoxicology 2008, 17, 246–251. [Google Scholar] [CrossRef]
- Gómez-Oliván, L.M.; Galar-Martínez, M.; García-Medina, S.; Valdés-Alanís, A.; Islas-Flores, H.; Neri-Cruz, N. Genotoxic response and oxidative stress induced by diclofenac, ibuprofen and naproxen in Daphnia magna. Drug Chem. Toxicol. 2014, 37, 391–399. [Google Scholar] [CrossRef]
- Pascoe, D.; Karntanut, W.; Müller, C.T. Do pharmaceuticals affect freshwater invertebrates? A study with the cnidarian Hydra vulgaris. Chemosphere 2003, 51, 521–528. [Google Scholar] [CrossRef]
- Parolini, M.; Binelli, A.; Provini, A. Chronic effects induced by ibuprofen on the freshwater bivalve Dreissena polymorpha. Ecotoxicol. Environ. Saf. 2011, 74, 1586–1594. [Google Scholar] [CrossRef]
- Mathias, F.T.; Fockink, D.H.; Disner, G.R.; Prodocimo, V.; Ribas, J.L.C.; Ramos, L.P.; Cestari, M.M.; de Assis, H.C.S. Effects of low concentrations of ibuprofen on freshwater fish Rhamdia quelen. Environ. Toxicol. Pharmacol. 2018, 59, 105–113. [Google Scholar] [CrossRef]
- Flippin, J.L.; Huggett, D.; Foran, C.M. Changes in the timing of reproduction following chronic exposure to ibuprofen in Japanese medaka, Oryzias latipes. Aquat. Toxicol. 2007, 81, 73–78. [Google Scholar] [CrossRef]
- Wieczorkiewicz, F.; Sojka, J.; Poprawa, I. Effect of paracetamol on the storage cells of Hypsibius exemplaris—Ultrastructural analysis. Zool. J. Linn. Soc. 2024, 200, 258–268. [Google Scholar] [CrossRef]
- Jönsson, K.I.; Rebecchi, L. Experimentally induced anhydrobiosis in the tardigrade Richtersius coronifer: Phenotypic factors affecting survival. J. Exp. Zool. 2002, 293, 578–584. [Google Scholar] [CrossRef]
- Hyra, M.; Rost-Roszkowska, M.M.; Student, S.; Włodarczyk, A.; Deperas, M.; Janelt, K.; Poprawa, I. Body cavity cells of Parachela during their active life. Zool. J. Linn. Soc. 2016, 178, 878–887. [Google Scholar] [CrossRef]
- Hyra, M.; Poprawa, I.; Włodarczyk, A.; Student, S.; Sonakowska, L.; Kszuk-Jendrysik, M.; Rost-Roszkowska, M.M. Ultrastructural changes in the midgut epithelium of Hypsibius dujardini (Doyère, 1840) (Tardigrada, Eutardigrada, Hypsibiidae) in relation to oogenesis. Zool. J. Linn. Soc. 2016, 178, 897–906. [Google Scholar] [CrossRef]
- Kamińska, K.; Lipovšek, S.; Kaszuba, F.; Rost-Roszkowska, M. Ultrastructure of the fat body in the soil centipedes Lithobius forficatus (Lithobiidae) and Geophilus flavus (Geophilidae) according to their seasonal rhythms. Zool. Anz. 2019, 279, 82–93. [Google Scholar] [CrossRef]
- Miernik, A.; Student, S.; Fiałkowska, E.; Poprawa, I. Does the midgut of the tardigrade Paramacrobiotus experimentalis responds to the effects of ibuprofen. Eur. Zool. J. 2023; under review. [Google Scholar]
- Rost-Roszkowska, M.M.; Poprawa, I.; Kaczmarek, Ł. Autophagy as the cell survival in response to a microsporidian infection of the midgut epithelium of Isohypsibius granulifer granulifer (Eutardigrada: Hypsibiidae). Acta Zool. 2013, 94, 273–279. [Google Scholar] [CrossRef]
- Yin, Z.; Pascual, C.; Klionsky, D.J. Autophagy: Machinery and regulation. Microbial Cell. 2016, 3, 588. [Google Scholar] [CrossRef]
- Rost-Roszkowska, M.M.; Vilimova, J.; Sosinka, A.; Skudlik, J.; Franzetti, E. The role of autophagy in the midgut epithelium of Eubranchipus grubii (Crustacea, Branchiopoda, Anostraca). Arthropod Struct. Dev. 2012, 41, 271–279. [Google Scholar] [CrossRef]
- Włodarczyk, A.; Student, S.; Rost-Roszkowska, M. Autophagy and apoptosis in starved and refed Neocaridina davidi (Crustacea Malacostraca) midgut. Can. J. Zool. 2019, 97, 294–303. [Google Scholar] [CrossRef]
- Ostróżka, A.; Tiffert, Z.; Wilczek, G.; Rost-Roszkowska, M. Can insecticide-free clean water regenerate the midgut epithelium of the freshwater shrimp after dimethoate treatment? Micron 2022, 155, 103162. [Google Scholar] [CrossRef]
- Rost-Roszkowska, M.M.; Janelt, K.; Poprawa, I. The role of autophagy in the midgut epithelium of Parachela (Tardigrada). Zoomorphology 2018, 137, 501–509. [Google Scholar] [CrossRef]
- Gąsiorek, P.; Stec, D.; Morek, W.; Michalczyk, Ł. An integrative redescription of Hypsibius dujardini (Doyère, 1840), the nominal taxon for Hypsibioidea (Tardigrada: Eutardigrada). Zootaxa 2018, 4415, 45–75. [Google Scholar] [CrossRef]
- Poprawa, I.; Hyra, M.; Kszuk-Jendrysik, M.; Rost-Roszkowska, M.M. Ultrastructural changes and programmed cell death of trophocytes in the gonad of Isohypsibius granulifer granulifer Thulin, 1928 (Tardigrada, Eutardigrada, Isohypsibiidae). Micron 2015, 70, 26–33. [Google Scholar] [CrossRef]
- Hetts, S.W. To die or not to die: An overview of apoptosis and its role in disease. Jama 1998, 279, 300–307. [Google Scholar] [CrossRef]
- Velentzas, A.D.; Nezis, I.P.; Stravopodis, D.J.; Papassideri, I.S.; Margaritis, L.H. Apoptosis and autophagy function cooperatively for the efficacious execution of programmed nurse cell death during Drosophila virilis oogenesis. Autophagy 2007, 3, 130–132. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Włodarczyk, A.; Wilczek, G.; Wilczek, P.; Student, S.; Ostróżka, A.; Tarnawska, M.; Rost-Roszkowska, M. Relationship between ROS production, MnSOD activation and periods of fasting and re-feeding in freshwater shrimp Neocaridina davidi (Crustacea, Malacostraca). PeerJ 2019, 7, e7399. [Google Scholar] [CrossRef]
- Chaitanya, R.K.; Shashank, K.; Sridevi, P. Oxidative stress in invertebrate systems. In Free Radicals and Diseases; Intech: Manchester, UK, 2016; Volume 19. [Google Scholar]
- Keilin, D. The Leeuwenhoek Lecture-the problem of anabiosis or latent life: History and current concept. Proc. Royal Soc. 1959, 150, 149–191. [Google Scholar]
- Alpert, P. The limits and frontiers of desiccation-tolerant life. Integr. Comp. Biol. 2005, 45, 685–695. [Google Scholar] [CrossRef]
- Roszkowska, M.; Gołdyn, B.; Wojciechowska, D.; Kosicki, J.Z.; Fiałkowska, E.; Kmita, H.; Kaczmarek, Ł. Tolerance to anhydrobiotic conditions among two coexisting tardigrade species differing in life strategies. Zool. Stud. 2021, 60, e74. [Google Scholar] [PubMed]
- Roszkowska, M.; Gołdyn, B.; Wojciechowska, D.; Księżkiewicz, Z.; Fiałkowska, E.; Pluskota, M.; Kmita, H.; Kaczmarek, Ł. How long can tardigrades survive in the anhydrobiotic state? A search for tardigrade anhydrobiosis patterns. PLoS ONE 2023, 18, e0270386. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Reuner, A.; Brümmer, F.; Schill, R.O. DNA damage in storage cells of anhydrobiotic tardigrades. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 153, 425–429. [Google Scholar] [CrossRef]
- Nagwani, A.K.; Melosik, I.; Kaczmarek, Ł.; Kmita, H. Recovery from anhydrobiosis in the tardigrade Paramacrobiotus experimentalis: Better to be young than old and in a group than alone. bioRxiv 2023. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PeC | specimens cultured in the culture medium |
| PeC0 | specimens cultured in the culture medium with the addition of 1 mL of ethanol per liter of medium |
| PeC7 | specimens cultured 7 days in the culture medium with the addition of 1 mL of ethanol per liter of medium |
| PeC28 | specimens cultured 28 days in the culture medium with the addition of 1 mL of ethanol per liter of medium |
| Ib1a | specimens cultured in ibuprofen at a concentration of 0.1 μg/L for 7 days |
| Ib1b | specimens cultured in ibuprofen at a concentration of 16.8 μg/L for 7 days |
| Ib1c | specimens cultured in ibuprofen at a concentration of 1 mg/L for 7 days |
| Ib2a | specimens cultured in ibuprofen at a concentration of 0.1 μg/L for 28 days |
| Ib2b | specimens cultured in ibuprofen at a concentration of 16.8 μg/L for 28 days |
| Ib2c | specimens cultured in ibuprofen at a concentration of 1 mg/L for 28 days |
| Light and Transmission Electron Microscopy | Confocal Microscopy | Induction and Recovery from Anhydrobiosis | |||
|---|---|---|---|---|---|
| TUNEL | LysoTracker Red | DHE | |||
| PeC | 20 | 10 | 10 | 10 | 24 |
| PeC0 | 20 | 10 | 10 | 10 | 24 |
| PeC7 | 20 | 10 | 10 | 10 | 24 |
| PeC28 | 20 | 10 | 10 | 10 | 24 |
| Ib1a | 20 | 10 | 10 | 10 | 24 |
| Ib1b | 20 | 10 | 10 | 10 | 24 |
| Ib1c | 20 | 10 | 10 | 10 | 24 |
| Ib2a | 20 | 10 | 10 | 10 | 24 |
| Ib2b | 20 | 10 | 10 | 10 | 24 |
| Ib2c | 20 | 10 | 10 | 10 | 24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miernik, A.; Wieczorkiewicz, F.; Student, S.; Poprawa, I. Investigation of Potential Effects of Ibuprofen on the Storage Cells and Anhydrobiosis Capacity of the Tardigrade Paramacrobiotus experimentalis. Diversity 2024, 16, 132. https://doi.org/10.3390/d16030132
Miernik A, Wieczorkiewicz F, Student S, Poprawa I. Investigation of Potential Effects of Ibuprofen on the Storage Cells and Anhydrobiosis Capacity of the Tardigrade Paramacrobiotus experimentalis. Diversity. 2024; 16(3):132. https://doi.org/10.3390/d16030132
Chicago/Turabian StyleMiernik, Aleksandra, Filip Wieczorkiewicz, Sebastian Student, and Izabela Poprawa. 2024. "Investigation of Potential Effects of Ibuprofen on the Storage Cells and Anhydrobiosis Capacity of the Tardigrade Paramacrobiotus experimentalis" Diversity 16, no. 3: 132. https://doi.org/10.3390/d16030132
APA StyleMiernik, A., Wieczorkiewicz, F., Student, S., & Poprawa, I. (2024). Investigation of Potential Effects of Ibuprofen on the Storage Cells and Anhydrobiosis Capacity of the Tardigrade Paramacrobiotus experimentalis. Diversity, 16(3), 132. https://doi.org/10.3390/d16030132