
Palynological Record of the Aalenian–Bajocian Cooling Event from the Santanghu Basin, Northwest China

Abstract

1. Introduction
2. Geological Setting
3. Materials and Methods
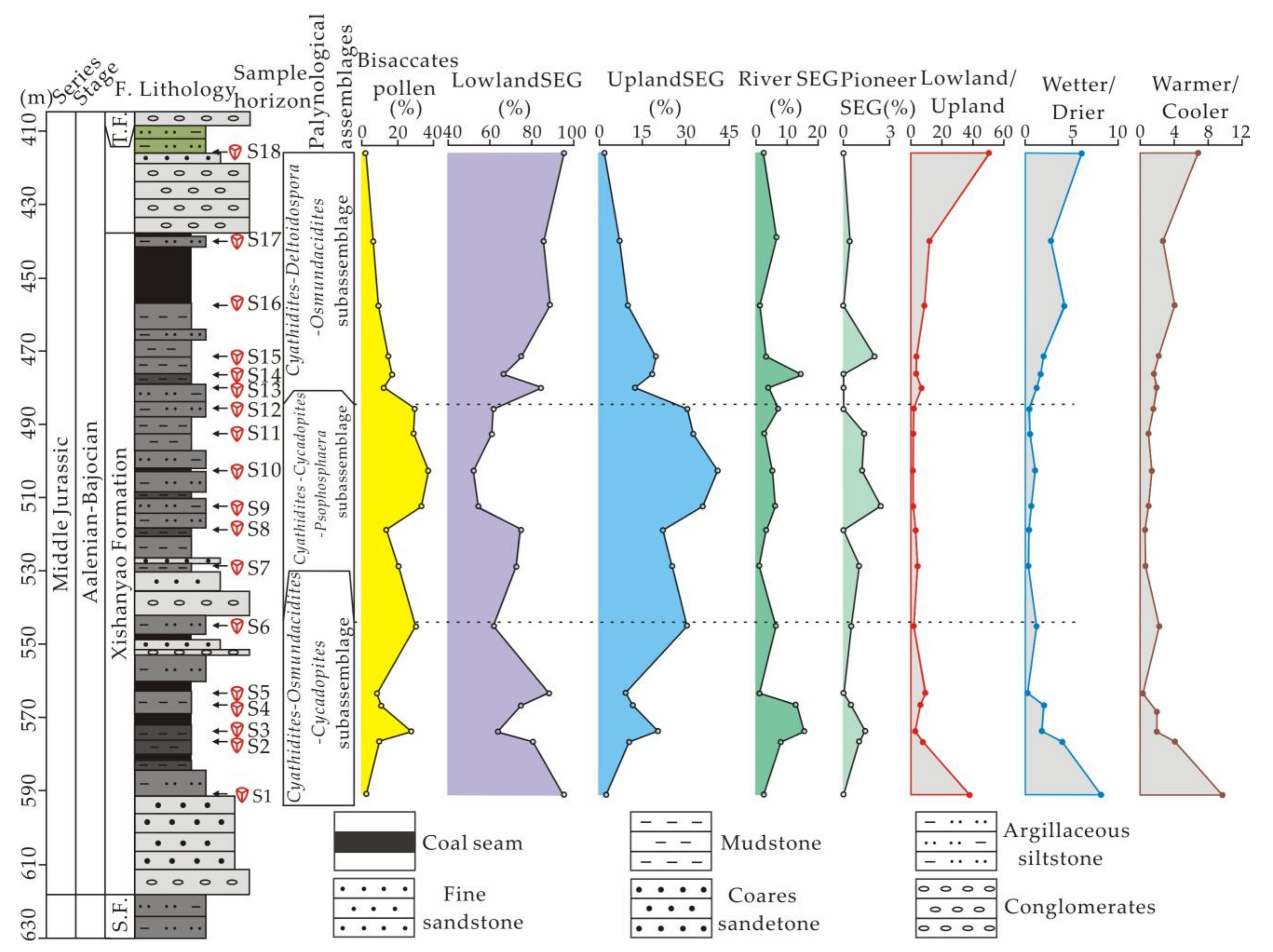
4. Results
4.1. Palynological Assemblage
4.1.1. Cyathidites–Osmundacidites–Cycadopites (COC) Subassemblage
4.1.2. Cyathidites–Cycadopites–Psophosphaera (CCP) Subassemblage
4.1.3. Cyathidites–Deltoidospora–Osmundacidites (CDO) Subassemblage
4.2. Sporomorph EcoGroup Analysis
5. Discussion
5.1. Geological Time
5.2. Paleovegetation Reconstruction
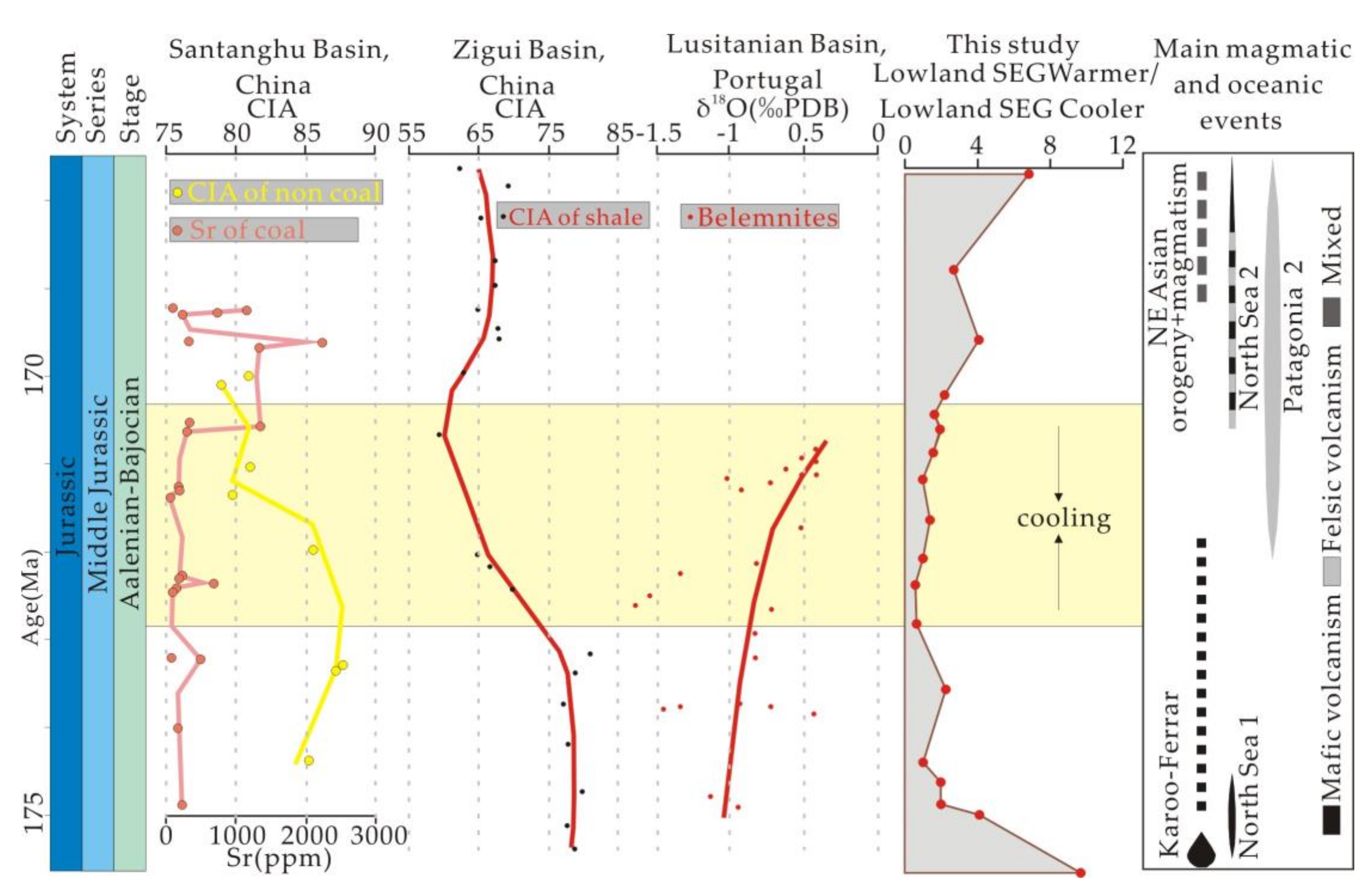
5.3. The Aalenian–Bajocian Cooling Event
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berner, R.A. GEOCARB II: A revised model of atmospheric CO2 over Phanerozoic time. Am. J. Sci. 1994, 294, 56–91. [Google Scholar] [CrossRef]
- Sellwood, B.W.; Valdes, P.J. Jurassic climates. Proc. Geol. Assoc. 2008, 119, 5–17. [Google Scholar] [CrossRef]
- Jenkyns, H.C. The early Toarcian (Jurassic) anoxic event: Stratigraphic, sedimentary, and geochemical evidence. Am. J. Sci. 1988, 288, 101–151. [Google Scholar] [CrossRef]
- Dera, G.; Brigaud, B.; Monna, F.; Laffont, R.; Pucéat, E.; Deconinck, J.F.; Pellenard, P.; Joachimski, M.M.; Durlet, C. Climatic ups and downs in a disturbed Jurassic world. Geology 2011, 39, 215–218. [Google Scholar] [CrossRef]
- Deng, S.H.; Lu, Y.Z.; Zhao, Y.; Fan, R.; Yang, X.J.; Li, X.; Sun, B.N. The Jurassic palaeoclimate regionalization and evolution of China. Earth Sci. Front. 2017, 24, 107–142. [Google Scholar]
- Dera, G.; Pellenard, P.; Niege, P.; Deconinck, J.F.; Pucéat, E.; Dommergues, J.L. Distribution of clay minerals in Early Jurassic peritethyan seas: Palaeoclimatic significance inferred from multiproxy comparisons. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 271, 39–51. [Google Scholar] [CrossRef]
- Korte, C.; Hesselbo, S.P. Shallow marine carbon and oxygen isotope and elemental records indicate icehouse-greenhouse cycles during the Early Jurassic. Paleoceanography 2011, 26, PA4219. [Google Scholar] [CrossRef]
- Zhang, L.W.; Yan, D.T.; Yang, S.G.; Li, B.Q.; Fu, H.J.; Wang, G.; Yang, X.R.; Zhang, W.L.; Zhang, J.F. Evolution of the Middle Jurassic paleoclimate: Sedimentary evidence from coal-bearing strata in the Santanghu Basin, NW China. J. Asian Earth Sci. 2023, 242, 105495. [Google Scholar] [CrossRef]
- Suchéras-Marx, B.; Giraud, F.; Fernandez, V.; Pittet, B.; Lécuyer, C.; Olivero, D.; Mattioli, E. Duration of the Early Bajocian and the associated δ13C positive excursion based on cyclostratigraphy. J. Geol. Soc. 2013, 170, 107–118. [Google Scholar] [CrossRef]
- Suchéras-Marx, B.; Suan, G.; Giraud, F.; Mattioli, E.; Khozyem, H.M.; Mazur, J.-C.; Fantasia, A.; Spangenberg, J.E.; Adatte, T. New insights into the early Bajocian (Middle Jurassic) carbon cycle perturbation. Sedimentologika 2023, 1, 1–18. [Google Scholar] [CrossRef]
- Price, G.D. Carbon-isotope stratigraphy and temperature change during the Early–Middle Jurassic (Toarcian–Aalenian), Raasay, Scotland, UK. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 285, 255–263. [Google Scholar] [CrossRef]
- Dai, X.; Du, Y.; Ziegler, M.; Wang, C.; Ma, Q.; Chai, R.; Guo, H. Middle Triassic to Late Jurassic climate change on the northern margin of the South China Plate: Insights from chemical weathering indices and clay mineralogy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 585, 110744. [Google Scholar] [CrossRef]
- Sun, L.X.; Zhang, Y.; Zhang, T.F.; Cheng, Y.H.; Li, Y.F.; Ma, H.L.; Yang, C.; Guo, J.C.; Lu, C.; Zhou, X.G. Jurassic sporopollen of Yan’an formation and Zhiluo formation in the northeastern Ordos basin, Inner Mongolia, and its paleoclimatic significance. Earth Sci. Front. 2017, 24, 32–51. [Google Scholar]
- Zhang, H.Q.; Wang, T.D.; Yan, C.F.; Yuan, J.Y. The subdivision and correlation of the Jurassic system of the Minhe basin. J. Stratigr. 2007, 31, 62–67. [Google Scholar]
- Wang, Y.D.; Jiang, D.X.; Yang, H.Q.; Sun, F. Middle Jurassic palynoflora and its palaeoenvironmental implication in Turpan depression, Xinjiang. Acta Sedimentol. Sin. 1997, 15, 133–140. [Google Scholar]
- Liu, Z.S.; He, Z.S.; Dong, K.L. Palynological assemblage of an adrilling-section (basal part of the Kizilnur formation) from Yaha, Kuqa, Xinjiang, N W China. Acta Micropalaeontol. Sin. 1999, 16, 82–88. [Google Scholar]
- Wei, Y.; Yang, B.; Xia, H.D.; Deng, H.J. Paleovegetation and paleoclimate during Mid-Late Eocene in Fushun basin. Earth Sci. 2021, 46, 1848–1861. [Google Scholar]
- Guiot, J.; Pons, A.; de Beaulieu, L.; Reille, M. A 140000 year continental climate reconstruction from two European pollen records. Nature 1989, 338, 309–313. [Google Scholar] [CrossRef]
- Li, B.; Wen, X.F.; Zhao, B.; Qiu, H.T.; Wang, X.Q.; Pei, Z.C. Sporo-pollen assemblages and paleoclimate analysis since the Late Pleistocene sediments of Nanyang basin. Geol. Sci. Technol. Inf. 2015, 34, 49–56. [Google Scholar]
- Birks, H.J.B.; Birks, H.H.; Ammann, B. The fourth dimension of vegetation. Science 2016, 354, 412–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K. The characteristic of plate tectonic evolution and the evaluation of hydrocarbon prospect of Santanghu basin, Xinjiang. Xinjiang Pet. Geol. 1993, 14, 1–13. [Google Scholar]
- Pan, Y.S.; Huang, Z.L.; Li, T.J.; Guo, X.B.; Xu, X.F.; Chen, X. Environmental response to volcanic activity and its effect on organic matter enrichment in the Permian Lucaogou formation of the Malang Sag, Santanghu basin, Northwest China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 560, 110024. [Google Scholar] [CrossRef]
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Grimm, E.C. Tilia, TiliaGraph and TGView Software; Illinois State Museum: Springfield, IL, USA, 2001. [Google Scholar]
- Abbink, O.A.; Van Konijnenburg-Van Cittert, J.H.A.; Visscher, H. A sporomorph ecogroup model for the Northwest European Jurassic—Lower Cretaceousi: Concepts and framework. Neth. J. Geosci. Geol. Mijnb. 2004, 83, 17–31. [Google Scholar] [CrossRef]
- Li, L.Q.; Wang, Y.D.; Liu, Z.S.; Zhou, N.; Wang, Y. Late Triassic palaeoclimate and palaeoecosystem variations inferred by palynological record in the northeastern Sichuan Basin, China. Paläontol. Z. 2016, 90, 327–348. [Google Scholar] [CrossRef]
- Li, L.Q.; Wang, Y.D.; Vajda, V.; Liu, Z.S. Late Triassic ecosystem variations inferred by palynological records from Hechuan, southern Sichuan Basin, China. Geol. Mag. 2018, 155, 1793–1810. [Google Scholar] [CrossRef]
- Kershaw, P.; Wagstaff, B. The southern conifer family Araucariaceae: History, status, and value for paleoenvironmental reconstruction. Annu. Rev. Ecol. Syst. 2001, 32, 397–414. [Google Scholar] [CrossRef]
- Couper, R.A. British Mesozoic microspores and pollen grains. A systematic and stratigraphic study. Palaeontogr. Abt. B Palaeophytol. Palaeobot.-Palaeophytol. 1958, 103, 75–179. [Google Scholar]
- Huang, L.; Zhang, X.J.; Li, G.Z.; Ji, X.K.; Han, X.; Wang, Z.Y.; Liu, Z.Q.; Hou, Z.M.; Ai, Z.H. Sporopollen assemblage from the Middle Jurassic of the northeastern Ordos Basin, Inner Mongolia, and their paleoclimatic implications. Acta Geol. Sin. 2023, 97, 1390–1406. [Google Scholar]
- Liu, Z.S. Early and middle Jurassic sporo-pollen assemblages from the Shiguai coal-field of Baotou, Nei Monggol. Acta Palaeontol. Sin. 1982, 21, 371–381. [Google Scholar]
- Du, B.A.; Li, X.R.; Duan, W.H. Sporo-pollen assemblages from Yan’an and Zhiluo formation in Chongxin county, Gansu province. Acta Micropalaeontol. Sin. 1982, 21, 597–609. [Google Scholar]
- Huang, P. Early-middle Jurassic sporopollen assemblages from Dananhu coalfield of Thuha basin, Xinjiang and their stratigraphical significance. Acta Palaeontol. Sin. 1995, 34, 171–197. [Google Scholar]
- Wang, Y.D.; Jiang, D.X.; Yang, H.Q.; Sun, F. Middle Jurassic spore-pollen assemblages from Turpan-Shanshan area, Xinjiang. Acta Bot. Sin. 1998, 40, 969–976. [Google Scholar]
- Qian, L.J.; Wang, J.G. Peat-Forming Materials and Swampy Environment of Middle Jurassic Terrestrial Bituminite; China Coal Industry Publishing House: Beijing, China, 1994; pp. 1–139. [Google Scholar]
- Liu, Z.S.; Zhang, Z.H. Late Early Jurassic Fuxian Formation spore pollen assemblage and its significance, Fuxian county, Shaanxi. In International Symposium on Geological Sciences at Peking University; Seismological Press: Beijing, China, 1998; pp. 430–438. [Google Scholar]
- Xu, Y.L.; Zhang, W.P. Mesozoic Strata and Paleontologyinthe Shaanxi-Gansu-Ningxia Basin (Volume I); Geological Publishing House: Beijing, China, 1980; pp. 143–186. [Google Scholar]
- Liu, Z.S. Sporo-pollen assemblage from middle Jurassic Xishanyao formation of Shawan, Xinjiang, China. Acta Palaeontol. Sin. 1990, 29, 63–87. [Google Scholar]
- Santos, A.A.; Wang, X.; Qiang, F.Q.; Diez, J.B. First palynological data from the Jurassic South Xiangshan Formation (Nanjing area, China). Geobios 2018, 51, 559–570. [Google Scholar] [CrossRef]
- Huang, P.; Li, J.G. Sporopollen assemblages from the Xishanyao and Toutunhe formation at the Honggou Section of the Manasi River, Xinjiang and their stratigraphical significance. Acta Micropalaeontol. Sin. 2007, 24, 170–193. [Google Scholar]
- Ilyina, V.I. Subdivision and correlation of the marine and non-marine Jurassic sediments in Siberia based on palynological evidence. Rev. Palaeobot. Palynol. 1986, 48, 357–364. [Google Scholar] [CrossRef]
- Huang, P. Middle Jurassic sporopollen assemblages from Tangqiang-3 well of Santanghu basin, Xinjiang. Acta Micropalaeontol. Sin. 2002, 19, 178–192. [Google Scholar]
- Sun, G.; Miao, Y.Y.; Mosbrugger, V.; Ashraf, A.R. The Upper Triassic to Middle Jurassic strata and floras of the Junggar Basin, Xinjiang, Northwest China. Palaeobiodivers. Palaeoenviron. 2010, 90, 203–214. [Google Scholar] [CrossRef]
- Korte, C.; Hesselbo, S.P.; Ullmann, C.V.; Dietl, G.; Ruhl, M.; Schweigert, G.; Thibault, N. Jurassic climate mode governed by ocean gateway. Nat. Commun. 2015, 6, 10015. [Google Scholar] [CrossRef]
- Jones, C.E.; Jenkyns, H.C.; Coe, A.L.; Stephen, H.P. Strontium isotopic variations in Jurassic and Cretaceous seawater. Geochim. Cosmochim. Acta 1994, 58, 3061–3074. [Google Scholar] [CrossRef]
- Ferreira, J.; Mattioli, E.; Sucherás-Marx, B.; Giraud, F.; Duarte, L.V.; Pittet, B.; Suan, G.; Hassler, A.; Spangenberg, J.E. Western Tethys Early and Middle Jurassic calcareous nannofossil biostratigraphy. Earth-Sci. Rev. 2019, 197, 102908. [Google Scholar] [CrossRef]
- Bodin, S.; Mau, M.; Sadki, D.; Danisch, J.; Nutz, A.; Krencker, F.-N.; Kabiri, L. Transient and secular changes in global carbon cycling during the early Bajocian event: Evidence for Jurassic cool climate episodes. Glob. Planet. Change 2020, 194, 103287. [Google Scholar] [CrossRef]
- Shen, J.; Yu, J.X.; Chen, J.B.; Algeo, T.J.; Xu, G.Z.; Feng, Q.L.; Shi, X.; Planavsky, N.J.; Shu, W.C.; Xie, S.C. Mercury evidence of intense volcanic effects on land during the Permian-Triassic transition. Geology 2019, 47, 1117–1121. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, X.Z.; Yi, J.J.; Shi, W.X.; Sun, S.Y.; Sun, S.Y.; Su, G.F. Carbon-cycle perturbations and intensified continental chemical weathering linked to volcanism during the Jenkyns Event in the Ordos Basin. Geol. J. 2024, 59, 1298–1321. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Botanical Affinity | Sporomorph Genera | SEG | Ecological Remarks |
|---|---|---|---|
| Ferns (Cyatheaceae) | Cyathidites, Deltoidospora | Lowland | Wetter, warmer |
| Ferns (Marattiaceae) | Cyclogranisporites | ||
| Ferns (Pteridaceae) | Asseretospora | ||
| Ferns (Dipteridaceae/Matoniaceae) | Dictyophyllidites, Concavisporites | ||
| Ferns (Dipteridaceae) | Kyrtomisporis | ||
| Ferns (Dicksoniaceae) | Converrucosisporites, Undulatisporites, Leiotriletes | ||
| Ferns (Osmundaceae) | Osmundacidites, Conbaculatisporites, Todisporites | ||
| Ferns | Auritulinasporites | ||
| Horsetails | Laevigatosporites, Calamospora | ||
| Lycopsids/Horsetails/Marttiaceae | Punctatisporites | ||
| Conifers (Taxodiaceae) | Inaperturopollenites | Lowland | Wetter, cooler |
| Conifers (Araucariaceae) | Psophosphaera | Lowland | Drier, warmer |
| Conifers (Araucariaceae) | Araucariacites, Callialasporites | ||
| Gymnosperms | Granasporites | Lowland | Drier, cooler |
| Cycads/Ginkgophytes | Chasmatosporites, Cycadopites, Monosulcites | ||
| Conifers (Podocarpaceae) | Podocarpidites, Quadraeculina, | Upland | |
| Conifers (Pinaceae) | Cedripites, Piceaepollenites, Pinuspollenites, Piceites | ||
| Gymnosperms | Cordaitina | ||
| Conifers | Erlianpollis, Pityosporites, Pseudopinus, Pseudopicea, Protoconiferus, Caytonipollenites | ||
| Conifers (Pinaceae)/Pteridospermae | Alisporites | ||
| Bryophytes | Alsophilidites, Annulispora, Sphagnumsporites | River | |
| Lycopsids (Lycopodiaceae) | Lycopodiumsporites | ||
| Lycopsids (Selaginellaceae) | Lycopodiacidites, Kraeuselisporites | ||
| Lycopsids (Isoetales) | Aratrisporites | ||
| Lycopsids | Neoraistrickia, Densoisporites | ||
| Conifers (Taxodiaceae) | Cerebropollenites | Pioneer | |
| Spheripollenties, Concentrisporites | Not attributed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Zhang, X.; Li, W.; Sun, S.; Yi, J. Palynological Record of the Aalenian–Bajocian Cooling Event from the Santanghu Basin, Northwest China. Diversity 2024, 16, 369. https://doi.org/10.3390/d16070369
Yang B, Zhang X, Li W, Sun S, Yi J. Palynological Record of the Aalenian–Bajocian Cooling Event from the Santanghu Basin, Northwest China. Diversity. 2024; 16(7):369. https://doi.org/10.3390/d16070369
Chicago/Turabian StyleYang, Bing, Xinzhi Zhang, Weitong Li, Siyuan Sun, and Jinjun Yi. 2024. "Palynological Record of the Aalenian–Bajocian Cooling Event from the Santanghu Basin, Northwest China" Diversity 16, no. 7: 369. https://doi.org/10.3390/d16070369
APA StyleYang, B., Zhang, X., Li, W., Sun, S., & Yi, J. (2024). Palynological Record of the Aalenian–Bajocian Cooling Event from the Santanghu Basin, Northwest China. Diversity, 16(7), 369. https://doi.org/10.3390/d16070369