
Age Determination of Rocky Mountain Ridged Mussels (Gonidea angulata) in the Okanagan Basin, Canada
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Mussels
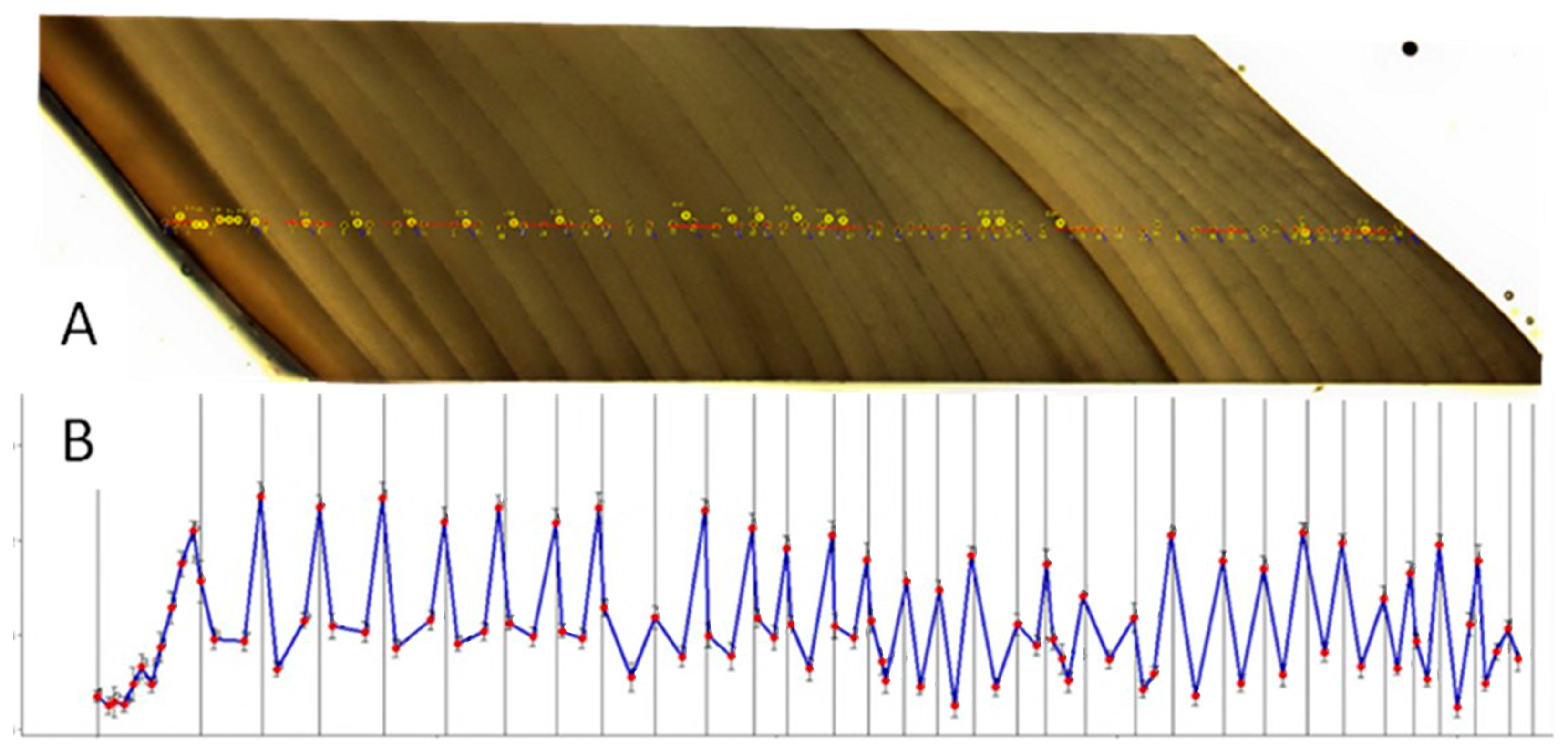
2.2. Age Determination and Validation
2.3. Data Analysis
3. Results
3.1. Mussel Age Determination
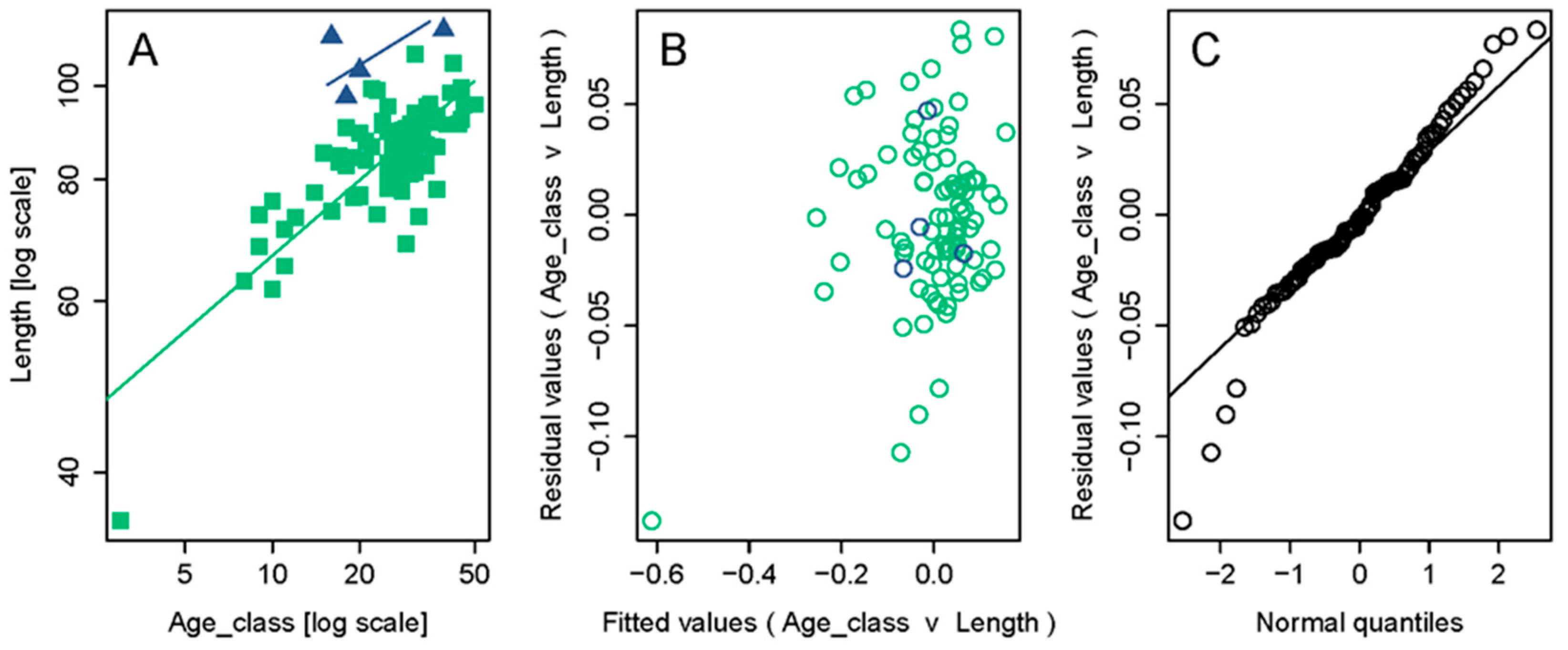
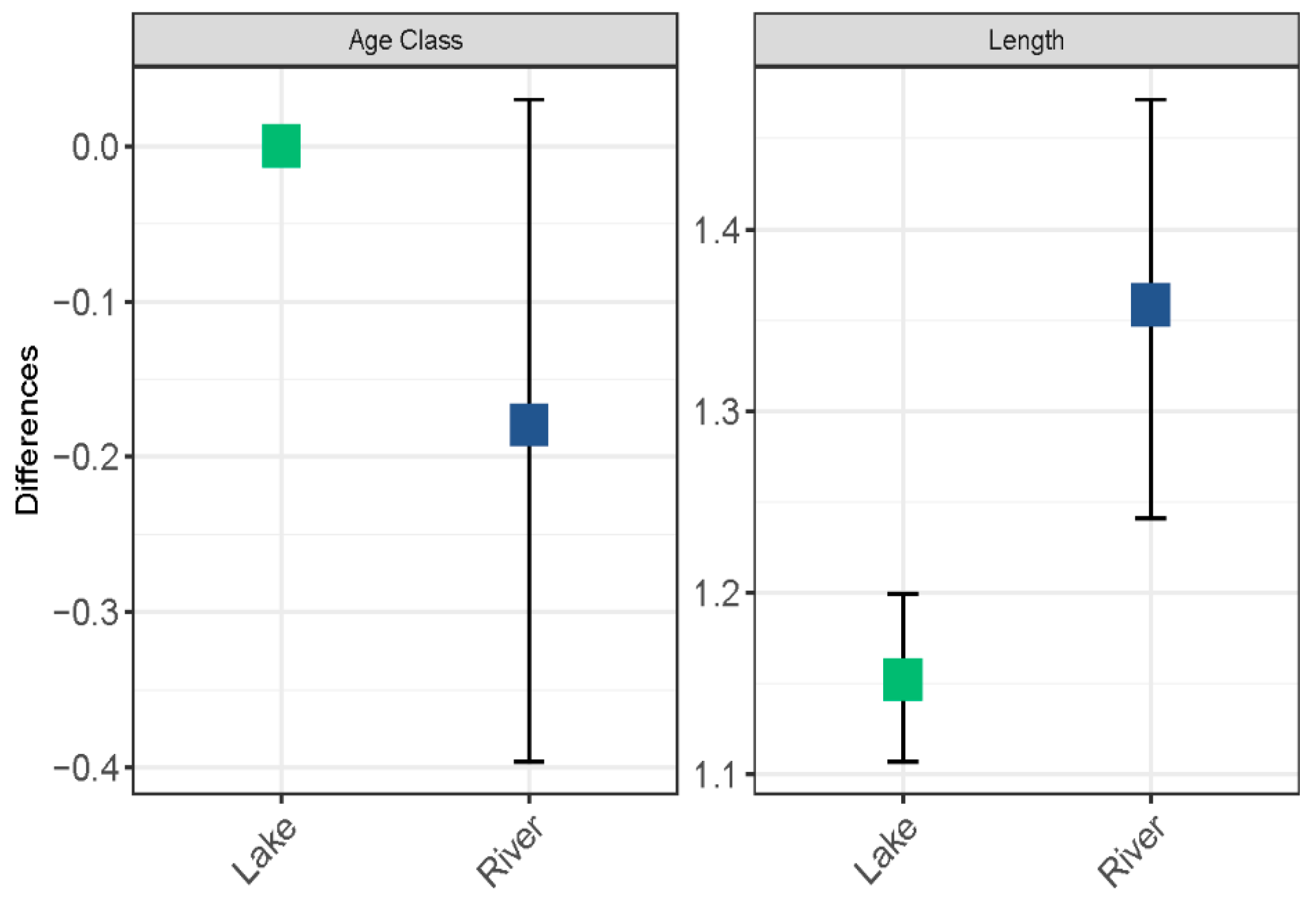
3.2. Mussel Age and Growth Patterns
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.; Soto, D.; Stiassny, M.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Vari, A.; Poschun, S.A.; Eros, T.; Hein, T.; Pataki, B.; Ioja, I.; Adamescu, C.M.; Gerhardt, A.; Gruber, T.; Dedic, A.; et al. Freshwater systems and ecosystem services: Challenges and chances for cross-fertilization of disciplines. Ambio 2022, 51, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, B.; Bouchard, R.W.; Dietz, R.; Hornbach, D.; Monson, P.; Sietman, B.; Wasley, D. Freshwater Mussels, Ecosystem Services, and Clean Water Regulation in Minnesota: Formulating an Effective Conservation Strategy. Water 2023, 15, 2560. [Google Scholar] [CrossRef]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Arthington, A.H.; Dulvy, N.K.; Gladstone, W.; Winfield, I.J. Fish conservation in freshwater and marine realms: Status, threats and management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 838–857. [Google Scholar] [CrossRef]
- Haag, W.R.; Culp, J.J.; McGregor, M.A.; Bringolf, R.; Stoeckel, J.A. Growth and survival of juvenile freshwater mussels in streams: Implications for understanding enigmatic mussel declines. Freshw. Sci. 2019, 38, 753–770. [Google Scholar] [CrossRef]
- Stagliano, D. Western Ridged Mussel Age-Length Determinations for Idaho Rivers; Report to the US Fish and Wildlife Service; US Fish and Wildlife Service: Chubbuck, ID, USA, 2023. [Google Scholar]
- Vaughn, C.C. Ecosystem services provided by freshwater mussels. Hydrobiologia 2018, 810, 15–27. [Google Scholar] [CrossRef]
- Taylor, D.W. Freshwater molluscs of California: A distributional checklist. Calif. Fish Game 1981, 67, 140–163. [Google Scholar]
- COSEWIC. COSEWIC Assessment and Status Report on the Rocky Mountain Ridged Mussel Gonidea angulata in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2010; 56p. [Google Scholar]
- Gurney, L.J.; Mundy, C.; Porteus, M.C. Determining age and growth of abalone using stable oxygen isotopes: A tool for fisheries management. Fish. Res. 2005, 72, 353–360. [Google Scholar] [CrossRef]
- MacConnachie, S.; Dealy, L.; Wade, J.; Grant, P. Rocky Mountain Ridged Mussel (Gonidea angulata) Index Site Surveys in the Okanagan Basin, British Columbia, 2017–2020; Canadian Manuscript Report of Fisheries and Aquatic Sciences; Fisheries and Oceans Canada: Nanaimo, BC, Canada, 2021; Volume 3211, 9p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 24 February 2024).
- Warton, D.I.; Weber, N.C. Common slope tests for errors-in-variables models. Biom. J. 2002, 44, 161–174. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. 2006, 81, 259–291. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Freire, S.; Read, L.; Lewis, T.R. Assessing Bird Collisions in the United Kingdom: Modelling Frequency of Birdstrike from Road and Rail Mortality Using a Bayesian Hierarchical Approach; Specialist Research Project; Kingston Maurward College: Dorchester, UK, 2020; 28p. [Google Scholar]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef]
- Hooten, M.B.; Hobbs, N.T. A guide to Bayesian model selection for ecologists. Ecol. Monogr. 2015, 85, 3–28. [Google Scholar] [CrossRef]
- Lewis, T.R.; Griffin, R.K.; Martin, I.M.; Figueroa, A.; Ray, J.M.; Feltham, J.; Grant, P.B.C. Ecology and morphology of the dwarf bromeliad boa Ungaliophis panamensis (Squamata, Boidae, Ungaliophiinae) in Costa Rica and Panama. Neotrop. Biol. Conserv. 2021, 16, 317–331. [Google Scholar] [CrossRef]
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. smatr 3—An R package for estimation and inference about allometric lines. Methods Ecol. Evol. 2012, 3, 257–259. [Google Scholar] [CrossRef]
- Taskinen, S.; Warton, D.I. Robust estimation and inference for bivariate line-fitting in allometry. Biom. J. 2011, 53, 652–672. [Google Scholar] [CrossRef] [PubMed]
- Dunn, P.; Smyth, G. Randomized quantile residuals. J. Comput. Graph. Stat. 1996, 5, 236–244. [Google Scholar] [CrossRef]
- Wickham, H. Reshaping data with the reshape Package. J. Stat. Softw. 2007, 21, 1–20. Available online: http://www.jstatsoft.org/v21/i12/ (accessed on 24 February 2024). [CrossRef]
- Plummer, M. JAGS: A program for analysis of Bayesian graphical models using Gibbs sampling. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing (DSC 2003), Vienna, Austria, 20–22 March 2003. [Google Scholar]
- Denwood, M.J. runjags: An R package providing interface utilities, model templates, parallel computing methods and additional distributions for MCMC models in JAGS. J. Stat. Softw. 2016, 71, 1–25. [Google Scholar] [CrossRef]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis, 3rd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2013. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Haag, W.R.; Rype, A.L. Growth and longevity in freshwater mussels: Evolutionary and conservation implications. Biol. Rev. 2011, 86, 225–247. [Google Scholar] [CrossRef]
- Dennis, B.; Munholland, P.L.; Scott, J.M. Estimation of growth and extinction parameters for endangered species. Ecol. Monogr. 1991, 61, 115–143. [Google Scholar] [CrossRef]
- Hastie, L.C.; Young, M.R.; Boon, P.J. Growth characteristics of freshwater pearl mussels Margaritifera margaritifera (L.). Freshw. Biol. 2000, 43, 243–256. [Google Scholar] [CrossRef]
- Black, B.A.; Dunham, J.B.; Blundon, B.W.; Brim-Box, J.; Tepley, A.J. Long-term growth-increment chronologies reveal diverse influences of climate forcing on freshwater and forest biota in the Pacific Northwest. Glob. Chang. Biol. 2015, 21, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Downing, J.A.; Haag, W.R.; King, T.L.; Layzer, J.B.; Newton, T.J.; Nichols, S.J. Changing perspectives on pearly mussels, America’s most imperiled animals. BioScience 2004, 54, 429–439. [Google Scholar] [CrossRef]
- Peharda, M.; Zupan, I.; Bavcevic, L.; Frankic, A.; Klanjscek, T. Growth and condition index of mussel Mytilus galloprovincialis in experimental integrated aquaculture. Aquac. Res. 2007, 38, 1714–1720. [Google Scholar] [CrossRef]
- Randall, R.G.; Kelso, J.R.M.; Minns, C.K. Fish productivity in freshwater: Are rivers more productive that lakes? Can. J. Fish. Aquat. Sci. 1995, 52, 631–643. [Google Scholar] [CrossRef]
- Strayer, D. Understanding how nutrient cycles and freshwater mussels (Unionoida) affect one another. Hydrobiologia 2014, 735, 277–292. [Google Scholar] [CrossRef]
- Strayer, D.L.; Malcom, H.M. Causes of recruitment failure in freshwater mussel populations in southeastern New York. Ecol. Appl. 2012, 22, 1780–1790. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grant, P.; Wade, J.; Lewis, T.R.; Castañeda, R.A.; Branquinho, E.; MacConnachie, S.; Wischniowski, S.; Campbell, B. Age Determination of Rocky Mountain Ridged Mussels (Gonidea angulata) in the Okanagan Basin, Canada. Diversity 2024, 16, 415. https://doi.org/10.3390/d16070415
Grant P, Wade J, Lewis TR, Castañeda RA, Branquinho E, MacConnachie S, Wischniowski S, Campbell B. Age Determination of Rocky Mountain Ridged Mussels (Gonidea angulata) in the Okanagan Basin, Canada. Diversity. 2024; 16(7):415. https://doi.org/10.3390/d16070415
Chicago/Turabian StyleGrant, Paul, Joy Wade, Todd R. Lewis, Rowshyra A. Castañeda, Emma Branquinho, Sean MacConnachie, Stephen Wischniowski, and Barbara Campbell. 2024. "Age Determination of Rocky Mountain Ridged Mussels (Gonidea angulata) in the Okanagan Basin, Canada" Diversity 16, no. 7: 415. https://doi.org/10.3390/d16070415