
A New Species of Smithophis Giri et al. 2019 from the Indo-Burma Region

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Collection and Morphology
2.2. Morphology
2.3. Nomenclature Acts
2.4. Molecular Methods
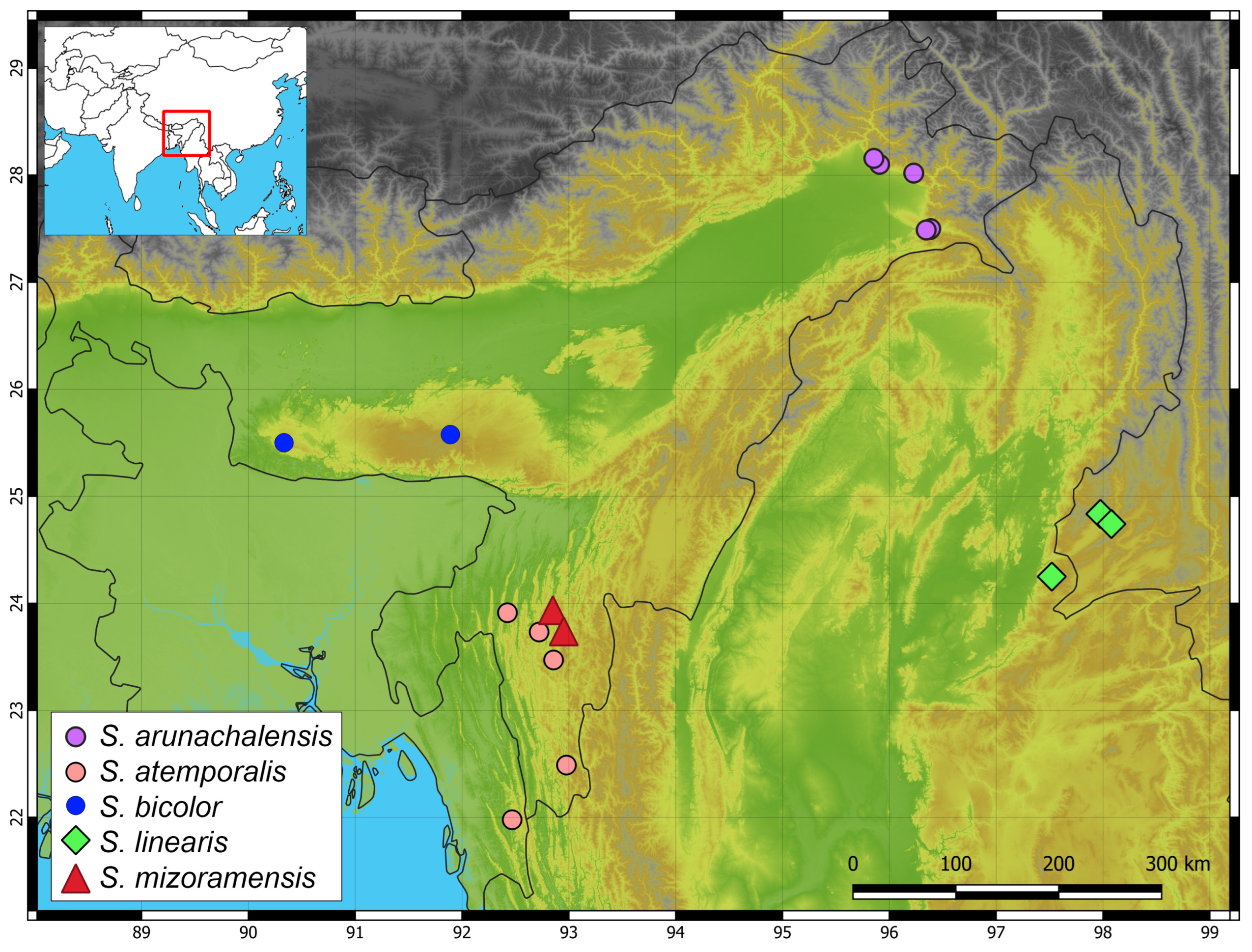
3. Results
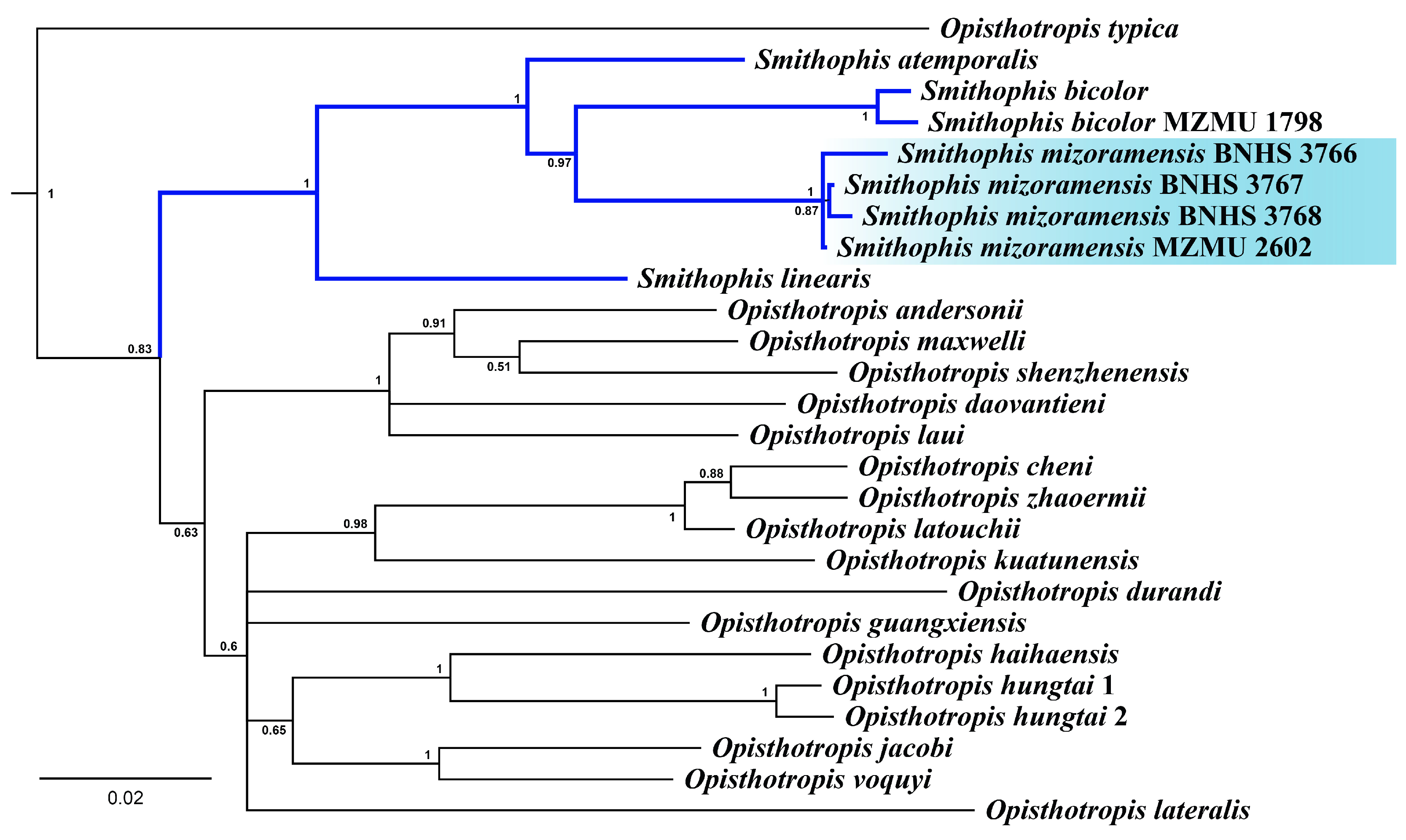
3.1. Molecular Phylogeny
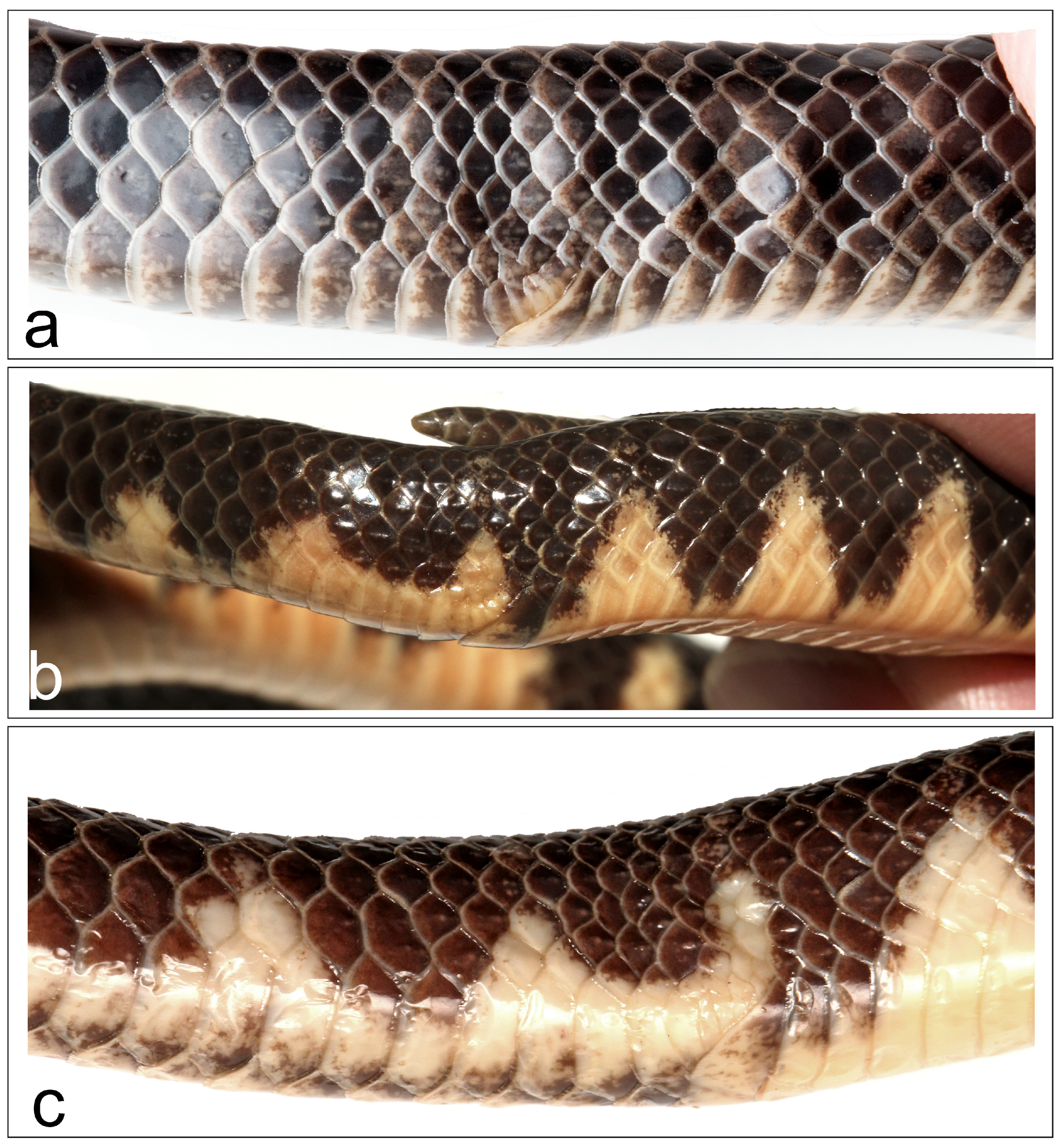
3.2. Morphology
3.3. Systematics
4. Discussion and Conclusions
| Key to members of the genus Smithophis |
| 1a. Temporal scales absent..................................................................................... S. atemporalis |
| 1b. Temporal scales present........................................................................................................2 |
| 2a. Sacral scales keeled in males.................................................................................................3 |
| 2b. Sacral scales smooth in both sexes.......................................................................................4 |
| 3a. Dorsum black, venter yellow with a zig-zag ventrolateral pattern; sacral scales strongly keeled................................................................................................... S. arunachalensis |
| 3b. Dorsal in shades of olive (never black) with indistinct brown reticulate patterns; sacral scales moderately keeled.......................................................................S. mizoramensis sp. nov. |
| 4a. Dorsum brown or grayish-brown without any markings; circumorbital scales 4–5..........................................................................................................................................S. bicolor |
| 4b. Dorsum brown with narrow dark longitudinal stripes; circumorbital scales 6–7.........................................................................................................................................S. linearis |
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vogel, G.; Chen, Z.; Deepak, V.; Gower, D.J.; Jingsong, S.H.I.; Ding, L.; Mian, H.O.U. A new species of the genus Smithophis (Squamata: Serpentes: Natricidae) from Southwestern China and northeastern Myanmar. Zootaxa 2020, 4803, 51–74. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 2 July 2024).
- Ruatpuii, R.; Biakzuala, L.; Santra, V.; Lalremsanga, H.T. Additional notes on morphology and distributional records of the snake genus Smithophis Giri, Gower, Das, Lalremsanga, Lalronunga, Captain Et Deepak, 2019 (Squamata: Serpentes: Natricidae) from north-east India. Russ. J. Herpetol. 2022, 29, 331–340. [Google Scholar] [CrossRef]
- Das, A.; Deepak, V.; Captain, A.; Wade, E.O.Z.; Gower, D.J. Description of a new species of Smithophis Giri et al. 2019 (Serpentes: Colubridae: Natricinae) from Arunachal Pradesh, India. Zootaxa 2020, 4860, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Giri, V.B.; Gower, D.J.; Das, A.; Lalremsanga, H.T.; Lalronunga, S.; Captain, A.; Deepak, V. A new genus and species of natricine snake from Northeast India. Zootaxa 2019, 4603, 241–264. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A. Fauna of British India, Ceylon and Burma, Including the Whole of the Indo-Chinese Sub-Region. Reptilia and Amphibia. Vol. 3. Serpentes; Taylor and Francis: London, UK, 1943; p. 440. [Google Scholar]
- Chandramouli, S.R.; Karthik, P.; Naveen, R.; Babu, S.; Karunakaran, P.; Kumara, H. A Two-Colored Forestsnake, Smithophis bicolor (Blyth 1855) (Reptilia: Natricidae), from the Khasi Hills, Meghalaya, India. Reptil. Amphib. 2021, 28, 24–25. [Google Scholar] [CrossRef]
- Patel, H.; Thackeray, T.; Campbell, P.D.; Mirza, Z.A. Systematic assessment of Hebius beddomei (Günther, 1864) (Serpentes: Colubridae: Natricinae) with description of a new genus and a new allied species from the Western Ghats, India. Taxonomy 2023, 3, 415–434. [Google Scholar] [CrossRef]
- Mirza, Z.A.; Vyas, R.; Patel, H.; Maheta, J.; Sanap, R. A new Miocene-Divergent lineage of Old World Racer snake from India. PLoS ONE 2016, 11, e0154301. [Google Scholar] [CrossRef] [PubMed]
- Dowling, H.; Savage, J. A guide to the snakes hemipenis: A survey of basic structure and systematic characteristics. Zoologica 1960, 45, 17–31. [Google Scholar] [CrossRef]
- Zaher, H. Hemipenial morphology of the South American Xenodontine Snakes, with a proposal for a monophyletic Xenodontinae and a reappraisal of colubrid hemipenes. Bull. Am. Museum Nat. Hist. 1999, 240, 1–240. [Google Scholar]
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature; International Trust for Zoological Nomenclature: London, UK, 1999. [Google Scholar]
- Burbrink, F.T.; Lawson, R.; Slowinski, J.B. Mitochondrial DNA phylogeography of the polytypic North American rat snake (Elaphe obsoleta): A critique of the subspecies concept. Evolution 2000, 54, 2107–2118. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Higgins, D.; Gibson, T. ClustalW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.; Wong, T.; Von Haeseler, A.; Jermiin, L. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.; Schmidt, H.; Chernomor, O.; Schrempf, D.; Woodhams, M.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for Phylogenetic Inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Deepak, V.; Cooper, N.; Poyarkov, N.A.; Kraus, F.; Burin, G.; Das, A.; Narayanan, S.; Streicher, J.W.; Smith, S.-J.; Gower, D.J. Multilocus phylogeny, natural history traits and classification of natricine snakes (Serpentes: Natricinae). Zool. J. Linn. Soc. 2022, 195, 279–298. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Pawar, S.S.; Birand, A.C.; Ahmed, M.F.; Sengupta, S.; Raman, T.R.S. Conservation biogeography in north-east India: Hierarchical analysis of cross-taxon distributional congruence. Divers. Distrib. 2007, 13, 53–65. [Google Scholar] [CrossRef]
- Lalremsanga, H.T.; Bal, A.K.; Vogel, G.; Biakzuala, L. Molecular phylogenetic analyses of lesser known colubrid snakes reveal a new species of Herpetoreas (Squamata: Colubridae: Natricinae), and new insights into the systematics of Gongylosoma scriptum and its allies from northeastern India. Salamandra 2022, 58, 101–115. [Google Scholar]
- Lalremsanga, H.T.; Muansanga, L.; Vabeiryureilai, M.; Mirza, Z.A. A new species of parachute gecko of the subgenus Ptychozoon (Sauria: Gekkonidae: Gekko) from the Indo-Burma Region. Salamandra 2023, 59, 125–135. [Google Scholar]
- Idiiatullina, S.S.; Van Nguyen, T.; Pawangkhanant, P.; Suwannapoom, C.; Chanhome, L.; Mirza, Z.A.; David, P.; Vogel, G.; Poyarkov, N.A. An integrative taxonomic revision of the Trimeresurus popeiorum Group of pitvipers (Reptilia: Serpentes: Viperidae) with descriptions of two new species from the Indo-Burma Biodiversity Hotspot. Vertebr. Zool. 2024, 74, 303–342. [Google Scholar] [CrossRef]
- Agarwal, I.; Mahony, S.; Giri, V.B.; Chaitanya, R.; Bauer, A.M. Six new Cyrtodactylus (Squamata: Gekkonidae) from Northeast India. Zootaxa 2018, 4524, 501–535. [Google Scholar] [CrossRef] [PubMed]
- Lalronunga, S.; Lalhmangaiha, K.; Zosangliana, I.; Lalhmingliani, E.; Gower, D.J.; Das, A.; Deepak, V. A new species of Stoliczkia Jerdon, 1870 (Serpentes: Xenodermidae) from Mizoram, India. Zootaxa 2021, 4996, 555–568. [Google Scholar] [CrossRef]
- Rathee, Y.S.; Purkayastha, J.; Lalremsanga, H.T.; Dalal, S.; Biakzuala, L.; Muansanga, L.; Mirza, Z.A. A new cryptic species of green pit viper of the genus Trimeresurus Lacépède, 1804 (Serpentes, Viperidae) from Northeast India. PLoS ONE 2022, 17, e0268402. [Google Scholar] [CrossRef] [PubMed]
- Purkayastha, J.; Lalremsanga, H.T.; Bohra, S.C.; Biakzuala, L.; Decemson, H.; Muansanga, L.; Vabeiryureilai, M.; Chauhan, S.; Rathee, Y.S. Four new bent-toed geckos (Cyrtodactylus Gray: Squamata: Gekkonidae) from Northeast India. Zootaxa 2021, 4980, 451–489. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BNHS 3766 | BNHS 3767 | BNHS 3768 | |
|---|---|---|---|
| Type status | holotype | paratype | paratype |
| Sex | male | female | male |
| SVL | 477.8 | 421.4 | 453.0 |
| TaL | 134.0 | 140.0 | 125.0 |
| TL | 611.8 | 561.4 | 578.0 |
| DSR | 17:17:17 | 17:17:17 | 17:17:17 |
| VS | 214 (4) | 200 (3) | 213 (3) |
| Sc | 73 | 67 | 74 |
| Anal | divided | divided | divided |
| SL | 5/5 (-) | 5/5 (3rd) | 5/5 (3rd) |
| IL | 8/7 | 7/7 | 7/7 |
| Temporals | 1 + 1 | 1 + 1 | 1 + 1 |
| STP | 11 | 10 | 11 |
| Circumorbital scales | 5 | 4 | 4 |
| Sacral keels | present | absent | present |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirza, Z.A.; Bhardwaj, V.K.; Lalmuanawma, J.C.; Choure, G.; Lalremsanga, H.T.; Vabeiryureilai, M.; Captain, A.; Zagade, A.; Patel, H. A New Species of Smithophis Giri et al. 2019 from the Indo-Burma Region. Diversity 2024, 16, 480. https://doi.org/10.3390/d16080480
Mirza ZA, Bhardwaj VK, Lalmuanawma JC, Choure G, Lalremsanga HT, Vabeiryureilai M, Captain A, Zagade A, Patel H. A New Species of Smithophis Giri et al. 2019 from the Indo-Burma Region. Diversity. 2024; 16(8):480. https://doi.org/10.3390/d16080480
Chicago/Turabian StyleMirza, Zeeshan A., Virender K. Bhardwaj, Jote Chawntual Lalmuanawma, Girish Choure, Hmar Tlawmte Lalremsanga, Mathipi Vabeiryureilai, Ashok Captain, Akshay Zagade, and Harshil Patel. 2024. "A New Species of Smithophis Giri et al. 2019 from the Indo-Burma Region" Diversity 16, no. 8: 480. https://doi.org/10.3390/d16080480
APA StyleMirza, Z. A., Bhardwaj, V. K., Lalmuanawma, J. C., Choure, G., Lalremsanga, H. T., Vabeiryureilai, M., Captain, A., Zagade, A., & Patel, H. (2024). A New Species of Smithophis Giri et al. 2019 from the Indo-Burma Region. Diversity, 16(8), 480. https://doi.org/10.3390/d16080480