


Taxonomic and Functional Diversity of Reptiles in a Heterogeneous Landscape of Jalisco State, West-Central Mexico


 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
2.3.1. Alpha Taxonomic and Functional Diversity
2.3.2. Beta Taxonomic and Functional Diversity
2.3.3. Taxonomic and Functional Diversity Facing Environmental Variables
3. Results
3.1. Alpha Taxonomic and Functional Diversity
3.2. Beta Taxonomic and Functional Diversity
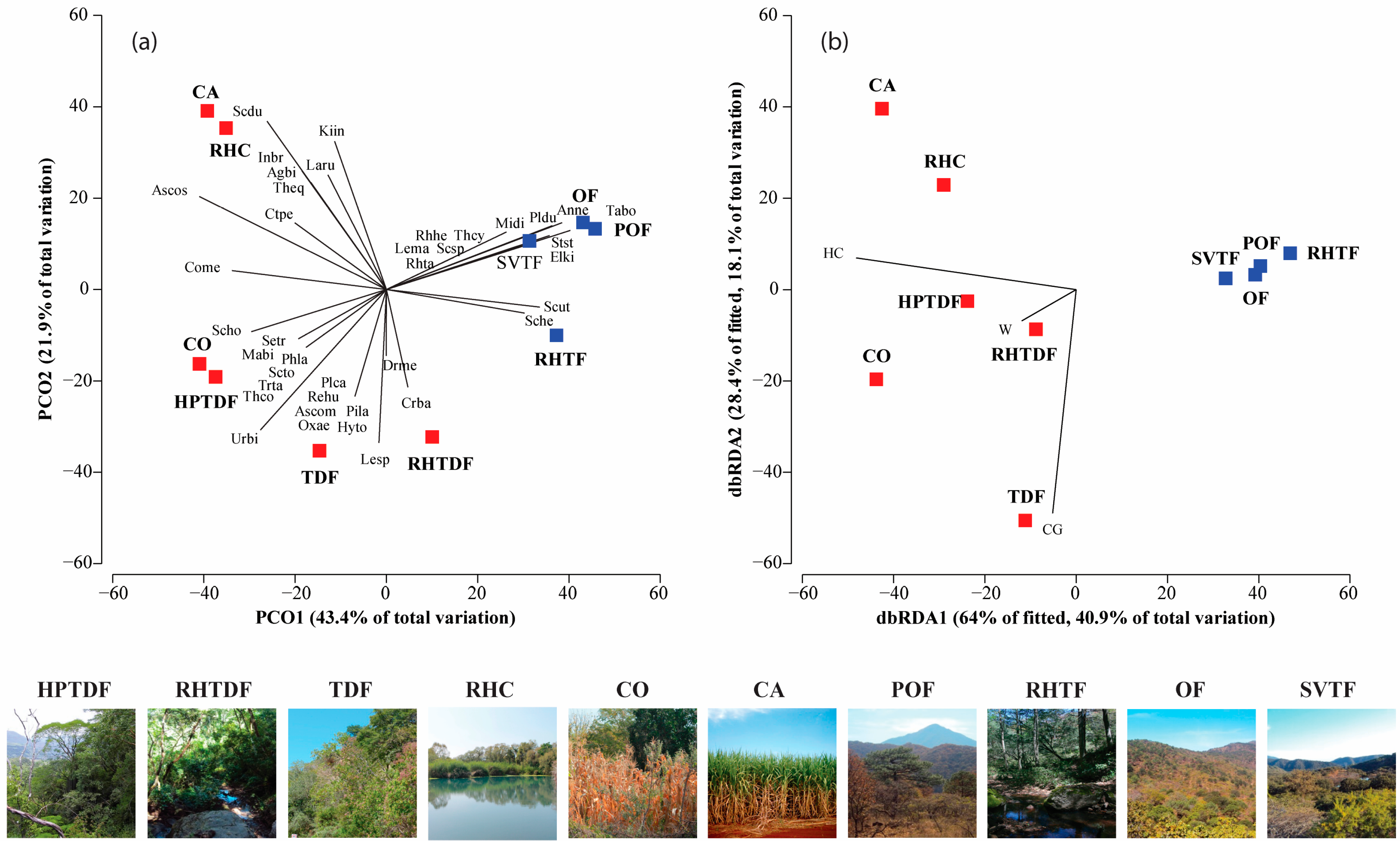
3.3. Reptile Diversity Facing Environmental Variables
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pincheira-Donoso, D.; Bauer, A.M.; Meiri, S.; Uetz, P. Global Taxonomic Diversity of Living Reptiles. PLoS ONE 2013, 8, e59741. [Google Scholar] [CrossRef]
- Valencia-Aguilar, A.; Cortés-Gómez, A.M.; Ruiz-Agudelo, C.A. Ecosystem services provided by amphibians and reptiles in Neotropical ecosystems. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2013, 9, 257–272. [Google Scholar] [CrossRef]
- Vitt, L.J.; Caldwell, J.P. Herpetology, Introductory Biology of Amphibians and Reptiles, 4th ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2014; p. 776. [Google Scholar]
- Nelson, E.C. Lizards on Aeonium lancerottense in Lanzarote, Canary Islands: A new example of pollination by reptiles? Bradleya 2010, 28, 15–18. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Aguilar, R.; Reyes, F.; Hošek, J. (Eds.) The Reptile Database. Available online: https://www.reptile-database.org (accessed on 5 May 2024).
- Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad (CONABIO). Biodiversidad Mexicana. Available online: http://www.biodiversidad.gob.mx (accessed on 5 May 2024).
- Flores-Villela, O.; García-Vázquez, U. Biodiversidad de reptiles en México. Rev. Mex. Biodivers. 2014, 85, 467–475. [Google Scholar] [CrossRef]
- Ramírez-Bautista, A.; Torres-Hernández, L.A.; Cruz-Elizalde, R.; Berriozabal-Islas, C.; Hernández-Salinas, U.; Wilson, L.D.; Johnson, J.D.; Porras, L.W.; Balderas-Valdivia, C.J.; González-Hernández, A.J.X.; et al. An updated list of the Mexican herpetofauna: With a summary of historical and contemporary studies. Zookeys 2023, 1166, 287–306. [Google Scholar] [CrossRef] [PubMed]
- García, A. Using ecological niche modeling to identify diversity hotspots for the herpetofauna of Pacific lowlands and adjacent interior valleys of Mexico. Biol. Conserv. 2006, 130, 25–46. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.M.; Flores-Villela, O.A. Áreas de Diversidad y Endemismo de la Herpetofauna Mexicana; Universidad Nacional Autónoma de México-CONABIO: Mexico City, México, 2006; p. 218. [Google Scholar]
- Baena, M.L.; Halfter, G. Extinción de especies. In Capital Natural de México, Volume I: Conocimiento Actual de la Biodiversidad; Sarukhán, J., Ed.; CONABIO: Mexico City, México, 2008; pp. 264–275. [Google Scholar]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- IUCN. World’s Reptiles Comprehensively Assessed-IUCN Red List. 2022. Available online: https://www.iucn.org/news/species/202204/worlds-reptiles-comprehensively-assessed-iucn-red-list (accessed on 10 June 2024).
- Cox, N.; Young, B.E.; Bowles, P.; Fernadez, M.; Marin, J.; Rapacciuolo, G.; Böhm, M.; Brooks, T.M.; Hedges, S.B.; Hilton-Taylor, C.; et al. A global reptile assessment highlights shared conservation needs of tetrapods. Nature 2022, 605, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Rincón-Aranguri, M.; Toro-Cardona, F.A.; Galeano, S.P.; Roa-Fuentes, L.; Urbina-Cardona, N. Functional diversity of snakes is explained by the landscape composition at multiple areas of influence. Ecol. Evol. 2023, 13, e10352. [Google Scholar] [CrossRef]
- Fisher, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Instituto de Información Estadística y Geografía del Estado de Jalisco (IIEG). Valles Diagnóstico de la Región. Available online: https://iieg.gob.mx/ns/wp-content/uploads/2023/08/Valles.pdf (accessed on 5 May 2024).
- Petchey, O.L.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Uribe, D.M.; Hoyos-Hoyos, J.M.; Isaacs-Cubides, P.; Corral-Gómez, N.; Urbina-Cardona, N. Classification and sensitivity of taxonomic and functional diversity indices of anurans in the Andean coffee cultural landscape. Ecol. Indic. 2022, 135, 108650. [Google Scholar] [CrossRef]
- Hernández-Salinas, U.; Cruz-Elizalde, R.; Ramírez-Bautista, A.; Wilson, L.D.; Berriozabal-Islas, C.; Johnson, J.D.; Mata-Silva, V. Taxonomic and functional diversity of the amphibian and reptile community of the state of Durango, Mexico. Community Ecol. 2023, 24, 229–242. [Google Scholar] [CrossRef]
- Villeger, S.; Ramos-Miranda, J.; Flores-Hernández, D.; Mouillot, D. Contrasting changes in taxonomic vs. functional diversity of tropical fish communities after habitat degradation. Ecol. Appl. 2010, 20, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Santos, X.; Cheylan, M. Taxonomic and functional response of a Mediterranean reptile assemblage to a repeated fire regime. Biol. Conserv. 2013, 168, 90–98. [Google Scholar] [CrossRef]
- Tsianou, M.A.; Lazarina, M.; Andrikou-Charitidou, A.; Michailidou, D.; Kallimanis, A.S. The Effect of Climate and Human Pressures on Functional Diversity and Species Richness Patterns of Amphibians, Reptiles and Mammals in Europe. Diversity 2021, 13, 275. [Google Scholar] [CrossRef]
- Berriozabal-Islas, C.; Ramírez-Bautista, A.; Cruz-Elizalde, R.; Hernández-Salinas, U. Modification of landscape as promoter of change in structure and taxonomic diversity of reptile’s communities: An example in tropical landscape in the central region of Mexico. Nat. Conserv. 2018, 28, 33–49. [Google Scholar] [CrossRef]
- López-Bedoya, P.A.; Cardona-Galvis, E.A.; Urbina-Cardona, J.N.; Edwards, F.A.; Edwards, D.P. Impacts of pastures and forestry plantations on herpetofauna: A global meta-analysis. J. Appl. Ecol. 2022, 59, 3038–3048. [Google Scholar] [CrossRef]
- Barnagaud, J.; Geniez, P.; Cheylan, M.; Crochet, P.A. Climate overrides the effects of land use on the functional composition and diversity of Mediterranean reptile assemblages. Divers. Distrib. 2021, 27, 50–64. [Google Scholar] [CrossRef]
- Berriozabal-Islas, C.; Badillo-Saldaña, L.M.; Ramírez-Bautista, A.; Moreno, C.M. Effects of hábitat disturbance on lizard functional diversity in a tropical dry forest of the Pacific Coast of Mexico. Trop. Conserv. Sci. 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Pollock, L.J.; O’Connor, L.M.J.; Mokany, K.; Rosauer, D.F.; Talluto, M.V.; Thuiller, W. Protecting biodiversity (in all its complexity): New models and methods. Trends Ecol. Evol. 2020, 35, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Sayer, J.; Sunderland, T.; Ghazoul, J.; Pfund, J.L.; Sheil, D.; Meijaard, E.; Venter, M.; Boedhihartono, A.K.; Day, M.; Garcia, C.; et al. Ten principles for a landscape approach to reconciling agriculture, conservation, and other competing land uses. Proc. Natl. Acad. Sci. USA 2013, 110, 8349–8356. [Google Scholar] [CrossRef] [PubMed]
- Freemark, K.E.; Probst, J.R.; Dunning, J.B.; Hejl, S.J. Status and management of neotropical migratory birds: 21–25 September 1992, Estes Park, Colorado. In General Technical Reports RM-229; Finch, D.M., Stangel, P.W., Eds.; Department of Agriculture, Forest Service: Washington, DC, USA, 1993; pp. 346–352. [Google Scholar]
- Instituto Nacional de Estadística y Geografía (INEGI). Conjunto de Datos Vectoriales Climatología, Escala 1:250,000, Serie VI. Available online: https://www.inegi.org.mx/temas/climatologia/ (accessed on 5 May 2024).
- Rzedowski, J. Vegetación de México; Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad: México City, México, 2006; p. 504. Available online: https://www.biodiversidad.gob.mx/publicaciones/librosDig/pdf/VegetacionMxPort.pdf (accessed on 5 May 2024).
- García-Martínez, M.A.; Rodríguez, A. Vegetación y flora fanerogámica del área natural protegida Piedras Bola, Jalisco, México. Polibotánica 2018, 46, 71–90. [Google Scholar]
- Instituto Nacional de Estadística y Geografía (INEGI). Conjunto de Datos Vectoriales de Uso Del Suelo y Vegetación, Escala 1:250,000, Serie VII. Available online: https://www.inegi.org.mx/temas/usosuelo/default.html#Descargas/ (accessed on 5 May 2024).
- Rendón, S.F.J.; Cedano, M.M. Guía de excursión botánica: Los Guachimontones y humedales de Teuchitlán. In Guía de las Excursiones Botánicas en Jalisco. XVIII Congreso Mexicano; Ramírez, D.R., Reynoso, D.J.J., Rodríguez, C.A., Eds.; Universidad de Guadalajara: Guadalajara, México, 2010; pp. 88–106. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 7.5. Available online: https://www.robertkcolwell.org/pages/1407 (accessed on 5 May 2024).
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Cortés-Gómez, A.M.; Ruiz-Agudelo, C.A.; Valencia-Aguilar, A.; Ladle, R.J. Ecological functions of neotropical amphibians and reptiles: A review. Univ. Sci. 2015, 2, 229–245. [Google Scholar] [CrossRef]
- Laliberte, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 9, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Laliberte, E.; Legendre, P.; Shipley, B. FD Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0–12, 2014. Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 9 January 2022).
- RStudio Team. RStudio: Integrated Development for R. Computer Software v0.98.1074. Available online: www.rstudio.com/ (accessed on 5 May 2024).
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E Ltd.: Plymouth, UK, 2014. [Google Scholar]
- Diario Oficial de la Federación. Secretaría de Medio Ambiente y Recursos Naturales NOM–059–SEMARNAT–2010. Protección Ambiental. Especies Nativas de México de Flora y Fauna Silvestres-Categorías de Riesgo y Especificaciones Para su Inclusión, Exclusión o Cambio. Lista de Especies en Riesgo. Modificación del Anexo Normativo III, Publicada en el Diario Oficial de la Federación el 30 de Diciembre del 2010, Págs.: 1–78. Available online: https://www.dof.gob.mx/nota_detalle.php?codigo=5578808&fecha=14/11/2019#gsc.tab=0 (accessed on 5 May 2024).
- IUCN. The IUCN Red List of Threatened Species. 2021. Available online: https://www.iucnredlist.org/ (accessed on 5 May 2024).
- Luja, V.H.; López, J.A.; Cruz-Elizalde, R.; Ramírez-Bautista, A. Herpetofauna inside and outside from a natural protected area: The case of Reserva Estatal de la Biósfera Sierra San Juan, Nayarit, Mexico. Nat. Conserv. 2017, 21, 15–38. [Google Scholar] [CrossRef]
- Vega-Agavo, M.I.; Suazo-Ortuño, E.; Lopez-Toledo, L.; Gómez-Tagle, A.; Sillero, N.; Pineda-López, R.; Alvarado-Díaz, J. Influence of avocado orchard landscapes on amphibians and reptiles in the tans-Mexican volcanic belt. Biotropica 2021, 53, 1631–1645. [Google Scholar] [CrossRef]
- Suazo-Ortuño, I.; Alvarado-Díaz, J.; Mendoza, E.; López-Toledo, L.; Lara-Uribe, N.; Márquez-Camargo, C.; Paz-Gutiérrez, J.G.; Rangel-Orozco, J.D. High resilience of herpetofaunal communities in a human-modified tropical dry forest landscape in western Mexico. Trop. Conserv. Sci. 2015, 8, 396–423. [Google Scholar] [CrossRef]
- Suazo-Ortuño, I.; Alvarado-Díaz, J.; Martínez-Ramos, M. Riparian Areas and Conservation of Herpetofauna in a Tropical Dry Forest in Western Mexico. Biotropica 2011, 43, 237–245. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness, and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- McCranie, J.R. Crotalus basiliscus (Cope) Mexican west coast rattlesnake. Cat. Am. Amphib. Rept. 1981, 283, 1–2. Available online: https://repositories.lib.utexas.edu/browse/subject?scope=3ee72c9e-726f-45fd-8c72-4e7c583b3d74&value=Crotalus%20basiliscus&bbm.return=1 (accessed on 5 May 2024).
- Shine, R. The evolution of viviparity in reptiles: An ecological analysis. Biol. Reptil. 1985, 15, 605–694. [Google Scholar]
- Webb, J.K.; Shine, R.; Christian, K.A. The adaptive significance of reptilian viviparity in the tropics: Testing the maternal manipulation hypothesis. Evolution 2006, 60, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Tinkle, D.W.; Gibbons, J.W. The distribution and evolution of viviparity in reptiles. Misc. Publ. Mus. Zool. Univ. Mich. 1977, 154, 1–55. [Google Scholar]
- Bjorndal, K.A. Fermentation in reptiles and amphibians. In Gastrointestinal Microbiology; Mackie, R.I., White, B.A., Eds.; Springer Science+ Business Media: New York, NY, USA, 1997; Volume 1, pp. 199–230. [Google Scholar]
- Durtsche, D. Ontogenetic plasticity of food habits in the Mexican spiny-tailed iguana, Ctenosaura pectinata. Oecologia 2000, 124, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Macip-Ríos, R.; Sustaita-Rodriguez, V.H.; Barrios-Quiroz, G.; Casas-Andreu, G. Alimentary Habits of the Mexican Mud Turtle (Kinosternon integrum) in Tonatico, Estado de México. Chelonian Conserv. Biol. 2010, 9, 90–97. [Google Scholar] [CrossRef]
- Tuff, K.T.; Tuff, T.; Davies, K.F. A framework for integrating thermal biology into fragmentation research. Ecol. Lett. 2016, 19, 361–374. [Google Scholar] [CrossRef]
- Suazo-Ortuño, I.; Alvarado-Díaz, J.; Martínez-Ramos, M. Effects of conversion of dry tropical forest to agricultural mosaic on herpetofaunal assemblage. Conserv. Biol. 2008, 22, 362–374. [Google Scholar] [CrossRef]
- Rosas-Espinoza, V.C.; Peña-Joya, K.E.; Álvarez-Grzybowska, E.; Godoy-González, A.A.; Santiago-Pérez, A.L.; Rodríguez-Zaragoza, F.A. Amphibian Taxonomic and Functional Diversity in a Heterogeneous Landscape of West-Central Mexico. Diversity 2022, 14, 738. [Google Scholar] [CrossRef]
- Marroquín-Páramo, J.A.; Suazo-Ortuño, I.; Mendoza, E.; Alvarado-Díaz, J.; Siliceo-Cantero, H.H. Herpetofaunal diversity in avocado orchards and in conserved habitats in Michoacán, Mexico. Rev. Mex. Biodivers. 2017, 88, 234–240. [Google Scholar] [CrossRef]
- Peña-Joya, K.E.; Cupul-Magaña, F.G.; Rodríguez-Zaragoza, F.A.; Moreno, C.E.; Téllez-López, J. Spatio-temporal discrepancies in lizard species and functional diversity. Community Ecol. 2020, 21, 1–12. [Google Scholar] [CrossRef]
- Cruz-Elizalde, R.; Berriozabal-Islas, C.; Hernández-Salinas, U.; Martínez-Morales, M.A.; Ramírez-Bautista, A. Amphibian species richness and diversity in a modified tropical environment of central Mexico. Trop. Ecol. 2016, 57, 407–417. [Google Scholar]
- Leyte-Manrique, A.; Buelna-Chontal, A.A.; Torres-Diaz, M.A.; Berriozabal-Islas, C.; Maciel-Mata, C.A.A. Comparison of Amphibian and Reptile Diversity Between Disturbed and Undisturbed Environments of Salvatierra, Guanajuato, Mexico. Trop. Conserv. Sci. 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Gibbons, J.W.; Scott, D.E.; Ryan, T.J.; Buhlmann, K.A.; Tuberville, T.D.; Metts, B.S.; Greene, J.L.; Mills, T.; Leiden, Y.; Poppy, S.; et al. The global decline of reptiles, déjà vu amphibians: Reptile species are declining on a global scale. Six significant threats to reptile populations are habitat loss and degradation, introduced invasive species, environmental pollution, disease, unsustainable use, and global climate change. Bioscience 2000, 50, 653–666. [Google Scholar]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Sekercioglu, C.H.; Buchart, S.H.M.; Kauffman, M. A global analysis of traits predicting species sensitivity to habitat fragmentation. Glob. Ecol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- Macip-Ríos, R.; Casas-Andreu, G. Los cafetales en México y su importancia para la conservacion de los anfibios y reptiles. Acta Zool. Mex. 2008, 24, 143–159. [Google Scholar] [CrossRef]
- Macip-Ríos, R.; Muñoz-Alonso, A. Diversidad de lagartijas en cafetales y bosque primario en el Soconusco Chiapaneco. Rev. Mex. Biodiv. 2008, 79, 185–195. [Google Scholar] [CrossRef]
- Wilson, L.D.; Mata-Silva, V.; Johnson, J.D. A conservation reassessment of the reptiles of Mexico based on the EVS measure. Amphib. Reptile Conserv. 2013, 7, 97–127. [Google Scholar]
- Brandon, K.; Gorenflo, L.J.; Rodrigues, A.S.; Waller, R.W. Reconciling biodiversity conservation, people, protected areas, and agricultural suitability in Mexico. World Dev. 2005, 33, 1403–1418. [Google Scholar] [CrossRef]
- Ramirez-Reyes, C.; Sims, K.; Potapov, P.; Radeloff, V.C. Payments for ecosystem services in Mexico reduce forest fragmentation. Ecol. Appl. 2018, 28, 1982–1997. [Google Scholar] [CrossRef] [PubMed]
- Costedoat, S.; Corbera, E.; Ezzine-de-Blas, D.; Honey-Rosés, J.; Baylis, K.; Castillo-Santiago, M.A. How effective are biodiversity conservation payments in Mexico? PLoS ONE 2015, 10, e0119881. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.L.; Foster, C.N.; Blanchard, W.; Florance, D.; Michael, D.R.; Lindenmayer, D.B. Reversing habitat loss: An experimental test of the interactive effects of grazing exclusion and surface rock restoration on reptile conservation. J. Appl. Ecol. 2023, 60, 1778–1789. [Google Scholar] [CrossRef]
- Donald, P.F.; Evans, A.D. Habitat connectivity and matrix restoration: The wider implications of agri-environment schemes. J. Appl. Ecol. 2006, 43, 209–218. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family/Species | POF | OF | RH-TF | SV-TF | RH-TDF | TDF | HPTDF | RH-C | CO | CA | Endemism | NOM–059 | IUCN | Indiv |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plot Numbers | 10 | 10 | 5 | 7 | 5 | 5 | 6 | 9 | 5 | 5 | ||||
| Anguidae | ||||||||||||||
| Elgaria kingii | X | X | X | X | N | Pr | LC | 12 | ||||||
| Dactyloidae | ||||||||||||||
| Anolis nebulosus | X | X | X | X | X | X | X | X | X | E | - | LC | 199 | |
| Iguanidae | ||||||||||||||
| Ctenosaura pectinata | X | X | X | X | X | X | E | A | LC | 23 | ||||
| Phrynosomatidae | ||||||||||||||
| Sceloporus dugesii | X | X | E | - | LC | 123 | ||||||||
| Sceloporus heterolepis | X | X | X | X | X | X | E | - | LC | 9 | ||||
| Sceloporus horridus | X | X | X | X | X | LC | 71 | |||||||
| Sceloporus torquatus | X | E | - | LC | 1 | |||||||||
| Sceloporus spinosus | X | E | - | LC | 1 | |||||||||
| Sceloporus utiformis | X | X | X | X | X | E | - | LC | 17 | |||||
| Urosaurus bicarinatus | X | X | X | E | - | LC | 13 | |||||||
| Phyllodactylus lanei | X | E | - | LC | 1 | |||||||||
| Scincidae | ||||||||||||||
| Plestiodon dugesii | X | X | X | X | E | - | LC | 83 | ||||||
| Plestiodon callicephalus | X | X | N | - | LC | 1 | ||||||||
| Teiidae | ||||||||||||||
| Aspidoscelis costatus | X | X | X | X | E | Pr | LC | 60 | ||||||
| Aspidoscelis communis | X | E | Pr | LC | 17 | |||||||||
| Colubridae | ||||||||||||||
| Drymarchon melanurus | X | X | X | X | N | - | LC | 2 | ||||||
| Lampropeltis ruthveni | X | X | X | X | N | A | NT | 5 | ||||||
| Masticophis mentovarius | X | X | X | X | X | E | - | LC | 7 | |||||
| Masticophis bilineatus | X | X | N | - | LC | 1 | ||||||||
| Oxybelis microphthalmus | X | X | X | N | - | LC | 1 | |||||||
| Pituophis deppei | X | X | LC | 1 | ||||||||||
| Senticolis triaspis | X | X | N | - | LC | 1 | ||||||||
| Tantilla bocourti | X | X | E | - | LC | 12 | ||||||||
| Trimorphodon tau | X | X | E | - | LC | 1 | ||||||||
| Dipsadidae | ||||||||||||||
| Hypsiglena torquata | X | N | Pr | LC | 1 | |||||||||
| Leptodeira maculata | X | E | Pr | LC | 1 | |||||||||
| Leptodeira splendida | X | X | E | Pr | LC | 4 | ||||||||
| Rhadinaea hesperia | X | X | E | - | LC | 1 | ||||||||
| Rhadinaea taeniata | X | E | - | LC | 1 | |||||||||
| Elapidae | ||||||||||||||
| Micrurus distans | X | X | X | E | Pr | LC | 4 | |||||||
| Leptoyphlopidae | ||||||||||||||
| Rena humilis | X | X | N | - | LC | 1 | ||||||||
| Natricidae | ||||||||||||||
| Thamnophis copei | X | E | Pr | Vu | 1 | |||||||||
| Storeria storerioides | X | X | X | E | - | LC | 5 | |||||||
| Thamnophis eques | X | X | N | A | LC | 9 | ||||||||
| Thamnophis cyrtopsis | X | X | X | N | A | LC | 2 | |||||||
| Typhlopidae | ||||||||||||||
| Indotyphlops braminus | X | X | X | EX | - | - | 4 | |||||||
| Viperidae | ||||||||||||||
| Agkistrodon bilineatus | X | X | X | N | Pr | NT | 1 | |||||||
| Crotalus basiliscus | X | X | X | E | Pr | LC | 1 | |||||||
| Kinosternidae | ||||||||||||||
| Kinosternon integrum | X | X | X | X | X | X | E | Pr | LC | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosas-Espinoza, V.C.; Rodríguez-Zaragoza, F.A.; Álvarez-Grzybowska, E.; Peña-Joya, K.E.; Santiago-Pérez, A.L.; Godoy-González, A.A.; Huerta-Martínez, F.M. Taxonomic and Functional Diversity of Reptiles in a Heterogeneous Landscape of Jalisco State, West-Central Mexico. Diversity 2024, 16, 394. https://doi.org/10.3390/d16070394
Rosas-Espinoza VC, Rodríguez-Zaragoza FA, Álvarez-Grzybowska E, Peña-Joya KE, Santiago-Pérez AL, Godoy-González AA, Huerta-Martínez FM. Taxonomic and Functional Diversity of Reptiles in a Heterogeneous Landscape of Jalisco State, West-Central Mexico. Diversity. 2024; 16(7):394. https://doi.org/10.3390/d16070394
Chicago/Turabian StyleRosas-Espinoza, Verónica Carolina, Fabián Alejandro Rodríguez-Zaragoza, Eliza Álvarez-Grzybowska, Karen Elizabeth Peña-Joya, Ana Luisa Santiago-Pérez, Arquímedes Alfredo Godoy-González, and Francisco Martín Huerta-Martínez. 2024. "Taxonomic and Functional Diversity of Reptiles in a Heterogeneous Landscape of Jalisco State, West-Central Mexico" Diversity 16, no. 7: 394. https://doi.org/10.3390/d16070394
APA StyleRosas-Espinoza, V. C., Rodríguez-Zaragoza, F. A., Álvarez-Grzybowska, E., Peña-Joya, K. E., Santiago-Pérez, A. L., Godoy-González, A. A., & Huerta-Martínez, F. M. (2024). Taxonomic and Functional Diversity of Reptiles in a Heterogeneous Landscape of Jalisco State, West-Central Mexico. Diversity, 16(7), 394. https://doi.org/10.3390/d16070394