1. Introduction
The white-tailed eagle (
Haliaeetus albicilla) (WTE from hereon) has a northern Palearctic distribution, from Southern Greenland through Europe and Asia to the pacific coast in the East. The largest populations are in Norway and Russia (more than 55% of the population) [
1]. Many populations of WTEs have experienced massive declines during the last century due to hunting, poisoning, pesticides, and habitat degradation [
1,
2]. From the second half of the twentieth century, the population started to increase mainly due to legal protection in most European countries, phasing out problematic pesticides and successful reintroduction to certain areas [
3,
4].
In Denmark, the WTE officially returned as a breeding bird in 1995, after being extinct for decades due to many of the abovementioned reasons [
5]. Since its return, Dansk Ornitologisk Forening/DOF BirdLife has been extensively monitoring the species through Projekt Ørn (The Eagle Project) with regard to the number of breeding pairs, the rate of breeding success, and more [
6].
Thanks to the monitoring, which is primarily carried out by volunteers, we have a solid estimate of the number and distribution of breeding pairs in Denmark. The population has experienced a strong increase reaching ~150 breeding pairs in 2023 [
7,
8]. The breeding WTEs in Denmark are distributed mainly in the southeastern parts of Denmark on the islands of Lolland, Falster, Zealand, and the southeast of Funen [
7]. The Danish breeding population is believed to originate primarily from Germany but also partially from Sweden [
8,
9].
Despite the great increase in population throughout Denmark and the rest of Europe, WTEs still face significant threats, such as collision with wind turbines [
10,
11], lead poisoning [
2,
12,
13], and avian flu, especially in younger individuals [
14]. According to a study from Finland, anthropogenic factors are the main reason of mortality, accounting for up to 60% of the cases [
13]. Information about the causes of death was therefore also an important part of the present study.
Since 2017, DOF BirdLife and the Natural History Museum of Denmark have monitored Danish WTEs with the use of GPS tracking technology to acquire detailed knowledge of their movements. Other studies of the species using GPS technology have been carried out in other European countries, including Croatia [
15], Czech Republic, Hungary, Austria [
16], Finland [
17], Germany [
18,
19,
20], Ireland [
3], and Norway [
21].
Like other breeding birds, WTEs can be sensitive to disturbances at their nest sites during the breeding season. The official recommendation from DOF BirdLife is currently a year-round protection zone around the nest site of at least a 100 m radius with no public access, and during the breeding period between 1 February and 31 July, the protection zone is increased to a radius of 300 m. To test the efficiency of the nest protection zones, we present a quantification of the monthly affiliation of the first-calendar-year GPS-tagged individuals to the 300 m zone.
In Denmark, WTEs are considered mostly resident, but the species is known to undertake long-distance movements in other parts of its distributional range, e.g., in Central Asia [
22]. We still lack detailed knowledge of natal dispersal movements in Denmark, even though it is a vital process affecting both the dynamics, size, and gene flow of a population [
23]. Dispersal is a complex subject, and many factors seem to affect dispersal in raptors, both internal and external such as resource availability, quality of the natal habitat, and sex, and this has been the focus of several studies in recent years [
4,
16,
18,
24].
Here, we present quantifications of movements mainly during the emigration phase in the first few years after fledging, including (1) geographical distribution during the first and second calendar years, (2) age at first exploratory flight, and (3) dispersal age at different distance intervals. In general, WTEs are known to exhibit female-biased dispersal, where female birds are more prone to disperse and males either stay or return to their natal area [
23]. We therefore performed the analyses separately for both sexes.
2. Materials and Methods
2.1. Tagging and Transmitters
Between 2017 and 2022, 22 WTE nestlings were tagged at 10 different locations in Denmark (
Table 1). We chose the nests based on their accessibility, the degree of local-volunteer monitoring effort, as well as geographic considerations with the aim to achieve a broad coverage of the Danish WTE population. All nest sites in this study were in Jutland or on islands of Zealand and Lolland, Denmark.
We used solar-powered GPS/GSM-transmitters (CTT-1080-BT3 Series 3rd Gen from Cellular Tracking Technologies, Cape May, NJ, USA). All GPS transmitters had labels printed on the side with the study group’s contact information. The transmitters (~73 g) were attached in a backpack-style configuration with a Teflon ribbon body harness (~8 g) following Tøttrup et al. [
25]. The harnesses were fitted loosely on the eagles to allow subsequent body mass growth after tagging. Total weight of transmitter and harness corresponded to 1.4–2.3% of the weight of the nestlings at tagging. Nestlings were transported to the ground from the nests for GPS-tagging, and at the same time, they were measured, weighed, and ringed.
2.2. Data Filtering and Sampling
The rate and times of day at which GPS fixes (hereafter duty cycle) of the GPS tags were collected and transmitted varied throughout the project. The most common duty cycle included collecting positions every 30 min from one hour before astronomical dawn until one hour after astronomical dusk with data transmitted three times every week throughout the year. Early in the project during summertime, the tags were set to collect positions every 15 min with data transfer daily. Thirteen tags were set to produce so-called night bursts at midnight, in which roughly five positions were obtained in a few seconds. Night burst data were included in analyses.
In total 598,868 positions were obtained from tagged individuals between 3 July 2017 and 30 April 2022. In order to remove imprecise positions, the following technical criteria were used to filter the dataset: the horizontal dilution of precision must be less than 2 (HDOP < 2) (number of satellites used by tag for calculating position more than three (nsats > 3), the latitude higher than 1, and the longitude higher than 1. This left 337,450 filtered positions, of which 321,234 were from fledged individuals and 16,216 were from nestlings.
To avoid bias arising from the unequal number of obtained positions between individuals, 100 positions were randomly chosen from each individual for each month before analysis of nest zone affiliation and geographic distribution during 1st and 2nd calendar years. Only months with at least 100 positions available per individual were used for analyses.
2.3. Precision of Location Data
In order to estimate the accuracy of the filtered position data, we analyzed positions obtained between tagging date +1 day and fledging date −1 day. After applying the mentioned filter, each nest coordinate set (hereafter nest position) was calculated as the mean of the coordinates of observations for each individual. The distance between each position and the nest position for this and all subsequent analyses was calculated via the function
distGeo from the
geosphere package (v.1.5-14: Hijmans et al. 2021) in Ref. [
26].
The mean distance between positions and the corresponding nest position was 6.2 m, while 95% of the observations were less than 16.0 m from the nest position. The least precise position data were found for the individual Filsø2022m, which had a mean distance to the nest position of 30.3 m, with 95% of the positions less than 97.2 m from the nest position. Most individuals, however, had considerably better precision—the best one being Kongelunden2021m, whose mean distance to the nest position was 3.0 m and 95% of the positions less than 7.0 m from the nest position. See
Appendix A,
Table A1 for individual precision estimates.
2.4. Ageing and Sexing
In most cases, eagles were aged during tagging using wing length and body mass measurements. Two individuals, Hyllekrog2020f and Hyllekrog2020m, hatched in a nest was live-streamed via
www.ørnetv.dk, and thus their exact hatching date was known: 14 April 2020 and 16 April 2020.
Sexing was conducted using measures of tarsus as described by Helander et al. [
27]. Tarsus measurements from two individuals (Evensø2017m1 and Evensø2017m2) were unreliable due to faulty measuring equipment at the day of tagging. Evensø2017m1 though, is likely a male, based on visual observations from the field, including observations with a larger mate during two breeding attempts. Evensø2017m1 is therefore treated as a male in the analyses. Evensø2017m2 was sexed as male during autopsy. In total, 13 of the tagged individuals were males and 9 were females.
2.5. Fledging
Fledging date was defined as the first date on which several positions with hdop < 2 were well away from the nest, thus indicating that the individual had in fact fledged from the nest to neighboring trees or to the ground. This was determined visually by inspection of the obtained positions during the first weeks after tagging.
2.6. Survival
Survival was calculated as the proportion of individuals with known fate surviving past the estimated hatching date each subsequent year. Individuals with lost tags, tag malfunction, or not reaching the age for which survival was calculated were excluded from the calculation of survival rate but included in the category “status uncertain”.
Two individuals, Hostrup2018f1 and Kongelunden2021m, had to be taken into care during the project period. For survival and movement analyses, these were treated as dead from the day they were taken into care. One individual, Kongelunden2018m, has been photographed and observed several times in Denmark and Sweden after tag malfunction, thus allowing for inclusion in survival calculations after tag malfunction. For survival analyses, the studied period was extended to include 7 May 2023 to allow for the calculation of survival for the individual Evensø2017m1 in its 6th year of life.
We fitted known-fate monthly survival models in the MARK program [
28], including differences between sexes and separating survival into the first two months of life, first year of life, and after the first year of life.
2.7. Nest Zone Affiliation
We quantified nest zone affiliation in the age category “1st calendar year” by calculating the monthly proportion of sampled positions separately for both sexes from individuals that were (1) nestlings, (2) inside the recommended protection zone of 300 m, and (3) outside the recommended protection zone. One individual, Kongelunden2021m, never fledged and was therefore excluded from this analysis.
2.8. First Exploratory Flight
Age of first exploratory flight was defined as the age of a given individual on the date of the first position where the distance to the nest was greater than 2.0 km. We tested for difference between males and females using a two-tailed t-test.
2.9. Dispersal
Dispersal age was calculated at five distances for each individual: 0.3 km, 5 km, 25 km, 50 km, and 100 km. An individual was regarded as dispersed at each distance on the first 10 consecutive dates where all positions were further than the specified distance from the nest position, as performed by Balotari-Chiebao et al. [
10]. If not all 10 days during the investigated period contained positions, the individual was assumed to have been further from the nest than the investigated distance during the missing dates. We tested for difference between males and females using a two-tailed
t-test.
2.10. Distribution
Spatial distribution during the 1st and 2nd calendar years was quantified by counting positions in a 10 × 10 km grid covering Denmark, Sweden, and Germany. Positions from later malfunctioning tags and individuals that died during the corresponding age category were included. Individuals that had not yet fully reached their 1st or 2nd calendar year were included with the data available.
3. Results
3.1. Fledging
The tagged individuals (N = 22) hatched between 30 March and 12 May with a mean hatching date of ~17 April. Fledging happened between 11 June and 4 August, with a mean fledging date of ~3 July (N = 21). Mean age at fledging was ~78 days but varied considerably (range: 65–91 days) with no clear difference between males and females (
Figure 1). Fledging dates can be seen in
Table A2.
3.2. Survival
Survival was lowest during the first year of life where 14 of 21 (~67%) individuals with known fate survived (
Table 2). In total, three of eight females (~38%) and 11 of 13 males survived (~85%) their first year of life. All 10 individuals with known fate at the end of their second year survived. One male died during its third year and one male died during its fifth year.
Survival analyses showed considerable support for a model with differences in monthly survival between sexes in the first year and equal survival between sexes in later years (ΔAICc = 2.7;
Table 3). In this model, first-year monthly female survival (0.917, confidence interval = [0.815, 0.965]) was lower than male survival (0.986, CI = [0.945, 0.996]) and higher and equal thereafter (0.994, CI = [0.975, 0.998]). Survival during the first two months was generally lower but the difference was not supported when sex differences in the first year were included.
In the period of the study (2017–2022), a total of nine eagles died (five females, four males). The cause of death was determined in six cases: two cases of avian influenza (Evensø2017m2, avian flu type H5N6 and Hyllekrog2020f, avian flu type H5N8), three died from collisions with wind turbines (Hostrup2018f, Skanderborg2018m1, and Vilsted2019m), and one died from plausible lead poisoning in Northern Germany (Vintersborg2019f).
One individual, Skanderborg2018f, died close to the nest roughly 25 days after fledging. The corpse was retrieved approximately one week after the eagle died, in a state that did not allow autopsy. One individual, Kongelunden2021m, was shortly after fledging found near the nest site unable to fly with most primaries in its right wing moulting. Due to the nest being situated in a popular public area, the eagle was brought to a caring facility, where it was cared for until being able to fly. Upon its release near the nest, the eagle lived for ~20 days before again being taken into care. Unfortunately, it died at the caring facility, likely because of starvation during its prior release.
One individual, Evensø2017m1, attempted breeding in 2021, 2022, and 2023 having found a mate and begun nest building, potentially even incubating. Breeding attempts were unsuccessful potentially due to human disturbance in the nest area.
Five GPS tags ceased to transmit data during the project. One of the malfunctioned tags belonged to an individual (Kongelunden2018m) who has later had multiple sightings in field, with the latest in January 2022. The status of the other four individuals with inactive tags is uncertain, but it is likely that some have died since no confirmed sightings have been made after tag malfunction. One individual, Haderslev2018m, lost its transmitter in October of 2022.
3.3. Nest Zone Affiliation
Tagged individuals spent considerable time in the 300 m nest zone outside the recommended protection period between 1 February and 31 July (
Figure 2). In August, 20 out of 21 individuals still used the nest or nest zone for a combined ~46% of the sampled positions. One individual had not yet fledged from the nest by the beginning of August. During September, ~15% of the sampled positions came from individuals inside the nest zone.
We found strong support for an effect on the likelihood of staying close to the 300 m nest zone of an interaction between sex and month with males being more likely to stay near the nest in August to October, after which neither sex was likely to be found close to the nest (ΔAICc = 53.8 between model with and without interaction). For males, 18% of the sampled positions were inside the zone during September. In comparison, 8% of the sampled positions for females were inside the zone during the same month. Both males and females had only rather small proportions of their positions inside the protection zone during October–December (~5% or less).
3.4. First Exploratory Flight
Twenty individuals made exploratory flights before dispersal. Mean age of first exploratory flight for both sexes combined was ~112 days, corresponding to ~6 August, ranging from 99 to 133 days of age (
Table 4). Males were on average ~8 days older than females when undertaking their first exploratory flight, although means were not significantly different (t = −1.93, df = 17.3,
p = 0.07).
3.5. Dispersal
Mean dispersal age was ~153 days, corresponding to ~16 September, at 0.3 km (
n = 20, range: 104–251 days); 159 days, corresponding to ~22 September, at 5 km (
n = 19, range: 106–289 days); ~177 days at 25 km (
n = 17, range: 110–366 days); ~193 days at 50 km (
n = 15, range: 122–366 days); and ~238 days at 100 km (
n = 12, range: 140–401 days). On average, females were younger than males at all five dispersal distances (
Figure 3), although only significantly older at 5 km distance with a difference of 33.5 days (t = −2.46, df = 16,
p = 0.026) and at 100 km distance with a difference of ~92 days (t = −2.45, df = 10,
p = 0.034).
3.6. Distance to Nest
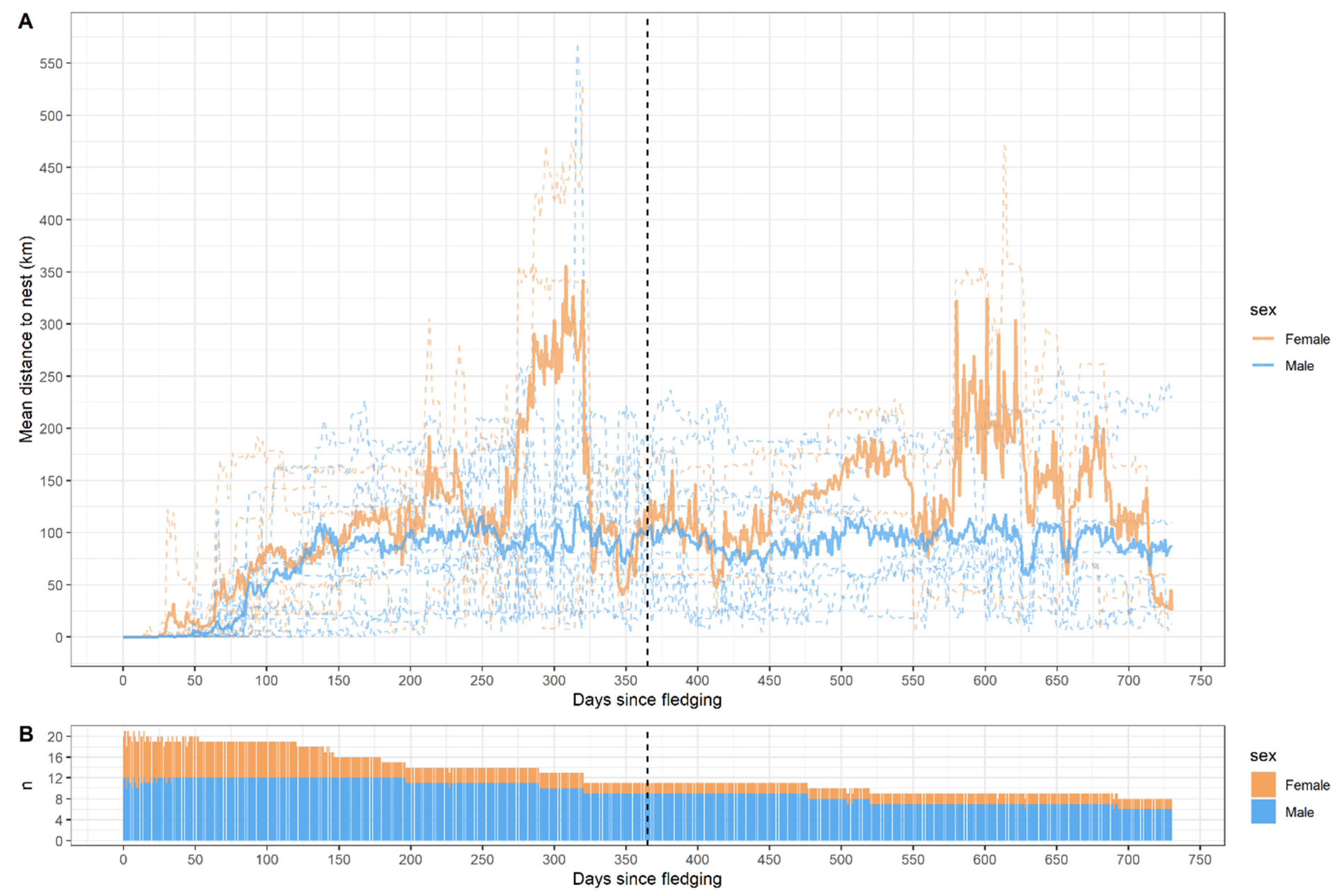
Mean distance to the nest increased gradually for both sexes during the first ~125 days after fledging (
Figure 4A). Distance to the nest site during the first two years varied greatly among individuals, with females on average being further away from the nest during the first ~125 days after fledging. Both sexes tended to remain within 250 km of the nest, but three individuals went on longer flights, with one female spending ~50 days more than 330 km away from the nest and another female flying northeast, reaching Lapland in Finland, Russia, and Norway, as far as ~1750 km from the nest medio May 2023. The average distance to the nest at day 365 since fledging was ~100 km for both sexes, but the sample size of females at this point was only 2 (
Figure 4B).
3.7. Distribution
Filtered positions from all individuals can be seen in
Figure 5. Individual maps from each fledged individual can be seen in
Appendix A,
Figure A1.
Figure 6 shows the calculated percentage of all sampled positions in a 10 × 10 km square grid of Denmark, Sweden, and Germany in the age categories of first and second calendar years.
4. Discussion
We found the average age of fledging to be ~78 days with no clear difference between males and females. This is similar to the findings of Balotari-Chiebao et al. [
10] (79 days), but later than those of Engler and Krone [
18] (72 days). Similar to our study, Engler and Krone [
18] found no difference between males and females in fledging age.
We found survival rate to be lower in the first year of life in comparison with later ages, although with small sample sizes later than year two and a large proportion of the tagged individuals having unknown fates in later age categories. Our reported survival of ~67% in the first year of life is comparable to Evans et al. [
29] and Krüger et al. [
30], who report a first-year survival of ~74% based on mark-recapture models. Evans et al. [
29] report a slight decrease in survival during the second year, whereas Krüger et al. [
30] report an increase. All 10 individuals with known fate in our study survived their second year. Our results show that the first year of life is critical for juvenile WTEs in Denmark, and that individuals surviving past their first year of life are very likely to reach maturity.
The low female survival rate during the first year (~38%) indicates that females are more prone to dangers in the early stages of life than males. This could be due to females leaving the natal territory earlier, which is supported by an earlier first exploratory flight and dispersal ages. Dispersing early from natal sites could result in leaving while parent birds are still likely to feed or teach survival techniques to their offspring. This seemed particularly the case for the female Hostrup2018f1, who dispersed very early from the nest site, and was found malnourished (~25 days after dispersing at a 5 km distance from the nest) and had to be taken into care. Upon release at its natal site, Hostrup2018f1 again traveled far from the site and once again ended up in care. Upon its final release, it apparently managed to survive on its own but ended up dying from collision with a wind turbine in May of 2019.
Although survival rate in our project is slightly lower in the first year compared to findings from Evans et al. [
29], we find higher survival rates in years 2–4, suggesting an overall similar survival rate. In this comparison, it is worth noting differences in methods: GPS technology potentially allows for a more precise calculation of survival rate, and the results by Evans et al. [
29] were based on observational data. The same could be true in Nygard et al.’s work [
24], who report a “remarkably high” annual survival rate of radio-tagged Norwegian WTEs during the first two years of life (90–95%).
GPS technology proved viable as a means of determining cause of death for WTEs in Denmark. In most cases when an individual died, we were able to track down the transmitter shortly after an individual had died and identify the cause of death from autopsy. In order to be able to locate dead individuals, the transmitter must continue to connect to the GSM-network, but this was not always the case. Skanderborg2018m1, from whom we lost signal in Northern Germany in April 2022, was later found dead by coincidence at a beach <10 km from its last known position. It appeared to have been washed ashore and thus, it is likely that the transmitters are not well suited to locate WTEs that perish at sea. Autopsy later revealed that the individual had died from trauma, and investigation of the last GPS fixes revealed that the individual had died at a wind turbine and apparently transported to the Wadden Sea for disposal.
Skanderborg2018m2 and Vilsted2019f likely died without the transmitters being able to reveal their final positions, but this remains uncertain.
We found four out of six causes of death determined with certainty to be of unnatural causes: three died by collision with wind turbines, suggesting a signification contribution to mortality in Danish WTE population. In all three cases, it would have been highly unlikely that the individuals would have been found if they had not been fitted with a GPS transmitter because of poor public access and generally low human activity at the locations. This result could indicate a rather large unknown proportion of Danish WTEs dying from collisions with wind turbines. BirdLife Denmark is currently aware of ~30 WTEs (including the three individuals from this study) that have died from collisions with wind turbines since 2013 (BirdLife Denmark), but this number is likely a conservative estimate of the actual number of wind-turbine-related casualties of WTEs in Denmark.
One individual apparently died of lead poisoning in Northern Germany, most likely from feeding on carrion containing lead-bullet fragments, which has been known as a general issue in Germany [
2,
31,
32], Sweden [
12], and Finland [
13].
Two individuals died of avian flu, indicating that this too could be a frequent cause of death among WTEs in Denmark as is also the case in other European countries [
14].
Even though most Danish WTEs will have fledged by 31 July, our results show that the individuals have close affiliation with nest sites in succeeding months. The tagged individuals in this study still used nest zones roughly half of the sunlit hours in August and a considerable amount of time in September. Additionally, dispersal from the nest zone (at 0.3 km) happened with a mean date of ~16 September. Our findings are similar to findings by Mirski et al. [
33] and Engler and Krone [
18]. The latter reported frequent visits by juvenile German WTEs at nest sites in September.
During daytime in the post-fledging months, WTEs will frequently be fed by parent WTEs. In this period, human disturbances can potentially lead to premature dispersal potentially reducing survival chances. Our findings thus suggest an extension of the present cautionary period to include at least the month of August and preferably September as well if year-round protection in the 300 m zones at nest sites is not possible.
The tagged individuals in this study were generally philopatric in the sense that they tended to reside relatively near to where they hatched, as shown in
Figure 4 and
Figure 5. Some individuals went on longer journeys, e.g., Vilsted2022f traveled far northeast in April and May 2023, reaching at least 1750 km from the nest site. This trip constitutes the current distance record for a Danish GPS-tagged WTE—the previous record being a ringed WTE from western Jutland that was recorded ~20 years later north of Stockholm ~770 km from where it was ringed in Denmark [
9].
The mean distance to the nest during the first two years of life after fledging was quite similar between the two sexes with a majority of the individuals of both sexes being closer to the nest by less than 250 km in most of the study period. Overall, our reported mean distances to the nest during the first year of life are very similar to those reported by Engler and Krone [
18] based on German-satellite-tagged WTEs, but the Danish WTEs in this study reached the ~100 km mark much earlier at ~125 days after fledging for both sexes. Three individuals, two females and one male, went on longer flights but two of these returned within 250 km of the nest sites during their first two years of dispersal.
Juvenile WTEs are known to exhibit more extensive movements within the first two years after fledging [
4]. Whitfield et al. suggested, based on observation records of tagged eagles (patagial tags and color-rings), that males initially dispersed further than females, while in their second year, females tended to disperse further than males. They found mean maximum juvenile dispersal distances to be between ~25 km (males, year 1) and ~80 km (females, year 2) during the first two years after fledging. Our results indicate that Danish WTEs disperse somewhat further than this with mean male distance to the nest being ~100 km and female mean distance to the nest being slightly longer, but with low sample size in females in year two (
Figure 4).
The distribution of the individuals was markedly different during the first and second calendar years (
Figure 6). During their first calendar year, they spent most of the time in close vicinity of nest sites, although other areas of Denmark and Northern Germany were also visited. In the second calendar year, the distribution was markedly different with the majority of positions coming from areas covering well-known WTE areas such as the Wadden Sea, Vejlerne, and Ringkøbing Fjord. Some local high proportions of the sampled positions during the second calendar year (
Figure 6) can be attributed to a single or only a few individuals that were stationary for a long term at certain sites. This was especially the case for the shown concentrations in northwestern Zealand at Saltbæk Vig and western and southern Funen and certain islands in Lillebælt. The location Saltbæk Vig is known to house a large number of resting WTEs. GPS telemetry thus proved useful in providing indications as to which areas are likely to house large concentrations of WTEs, which has been more thoroughly investigated by Skelmose and Larsen [
6].
We found a mean age of dispersal at 5 km to be 159 days corresponding to ~22 September. This is considerably later than the median dispersal date of 30 Aug at 5 km reported by Rymešová et al. [
16] based on GPS-tagged WTEs from Czech Republic, Hungary, and Austria. In Finland, Balotari et al. [
10] found the mean dispersal age at 5 km to be 164, which is similar to our findings. Engler and Krone [
18] visually defined the onset date of dispersal for each individual WTE in their study, but report a similar mean age of ~148 days at dispersal.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}