


Survival, Nest Site Affiliation and Post-Fledging Movements of Danish White-Tailed Eagles (Haliaeetus albicilla)

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Tagging and Transmitters
2.2. Data Filtering and Sampling
2.3. Precision of Location Data
2.4. Ageing and Sexing
2.5. Fledging
2.6. Survival
2.7. Nest Zone Affiliation
2.8. First Exploratory Flight
2.9. Dispersal
2.10. Distribution
3. Results
3.1. Fledging
3.2. Survival
3.3. Nest Zone Affiliation
3.4. First Exploratory Flight
3.5. Dispersal
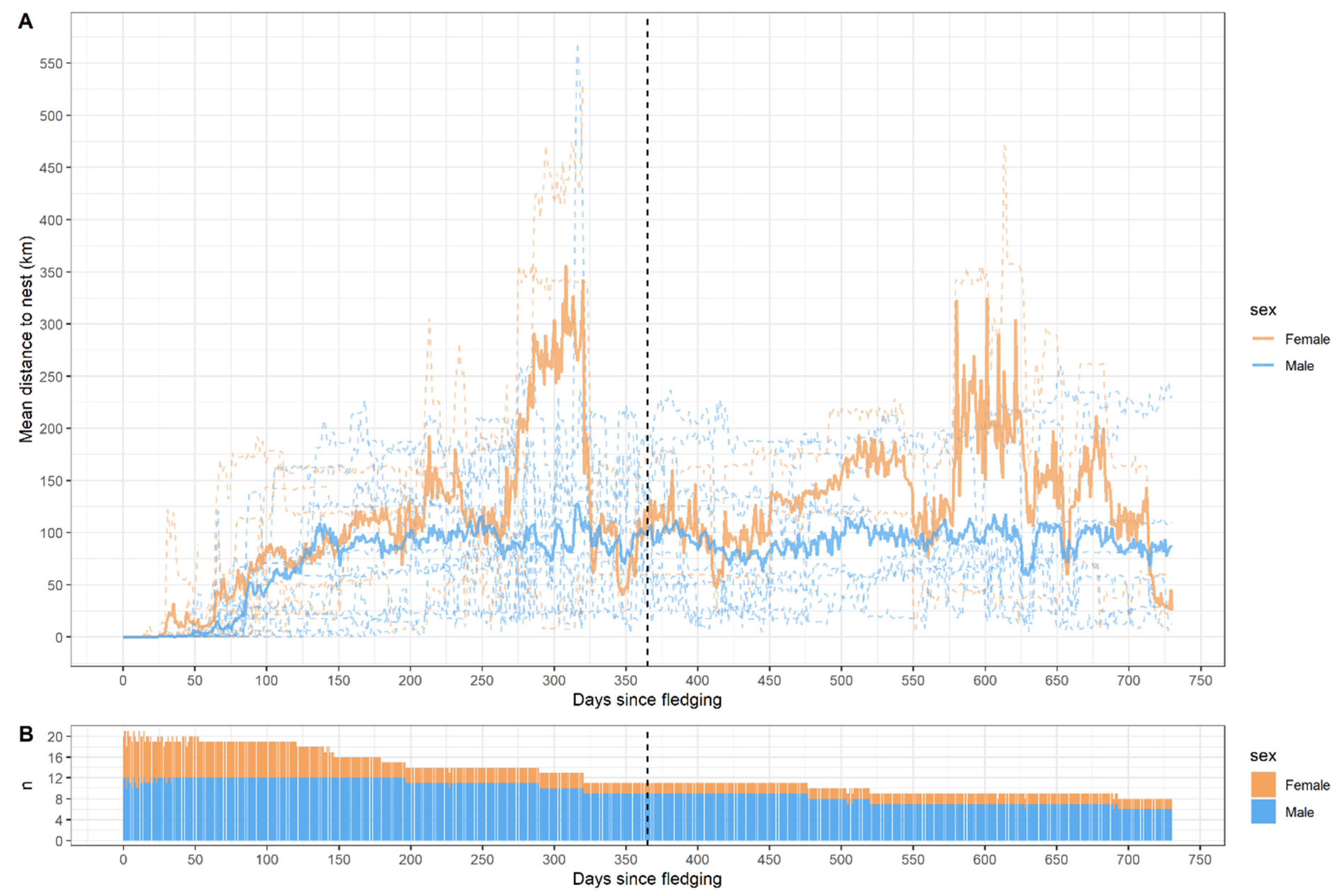
3.6. Distance to Nest
3.7. Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Mean Distance (km) | 95% Quartile Distance (km) |
|---|---|---|
| Evensø2017m1 (Gunhild) | 0.0053 | 0.0130 |
| Evensø2017m2 (Elna) | 0.0055 | 0.0140 |
| Vilsted2020f (Tove) | 0.0036 | 0.0090 |
| Hyllekrog2020f (Kim) | 0.0063 | 0.0150 |
| Hostrup2018f1 (Viola) | 0.0049 | 0.0120 |
| Vilsted2022f (Lisbeth) | 0.0040 | 0.0120 |
| Kongelunden2021m (Mads) | 0.0030 | 0.0070 |
| Hostrup2018f3 (Malthe-Ludvig) | 0.0065 | 0.0160 |
| Skanderborg2018m1 (Svend) | 0.0067 | 0.0170 |
| Haderslev2018m (Frode) | 0.0081 | 0.0210 |
| Kongelunden2018m (August) | 0.0037 | 0.0100 |
| Skanderborg2018m2 (Eskild) | 0.0069 | 0.0185 |
| Skanderborg2018f (Estrid) | 0.0063 | 0.0160 |
| Vilsted2019f (Mette) | 0.0038 | 0.0090 |
| Hyllekrog2020m (Romme) | 0.0054 | 0.0130 |
| Hostrup2018f2 (Ditte) | 0.0051 | 0.0121 |
| Rye2020m1 (Martin) | 0.0083 | 0.0217 |
| Vintersborg2019f (Maria-Anna) | 0.0074 | 0.0180 |
| Vilsted2019m (Asvi) | 0.0032 | 0.0070 |
| Vilsted2022m (Mikkel) | 0.0031 | 0.0090 |
| Rye2020m2 (Steen) | 0.0130 | 0.0501 |
| Filsø2022m (Aage) | 0.0327 | 0.0970 |

| Name | Sex | Fledging Date | Age at Fledging (Days) | First Exploratory Flight | Age at First Exploratory Flight (Days) | Dispersal at 5 km | Age at 5 km Dispersal | Approx. Date Dead | Approx. Age Dead (Days) | Cause of Death |
|---|---|---|---|---|---|---|---|---|---|---|
| Evensø2017m1 | Male | 04/08/2017 | 89 | 03/09/2017 | 119 | 14/10/2017 | 160 | |||
| Evensø2017m2 | Male | 27/07/2017 | 76 | 23/08/2017 | 103 | 03/11/2017 | 175 | 09/02/2018 | 273 | Avian flu (H5N6) |
| Haderslev2018m | Male | 26/06/2018 | 79 | 15/08/2018 | 129 | 24/08/2018 | 138 | 17/10/2022 | 1653 | |
| Hostrup2018f1 | Female | 30/06/2018 | 79 | 26/07/2018 | 105 | 27/07/2018 | 106 | 22/08/2018 | 132 | Wind turbine |
| Hostrup2018f2 | Female | 26/06/2018 | 75 | 26/07/2018 | 105 | 03/09/2018 | 144 | 23/12/2018 | 255 | Unknown |
| Hostrup2018f3 | Female | 30/06/2018 | 75 | 02/08/2018 | 108 | 02/09/2018 | 139 | |||
| Kongelunden2018m | Male | 15/07/2018 | 77 | 09/08/2018 | 102 | 09/10/2018 | 163 | |||
| Skanderborg2018m1 | Male | 11/06/2018 | 66 | 02/08/2018 | 118 | 05/09/2018 | 152 | 19/04/2022 | 1474 | Wind turbine |
| Skanderborg2018m2 | Male | 11/06/2018 | 65 | 15/08/2018 | 130 | 23/08/2018 | 138 | |||
| Skanderborg2018f | Female | 15/06/2018 | 66 | 10/07/2018 | 91 | Unknown | ||||
| Vilsted2019f | Female | 10/07/2019 | 89 | 24/07/2019 | 103 | 24/08/2019 | 134 | |||
| Vilsted2019m | Male | 23/06/2019 | 69 | 24/07/2019 | 100 | 15/09/2019 | 153 | 31/05/2021 | 777 | Wind turbine |
| Vintersborg2019f | Female | 22/07/2019 | 86 | 13/08/2019 | 108 | 15/09/2019 | 141 | 16/12/2019 | 233 | Lead poisoning |
| Hyllekrog2020f | Female | 11/07/2020 | 86 | 28/07/2020 | 103 | 09/11/2020 | 207 | Avian flu (H5N8) | ||
| Hyllekrog2020m | Male | 02/07/2020 | 79 | 01/08/2020 | 109 | 28/01/2021 | 289 | |||
| Rye2020m1 | Male | 14/07/2020 | 81 | 04/09/2020 | 133 | 28/09/2020 | 157 | |||
| Rye2020m2 | Male | 11/07/2020 | 81 | 25/08/2020 | 126 | 17/09/2020 | 149 | |||
| Vilsted2020f | Female | 01/07/2020 | 72 | 30/07/2020 | 101 | 03/09/2020 | 136 | |||
| Kongelunden2021m | Male | 03/08/2021 | 99 | 03/08/2021 | 99 | Unknown/starvation | ||||
| Filsø2022m | Male | 16/07/2022 | 91 | 24/07/2022 | 99 | 11/10/2022 | 178 | |||
| Vilsted2022m | Male | 15/06/2022 | 77 | 22/07/2022 | 114 | 20/10/2022 | 204 | |||
| Vilsted2022f | Female | 15/06/2022 | 71 | 05/08/2022 | 122 | 17/09/2022 | 165 |
References
- del Hoyo, J.; Elliott, A.; Sargatal, J.; Cabot, J. Handbook of the Birds of the World: New World Vultures to Guineafowl; Lynx Edicions: Barcelona, Spain, 1994. [Google Scholar]
- Krone, O.; Stjernberg, T.; Kenntner, N.; Tataruch, F.; Koivusaari, J.; Nuuja, I. Mortality factors, helminth burden, and contaminant residues in white-tailed sea eagles (Haliaeetus albicilla) from Finland. Ambio 2006, 35, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Mee, A.; Breen, D.; Clarke, D.; Heardman, C.; Lyden, J.; McMahon, F.; O’Sullivan, P.; O’Toole, L. Reintroduction of White-tailed Eagles Haliaeetus albicilla to Ireland. Ir. Birds 2016, 10, 301–314. [Google Scholar]
- Whitfield, P.D.; Duffy, K.; McLeod, D.R.A.; Evans, R.J.; MacLennan, A.M.; Reid, R.; Sexton, D.; Wilson, J.D.; Douse, A. Juvenile Dispersal of White-Tailed Eagles in Western Scotland. J. Raptor Res. 2009, 43, 110–120. [Google Scholar] [CrossRef]
- Ehmsen, E.; Pedersen, L.; Meltofte, H.; Clausen, T.; Nyegaard, T. The occurrence and reestablishment of White-tailed Eagle and Golden Eagle as breeding birds in Denmark. Dansk Orn. Foren. Tidssk. 2011, 105, 139–150. [Google Scholar]
- Skelmose, K.; Larsen, O.F. Projekt Ørn—Årsrapport 2021; DOF BirdLife Danmark: Copenhagen, Denmark, 2021. [Google Scholar]
- Christensen, J.S.; Hansen, T.H.; Rasmussen, P.A.F.; Nyegaard, T.; Eskildsen, D.P.; Clausen, P.; Nielsen, R.D.; Bregnballe, T. Systematisk Oversigt over Danmarks Fugle 1800–2019; Dansk Ornitologisk Forening (DOF)/BirdLife Denmark: Copenhagen, Denmark, 2022. [Google Scholar]
- Skelmose, K.; Larsen, O.F. Projekt Ørn—Årsrapport 2023; DOF BirdLife Danmark: Copenhagen, Denmark, 2023. [Google Scholar]
- Bønløkke, J.; Rahbek, C.; Madsen, J.J.; Thorup, K.; Pedersen, K.T.; Bjerrum, M. Dansk Trækfugleatlas; Rhodos A/S & The Zoological Museum, University of Copenhagen: Copenhagen, Denmark, 2007. [Google Scholar]
- Balotari-Chiebao, F.; Villers, A.; Ijäs, A.; Ovaskainen, O.; Repka, S.; Laaksonen, T. Post-fledging movements of white-tailed eagles: Conservation implications for wind-energy development. Ambio 2016, 45, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Krone, O.; Scharnweber, C. Two white-tailed sea eagles (Haliaeetus albicilla) collide with wind generators in northern Germany. J. Raptor Res. 2003, 37, 174. [Google Scholar]
- Helander, B.; Krone, O.; Räikkönen, J.; Sundbom, M.; Ågren, E.; Bignert, A. Major lead exposure from hunting ammunition in eagles from Sweden. Sci. Total Environ. 2021, 795, 148799. [Google Scholar] [CrossRef]
- Isomursu, M.; Koivusaari, J.; Stjernberg, T.; Hirvelä-Koski, V.; Venäläinen, E.-R. Lead poisoning and other human-related factors cause significant mortality in white-tailed eagles. Ambio 2018, 47, 858–868. [Google Scholar] [CrossRef]
- Krone, O.; Globig, A.; Ulrich, R.; Harder, T.; Schinköthe, J.; Herrmann, C.; Gerst, S.; Conraths, F.J.; Beer, M. White-Tailed Sea Eagle (Haliaeetus albicilla) Die-Off Due to Infection with Highly Pathogenic Avian Influenza Virus, Subtype H5N8, in Germany. Viruses 2018, 10, 478. [Google Scholar] [CrossRef]
- Radović, A.; Mikuska, T. Population size, distribution and habitat selection of the white-tailed eagle Haliaeetus albicilla in the alluvial wetlands of Croatia. Biologia 2009, 64, 156–164. [Google Scholar] [CrossRef]
- Rymešová, D.; Raab, R.; Machálková, V.; Horal, D.; Dorňáková, D.; Rozsypalová, L.; Spakovszky, P.; Literák, I. First-year dispersal in white-tailed eagles Haliaeetus albicilla. Eur. J. Wildl. Res. 2021, 67, 71. [Google Scholar] [CrossRef]
- Tikkanen, H.; Balotari-Chiebao, F.; Laaksonen, T.; Pakanen, V.; Rytkönen, S. Habitat use of flying subadult White-tailed Eagles (Haliaeetus albicilla): Implications for land use and wind power plant planning. Ornis Fenn. 2018, 95, 137–150. [Google Scholar] [CrossRef]
- Engler, M.; Krone, O. Movement patterns of the White-tailed Sea Eagle (Haliaeetus albicilla): Post-fledging behaviour, natal dispersal onset and the role of the natal environment. Ibis 2022, 164, 188–201. [Google Scholar] [CrossRef]
- Krone, O.; Berger, A.; Schulte, R. Recording movement and activity pattern of a White-tailed Sea Eagle (Haliaeetus albicilla) by a GPS datalogger. J. Ornithol. 2009, 150, 273–280. [Google Scholar] [CrossRef]
- Krone, O.; Nadjafzadeh, M.; Berger, A. White-tailed Sea Eagles (Haliaeetus albicilla) defend small home ranges in north-east Germany throughout the year. J. Ornithol. 2013, 154, 827–835. [Google Scholar] [CrossRef]
- Nygård, T.; Bevanger, K.; Dahl, E.L.; Flagstad, Ø.; Follestad, A.; Hoel, P.L.; Reitan, O. A study of White-tailed Eagle Haliaeetus albicilla movements and mortality at a wind farm in Norway. In BOU Proceedings—Climate Change and Birds; British Ornithologists’ Union: Peterborough, UK, 2010. [Google Scholar]
- Bragin, E.A.; Poessel, S.A.; Lanzone, M.J.; Katzner, T.E. Post-fledging movements and habitat associations of White-tailed Sea Eagles (Haliaeetus albicilla) in Central Asia. Wilson J. Ornithol. 2018, 130, 784–788. [Google Scholar] [CrossRef]
- Prugnolle, F.; de Meeus, T. Inferring sex-biased dispersal from population genetic tools: A review. Heredity 2002, 88, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Nygård, T.; Kenward, R.E.; Einvik, K. Radio telemetry studies of dispersal and survival in juvenile white-tailed sea eagles Haliaeetus albicilla in Norway. In Raptors at Risk; World Working Group on Birds of Prey: Berlin, Germany, 2000; pp. 487–497. [Google Scholar]
- Tøttrup, A.P.; Larsen, J.C.; Nielsen, J.T.; Christophersen, H.; Thorup, K. Individual spatiotemporal histories of first year Golden Eagles (Aquila chrysaetos) in Denmark using GPS-tracking. Dan. Ornitol. Foren. Tidsskr. 2023, 117, 131–137. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 September 2022).
- Helander, B.; Hailer, F.; Vilà, C. Morphological and genetic sex identification of white-tailed eagle Haliaeetus albicilla nestlings. J. Ornithol. 2007, 148, 435–442. [Google Scholar] [CrossRef]
- White, G.C.; Burnham, K.P. Program MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46 (Suppl. S1), S120–S139. [Google Scholar] [CrossRef]
- Evans, R.J.; Wilson, J.D.; Amar, A.; Douse, A.; Maclennan, A.; Ratcliffe, N.; Whitfield, P.D. Growth and demography of a re-introduced population of White-tailed Eagles Haliaeetus albicilla. Ibis 2009, 151, 244–254. [Google Scholar] [CrossRef]
- Krüger, O. The return of the white-tailed eagle (Haliaeetus albicilla) to northern Germany: Modelling the past to predict the future. Biol. Conserv. 2010, 143, 710–721. [Google Scholar] [CrossRef]
- Krone, O.; Kenntner, N.; Trinogga, A.; Nadjafzadeh, M.; Scholz, F.; Sulawa, J.; Totschek, K.; Schuck-Wersig, P.; Zieschank, R. Lead poisoning in White-tailed Sea Eagles: Causes and approaches to solutions in Germany. In Ingestion of Lead from Spent Ammunition: Implications for Wildlife and Humans; Watson, R.T., Fuller, M., Pokras, M., Hunt, W.G., Eds.; The Peregrine Fund: Boise, ID, USA, 2009. [Google Scholar]
- Nadjafzadeh, M.; Hofer, H.; Krone, O. The link between feeding ecology and lead poisoning in white-tailed eagles. J. Wildl. Manag. 2012, 77, 48–57. [Google Scholar] [CrossRef]
- Mirski, P.; Anderwald, D.; Pieczyński, P.; Zawadzka, D. Znazenie Miejsca Gniazdowego I Przebieg Usamodzielniania Sie Mlmodych Bielików Z Wigiweskiego Parku Narodowego. Studia i Materiały CEPL w Rogowie R. 20. Zeszyt 2018, 54, 134–144. [Google Scholar]





| Name | Popular Name | Sex | Location | Clutch Size | Hatching Date | Tagging Date | Age at Tagging (Days) |
|---|---|---|---|---|---|---|---|
| Evensø2017m1 | Gunhild | Male | Evensø, Præstø | 2 | 07/05/2017 | 03/07/2017 | 57 |
| Evensø2017m2 | Elna | Male | Evensø, Præstø | 2 | 12/05/2017 | 03/07/2017 | 52 |
| Haderslev2018m | Frode | Male | Haderslev | 1 | 08/04/2018 | 07/06/2018 | 60 |
| Hostrup2018f1 | Viola | Female | Hostrup Sø | 3 | 12/04/2018 | 01/06/2018 | 50 |
| Hostrup2018f2 | Ditte | Female | Hostrup Sø | 3 | 12/04/2018 | 07/06/2018 | 56 |
| Hostrup2018f3 | Malthe-Ludvig | Female | Hostrup Sø | 3 | 16/04/2018 | 07/06/2018 | 52 |
| Kongelunden2018m | August | Male | Kongelunden | 1 | 29/04/2018 | 19/06/2018 | 51 |
| Skanderborg2018m1 | Svend | Male | Skanderborg | 3 | 06/04/2018 | 01/06/2018 | 56 |
| Skanderborg2018m2 | Eskild | Male | Skanderborg | 3 | 07/04/2018 | 01/06/2018 | 55 |
| Skanderborg2018f | Estrid | Female | Skanderborg | 3 | 10/04/2018 | 01/06/2018 | 52 |
| Vilsted2019f | Mette | Female | Vilsted Sø | 2 | 12/04/2019 | 12/06/2019 | 61 |
| Vilsted2019m | Asvi | Male | Vilsted Sø | 2 | 15/04/2019 | 12/06/2019 | 58 |
| Vintersborg2019f | Maria-Anna | Female | Vintersborg Skov | 1 | 27/04/2019 | 17/06/2019 | 51 |
| Hyllekrog2020f | Kim | Female | Hyllekrog | 2 | 16/04/2020 | 09/06/2020 | 54 |
| Hyllekrog2020m | Romme | Male | Hyllekrog | 2 | 14/04/2020 | 09/06/2020 | 56 |
| Rye2020m1 | Martin | Male | Rye Sønderskov | 2 | 24/04/2020 | 17/06/2020 | 54 |
| Rye2020m2 | Steen | Male | Rye Sønderskov | 2 | 21/04/2020 | 17/06/2020 | 57 |
| Vilsted2020f | Tove | Female | Vilsted Sø | 1 | 20/04/2020 | 17/06/2020 | 58 |
| Kongelunden2021m | Mads | Male | Kongelunden | 1 | 26/04/2021 | 22/06/2021 | 57 |
| Filsø2022m | Aage | Male | Filsø | 1 | 16/04/2022 | 15/06/2022 | 60 |
| Vilsted2022m | Mikkel | Male | Vilsted Sø | 2 | 30/03/2022 | 29/05/2022 | 60 |
| Vilsted2022f | Lisbeth | Female | Vilsted Sø | 2 | 05/04/2022 | 29/05/2022 | 54 |
| Year of Life | 1st | 2nd | 3rd | 4th | 5th | 6th |
|---|---|---|---|---|---|---|
| Dead before end of year | ||||||
| Male | 2 | 0 | 1 | 0 | 1 | 0 |
| Female | 5 | 0 | 0 | 0 | 0 | 0 |
| Total | 7 | 0 | 1 | 0 | 1 | 0 |
| Alive at end of year | ||||||
| Male | 11 | 8 | 7 | 3 | 1 | 1 |
| Female | 3 | 2 | 2 | 1 | 1 | 0 |
| Total | 14 | 10 | 9 | 4 | 2 | 1 |
| Known fate at end of age category | ||||||
| Male | 13 | 8 | 8 | 3 | 2 | 1 |
| Female | 8 | 2 | 2 | 1 | 1 | 0 |
| Total | 21 | 10 | 10 | 4 | 3 | 1 |
| Status uncertain at end of year | ||||||
| Male | 0 | 3 | 3 | 7 | 8 | 8 |
| Female | 1 | 2 | 2 | 3 | 3 | 4 |
| Total | 1 | 5 | 5 | 10 | 11 | 12 |
| Survival rate | ||||||
| Male | 0.85 | 1.00 | 0.88 | 1.00 | 0.50 | 1.00 |
| Female | 0.38 | 1.00 | 1.00 | |||
| Sexes combined | 0.67 | 1.00 | 0.90 | 1.00 | 0.67 | 1.00 |
| Model | AICc | Delta AICc | AICc Weights | Model Likelihood | Num. Par | Deviance | −2log(L) |
|---|---|---|---|---|---|---|---|
| {s(sex*age1y,age2y+)} | 85.683 | 0.000 | 0.513 | 1.000 | 3 | 79.636 | 79.64 |
| {s(sex*age2m1y,age2y+)} | 88.419 | 2.736 | 0.131 | 0.255 | 5 | 78.301 | 78.30 |
| {s(age1y2y+)} | 88.960 | 3.277 | 0.100 | 0.194 | 2 | 84.936 | 84.94 |
| {s(age2m1y2y+)} | 89.315 | 3.633 | 0.083 | 0.163 | 3 | 83.268 | 83.27 |
| {s(sex*age2m1y2y+)} | 89.432 | 3.749 | 0.079 | 0.153 | 6 | 77.267 | 77.27 |
| {s(sex*age2m,age1y2y+)} | 90.541 | 4.858 | 0.045 | 0.088 | 4 | 82.462 | 82.46 |
| {s(sex)} | 91.059 | 5.376 | 0.035 | 0.068 | 2 | 87.036 | 87.04 |
| {s(.)} | 92.765 | 7.082 | 0.015 | 0.029 | 1 | 90.757 | 90.76 |
| Sex | Min. Age | Max._Age | Mean Age | Std. Err. | N |
|---|---|---|---|---|---|
| Female | 101 | 122 | 106.9 | 2.3 | 8 |
| Male | 99 | 133 | 115.2 | 3.6 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eskildsen, D.P.; Ali, N.Y.; Larsen, J.C.; Thorup, K.; Skelmose, K.; Tøttrup, A.P. Survival, Nest Site Affiliation and Post-Fledging Movements of Danish White-Tailed Eagles (Haliaeetus albicilla). Diversity 2024, 16, 314. https://doi.org/10.3390/d16060314
Eskildsen DP, Ali NY, Larsen JC, Thorup K, Skelmose K, Tøttrup AP. Survival, Nest Site Affiliation and Post-Fledging Movements of Danish White-Tailed Eagles (Haliaeetus albicilla). Diversity. 2024; 16(6):314. https://doi.org/10.3390/d16060314
Chicago/Turabian StyleEskildsen, Daniel Palm, Nina Yasmin Ali, Jonas Colling Larsen, Kasper Thorup, Kim Skelmose, and Anders P. Tøttrup. 2024. "Survival, Nest Site Affiliation and Post-Fledging Movements of Danish White-Tailed Eagles (Haliaeetus albicilla)" Diversity 16, no. 6: 314. https://doi.org/10.3390/d16060314
APA StyleEskildsen, D. P., Ali, N. Y., Larsen, J. C., Thorup, K., Skelmose, K., & Tøttrup, A. P. (2024). Survival, Nest Site Affiliation and Post-Fledging Movements of Danish White-Tailed Eagles (Haliaeetus albicilla). Diversity, 16(6), 314. https://doi.org/10.3390/d16060314