Abstract
Harrison’s Rule (HR) postulates a positive allometry between host and parasite body sizes. We tested HR for Syringophilid quill mites parasitizing birds. Using host body mass and parasite body length as size indices, this pattern was absent in the Syringophilidae family and the Syringophilinae subfamily as a whole. However, when considering the parasite genera as units of study, as proposed originally by Harrison, we found that host body mass positively correlates with both male and female parasite body length in seven genera (Aulobia, Aulonastus, Neoaulonastus, Picobia, Neopicobia, Syringophilopsis, and Torotrogla). Most of these relationships were non-significant. On the contrary, male and female Syringophiloidus mites exhibited negative relationships with host mass (both non-significant). This apparent contradiction disappeared when we applied wing length as an index of host body size. Since species of this genus are specific to the host flight feathers (secondaries and also primaries), wing length is a more meaningful index of host body size than body mass. Overall, most cases corresponded to the positive direction predicted by Harrison when examined on the genus level. This finding also implies a surprising reliability of the genus concept, at least in this group of ectoparasites.
1. Introduction
Body size is a fundamental property of organisms that affects most aspects of their metabolism, behavior, and ecological relationships. For example, body size is directly related to fecundity. In comparisons across large phylogenetic distances, the fecundity of larger-sized animal taxa tends to be lower [1,2]. On the contrary, in intraspecies comparisons, larger organisms usually give birth to more offspring [3,4]. In parasites, larger body size associated with increased metabolism and higher fecundity can also cause the higher virulence of infections [5].
More than a century ago, Launcelot Harrison published an article about the feather lice of kiwis, with a voluminous explanation of his views about the coevolution of birds and lice [6]. He mostly speculated about the phylogenetic relationship between rails (Rallidae) and kiwis (Apterygidae), which he erroneously presumed due to the presence of the Rallicola spp. lice they are both infested by. Hidden in this lengthy and somewhat rambling text, he stated the following:
“…in general, when a genus is well distributed over a considerable number of nearly related hosts, the size of the parasite is roughly proportional to the size of the host…”
Briefly, he recognized a positive relationship between host and parasite body sizes in comparisons across species, presumed that the hosts are “nearly related,” and that the parasites are congeneric. Unfortunately, “near relatedness” is not well defined, and similarly, the genus concept is also an arbitrary taxonomic artifact rather than an objective criterium. Later authors called this relationship “Harrison’s Rule” (HR), although this is more a hypothesis than a rule. Several studies verified it for diverse parasite taxa [7,8,9,10,11,12,13].
Our present study aims to test HR using a taxon of a most severely space-limited avian ectoparasites, the quill mites (Syringophilidae). They are prostigmatic mites (Acari: Acariformes: Prostigmata) strictly associated with avian (Vertebrata: Aves) hosts. Their ancestors presumably appeared on feathered dinosaurs in the Early Jurassic [14,15]. All species live and reproduce inside the feather quills (calamus) [16]. Due to a basal divergence at an early stage of their evolution, they are divided into two subfamilies, which exhibit different anatomic site (or “niche”, in a certain sense) specificity on the host body surface [17]. Species of the subfamily Syringophilinae mainly inhabit the quills of secondaries. There can be two deviations from this niche: relatively small-bodied species tend to also infest the wing coverts, while large ones can be found in the primaries. (Some small-bodied Syringophiline genera, such as Aulonastus, Neoaulonastus, and some others, may infest wing coverts and also body feathers [18]). By contrast, representatives of the subfamily Picobiinae always infest the quills of body feathers (except for Calamincola [19], which is not included in the present study).
Quill mites live a peculiar way of life. A fertilized female (or rarely two of them) enters a developing feather’s calamus through the superior umbilicus opening [20]. This opening closes soon, and the female will produce a single (rarely more) male and several female offspring in this enclosed capsule. Then, the brother(s) fertilizes the sisters, and the next generation still lives enclosed in the same quill. After the grandchildren of the founding mother also fertilized each other, and mites basically fill the whole cavity of the calamus, fertilized females disperse to search for developing new feathers either on the same host or on another individual [16,21].
Thus, these parasites live a strictly space-limited life. If they grow too large, they die due to the lack of enough space in the enclosed capsule of the feather quill. Conversely, if they are too small, they cannot pierce the quill wall with their mouthparts to obtain nutrients from the surrounding tissues [18]. The presumed strict optimization of their body size makes them an optimal choice to test HR.
Syringophilid mites have a haplodiploid sex determination system [22]. Several species appear to lack male individuals, possibly parthenogenetic, or the rare males may be unknown due to sampling bias. Consequently, they are highly inbred and, therefore, almost totally free of sexual selection pressure, although male vs. female body sizes are still affected by different selection pressures [23]. For this reason, below we analyze the host–parasite body size allometry separately for male and female quill mites.
2. Materials and Methods
We used the total body length (μm) measurements of male and female quill mites gathered from the taxonomic literature. Data from the holotype specimens were used whenever possible. Otherwise, we calculated the average of the extreme values of the paratype series. Picobiine species may have two alternative female morphologies. Some of the “normal” (non-physogastric) females may develop a physogastric morphology characterized by a greatly enlarged abdomen, containing a few, but huge, eggs. We only considered the “normal” (non-physogastric) female size and excluded data on the much less frequent physogastric morphs. Note that the species descriptions are based on preparations embedded in Canada balsam on microscopic slides. Therefore, they may not reflect the true body length of living parasites, but rather the size of almost two-dimensional arbitrary preparations. Thus, their apparent body length may partially depend on the pressure exerted on the cover glasses when preparing the microscopic slides. From this point of view, it is important to note that most of the species involved in the present study were described by a relatively small group of authors who closely collaborate with each other, which may make their preparation techniques more homogenous than in most other ectoparasite taxa where species are described by many different authors working independently of each other. Data were obtained from species descriptions [24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94].
First, we intended to test HR at levels above the genus level, the Syringophilidae family, and the Syringophilinae and Picobiinae subfamilies, and then to test it at the genus level. Further, we tested male and female parasite body sizes separately. Therefore, we included only those genera for which both male and female body length, and host body mass, were known for an appropriate number of species (>5, an arbitrary limit). This criterion excludes parthenogenetic species [22] or those whose males we do not know. Overall, we considered eight genera and 110 species in the present study, which significantly overlap with those in our recent study [19] on sexual size dimorphism, with new data from more recent species descriptions added. We consider this set of species a random sample representing the Syringophilidae family as a whole.
Host species were identified by the host designated as the type host in the taxonomic literature. Their mean body size was quantified as the mean body mass (g) obtained from the literature [95]. We also gathered data for wing length (the longest primary, in mm) from [96] for the hosts of Syringophiline species but not for the Picobiines which infest contour feathers. Body mass data were lacking for some, and wing length data were lacking for several host species. All body length, body mass, and wing length data were log-transformed. Then, we applied Type-1 linear regressions to test whether host body size measures predict parasite body size.
3. Results
When including all species of the Syringophilidae family or narrowing it to the Syringophilinae subfamily, we obtained only non-significant negative trends for both the female and male parasites. By contrast, the length of Picobiinae species is related positively with host body mass, a significant relationship in males (Figure 1, Table 1).
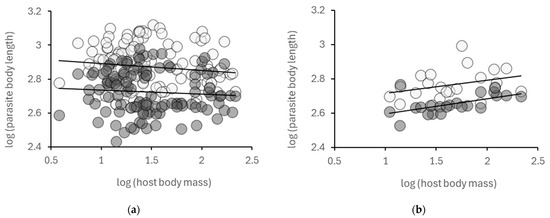
Figure 1.
(a) The relationship between host and parasite body sizes in the Syringophilidae family; (b) the relationship between host and parasite body sizes in the Picobiinae subfamily (a subset of the species of (a)). Note that the trendlines signify statistically non-significant tendencies (except for Picobiine males). (Light: females; dark: males).

Table 1.
Linear regressions between host body mass and parasite body length measures at the family and subfamily levels. * Indicates p < 0.05.
When analyzing all the parasite genera separately, as advised originally by Harrison [6], we documented mostly positive relationships between host and parasite body sizes. Eight quill mite genera were involved in the present study, one of which yielded an opposite result; both male and female Syringophiloidus mites exhibited non-significant negative relationships with host mass. The other seven genera showed positive host–parasite body size allometries, as predicted by HR, both for males and females, even though most of these relationships were statistically non-significant (Table 2). Considering only the directions of these relationships, the probability that the relationships of at least seven out of eight genera lead to the same direction is p = 0.0703 (like the probability of obtaining seven or eight identical results after tossing a coin eight times, from the binomial distribution). When considering the probability of obtaining at least seven-to−one in a predicted direction (predicted by HR), this probability is halved (p = 0.03515).
Table 2.
Linear regressions between host body size measures (body mass or wing length) and parasite body length at the genus level. * Indicates p < 0.05.
We do not know exactly the reason why Syringophiloidus exhibited a different (although non-significant) relationship to host body mass. However, since this genus inhabits the flight feathers (mostly secondaries, but sometimes also primaries) of host birds, we tested whether the size of these feathers would be more suitable indices of host body size. We found wing length (mm) data for an appropriate number of species (>6) in only three Syringophiline genera: Syringophiloidus, Syringophilopsis, and Torotrogla. We found a positive relationship between host wing length and parasite body length in all these genera, in both males and females, even though all six relationships were statistically non-significant (Figure 2, Table 2).
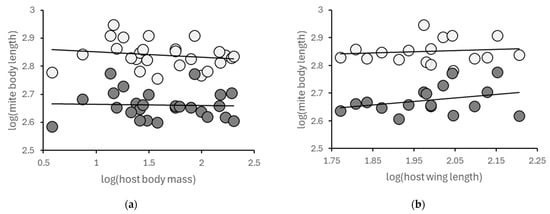
Figure 2.
In the case of Syringophiloidus spp., host body mass (a) appears to be an inferior index of host body size as compared to host wing length (b). This is plausible, considering that these mites mostly inhabit the host flight feathers (secondaries and also primaries). Note that the trendlines signify statistically non-significant tendencies. (Light: females; dark: males).
4. Discussion
Regarding the Syringophilid family as a whole, we see a negative relationship between host weight (g) and mite body length, contrary to HR. However, any scientific hypothesis can only be complete when its range of validity is determined. Harrison talked about comparisons across congeneric parasite species infecting similar hosts, even though subsequent authors often tested HR on taxa above the generic level with various results [7,8,10,11,12].
At the genus level, all genera, both males and females, exhibited a positive relationship between host mass and parasite length, except for the genus Syringophiloidus. In the case of three of these genera, host body size was also quantified as wing length, and all of them—including Syringophiloidus—obeyed HR. It is plausible to conclude that wing length is a better index of host body size than body mass for mites which inhabit the quills of wing flight feathers (mostly secondaries, but also primaries).
Generally speaking, the covariation of two animal traits should be analyzed by applying a phylogenetic control so as to separate phylogenetic artifacts (effects of conservative traits shared through common ancestry) from coordinated changes in the two traits occurring repeatedly in several independent cases along the phylogeny [97]. However, the present study involves a host trait which evolved along the host phylogeny and a parasite trait which evolved along the parasite phylogeny. Contrary to the classical hypotheses by Fahrenholz [98], host and parasite phylogenies are rarely similar and almost never identical [99], as was also shown for Syringophilopsis quill mites and their hosts [100]. Though it is theoretically possible to simultaneously control for the phylogenetic effects of both phylogenies [101], we chose a much simpler approach because we had too little information about the parasite phylogeny.
The method we used was to demonstrate that the relationship between the host and parasite traits repeats itself over and over again within each parasite genera, independently of each other. Thus, presuming that these genera are monophyletic taxa, our analysis corresponds to the basic logic of a phylogenetically controlled comparative analysis projected onto the parasite phylogeny.
5. Conclusions
Species descriptions usually attract rather few readers and a few citations. Our study above exemplifies an unusual utilization of such studies; we used species descriptions as the source of primary information on the body sizes of species. This approach is most appropriate for taxa where most species descriptions have been prepared by one author, or at least a small group of collaborating authors, using a standard preparation and measurement methodology, like in the case of quill mites.
Our results also indicate an unexpected reliability of the genus concept. Taxonomic ranks above the species level do not exist in nature, nor do they have any general definition. Despite that, Harrison declared that his observation was valid for genera but not necessarily for hierarchical levels above that. Although the genera we analyzed above were non-existent in Harrison’s age, his delineation of the range of validity is nicely supported by our present results.
The adaptive mechanism yielding the host–parasite body size allometry has yet to be fully understood. Host body size sets an upper limit on parasite body size, especially when parasites occupy narrowly space-limited anatomical structures. On the other hand, possible parasite body sizes also have a lower limit, which has rarely been considered [10]. Overall, it seems likely that host body size is typically constrained by environmental (like climatic, etc.) effects and phylogenetic constraints. In contrast, parasite body size tends to track that of the host within its own phylogenetic constraints [102].
Author Contributions
L.R., conceptualization, formal analysis, writing—review and editing, supervision; M.I., resources, data curation, writing—original draft preparation; M.H., writing—review and editing, supervision, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research, Development, and Innovation Fund of Hungary, grant number K143622; the National Research, Development, and Innovation Office of Hungary, grant number (RRF-2.3.1-21-2022-00006); the Agency of the Ministry of Education, Research and Sport of the Slovak Republic and the Slovak Academy of Sciences VEGA 1/0876/21; and by the Slovak Research and Development Agency under the contract APVV-22-0440.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Acknowledgments
We thank all the authors of the species descriptors on which the present study was based.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fenchel, T. Intrinsic rate of natural increase: The relationship with body size. Oecologia 1974, 14, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Hennemann, W.W. Relationship among body mass, metabolic rate and the intrinsic rate of natural increase in mammals. Oecologia 1983, 56, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Honěk, A. Intraspecific variation in body size and fecundity in insects: A general relationship. Oikos 1993, 66, 483–492. [Google Scholar] [CrossRef]
- Villa, S.M.; Evans, M.D.; Subhani, Y.K.; Altuna, J.C.; Bush, S.E.; Clayton, D.H. Body size and fecundity are correlated in feather lice (Phthiraptera: Ischnocera): Implications for Harrison’s rule. Ecol. Entomol. 2018, 43, 394–396. [Google Scholar] [CrossRef]
- Poulin, R. Evolutionary Ecology of Parasites; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Harrison, L. Mallophaga from Apteryx, and their significance; with a note on the genus Rallicola. Parasitology 1915, 8, 88–100. [Google Scholar] [CrossRef]
- Morand, S.; Legendre, P.; Gardner, S.L.; Hugot, J.P. Body size evolution of oxyurid (Nematoda) parasites: The role of hosts. Oecologia 1996, 107, 274–282. [Google Scholar] [CrossRef]
- Poulin, R.; Hamilton, W.J. Ecological correlates of body size and egg size in parasitic Ascothoracida and Rhizocephala (Crustacea). Acta Oecol. 1997, 18, 621–635. [Google Scholar] [CrossRef]
- Johnson, K.P.; Bush, S.E.; Clayton, D.H. Correlated evolution of host and parasite body size: Tests of Harrison’s rule using birds and lice. Evolution 2005, 59, 1744–1753. [Google Scholar] [CrossRef]
- Harnos, A.; Lang, Z.; Petras, D.; Bush, S.E.; Szabo, K.; Rózsa, L. Size matters for lice on birds: Coevolutionary allometry of host and parasite body size. Evolution 2017, 71, 421–431. [Google Scholar] [CrossRef]
- Maestri, R.; Fiedler, M.S.; Shenbrot, G.I.; Surkova, E.N.; Medvedev, S.G.; Khokhlova, I.S.; Krasnov, B.R. Harrison’s rule scales up to entire parasite assemblages but is determined by environmental factors. J. Anim. Ecol. 2020, 89, 2888–2895. [Google Scholar] [CrossRef]
- Ni, S.; de Angeli Dutra, D. Functional and sex-specific dynamics of ectoparasite size evolution in marine isopod–fish interactions: Harrison’s rule and increasing variance. Oecologia 2023, 201, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Gustafsson, D.R.; Tian, C.; Grossi, A.A.; Liu, Z.; Zou, F. Coevolutionary analysis of the Philopteroides Mey, 2004 (Phthiraptera: Ischnocera) parasitizing bulbuls (Passeriformes: Pycnonotidae). Front. Ecol. Evol. 2023, 11, 1053820. [Google Scholar] [CrossRef]
- Dabert, M.; Witalinski, W.; Kazmierski, A.; Olszanowski, Z.; Dabert, J. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and longbranch attraction artifacts. Mol. Phylogenet. Evol. 2010, 56, 222–241. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Fajfer, M.; Hromada, M.; Hušek, J.; Sikora, B. Tinamiphilopsis temmincki sp. n., a new Quill Mite species from Tataupa Tinamou, and the early history of Syringophilid mites. Animals 2022, 13, 2728. [Google Scholar] [CrossRef]
- Kethley, J.B. Population regulation in quill mites (Acarina: Syringophilidae). Ecology 1971, 52, 1113–1118. [Google Scholar] [CrossRef]
- Skoracki, M.; Bochkov, A.V.; Wauthy, G. Revision of the quill mites of the genus Picobia Haller, 1878 (Acari: Syringophilidae) with notes on their host-parasites relationships. Insect Syst. Evol. 2004, 35, 155–176. [Google Scholar] [CrossRef]
- Skoracki, M. Quill mites (Acari: Syringophilidae) of the Palaearctic region. Zootaxa 2011, 2840, 1–414. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B.; Jerzak, L.; Hromada, M. Tanopicobia gen. nov., a new genus of quill mites, its phylogenetic placement in the subfamily Picobiinae (Acariformes: Syringophilidae) and picobiine relationships with avian hosts. PLoS ONE 2020, 15, e0225982. [Google Scholar] [CrossRef]
- Casto, S.D. Entry and exit of syringophilid mites (Acarina: Syringophilidae) from the lumen of the quill. Wilson Bull. 1974, 86, 272–278. [Google Scholar]
- Skoracki, M.; Bochkov, A.V.; Zabludovskaya, S.A. A review of Prostigmata (Acariformes: Trombidiformes) permanently associated with birds. Acarina 2012, 20, 67–107. [Google Scholar]
- Glowska, E.; Dragun-Damian, A.; Dabert, M.; Gerth, M. New Wolbachia supergroups detected in quill mites (Acari: Syringophilidae). Infect. Genet. Evol. 2015, 30, 140–146. [Google Scholar] [CrossRef]
- Rózsa, L.; Moldovan, E. Relationship between body size and sexual size dimorphism in Syringophilid Quill Mites. Parasit. Res. 2022, 121, 891–898. [Google Scholar] [CrossRef]
- Bochkov, A.V. On the taxonomy of mites of the genus Syringophiloidus Kethley, 1970 (Acari: Syringophilidae) of European part of the Russia. Parasitologya 2001, 35, 149–153. [Google Scholar]
- Bochkov, A.V.; Apanaskevich, D. Two new species of the family Syringophilidae (Acari: Cheyletoidea) from passeriform birds collected in the Altai. Folia Parasit. 2001, 48, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Bochkov, A.V.; Galloway, T.D. Parasitic cheyletoid mites (Acari: Cheyletoidea) associated with passeriform birds (Aves: Passeriformes) in Canada. Can. J. Zool. 2001, 79, 2014–2028. [Google Scholar] [CrossRef]
- Bochkov, A.V.; Galloway, T.D. New species and records of cheyletoid mites (Acari: Cheyletoidea) from birds in Canada. J. Kansas Entomol. Soc. 2004, 77, 26–44. [Google Scholar] [CrossRef]
- Bochkov, A.V.; Mironov, S.V. Quill mites of the family Syringophilidae Lavoipierre, 1953 (Acariformes: Prostigmata) parasitic on birds (Aves) of the fauna of the former USSR. Acarina 1998, 6, 3–16. [Google Scholar]
- Bochkov, A.V.; Mironov, S.V. New quill mite species of the family Syringophilidae (Acari: Cheyletoidea) from the European part of Russia. Acarina 1999, 7, 35–45. [Google Scholar]
- Bochkov, A.V.; Fain, A.; Skoracki, M. New quill mites of the family Syringophilidae (Acari: Cheyletoidea). Syst. Parasitol. 2004, 57, 135–150. [Google Scholar] [CrossRef]
- Bochkov, A.V.; Flannery, M.E.; Spicer, G.S. Mites of the genus Torotrogla (Prostigmata: Syringophilidae) from North American passerines. J. Med. Entomol. 2009, 46, 183–197. [Google Scholar] [CrossRef]
- Bochkov, A.V.; Mironov, S.V.; Kravtsova, T. Two new Syringophilid mites from the Greenfinch Carduelis chloris (Passeriformes: Fringillidae) from Kirghizia (Acari: Syringophilidae). Genus 2000, 11, 351–358. [Google Scholar]
- Bochkov, A.V.; Mironov, S.V.; Skoracki, M. Four new Quill Mite species of the genus Syringophilopsis Kethley, 1970 (Acariformes: Prostigmata). Acarina 2001, 9, 233–239. [Google Scholar]
- Chirov, P.A.; Kravtsova, N.T. A new genus and new species of mites of the family Syringophilidae. Parazitologiya 1995, 29, 370–379. [Google Scholar]
- Fain, A.; Bochkov, A.V.; Mironov, S.V. New genera and species of quill mites of the family Syringophilidae (Acari: Prostigmata). Bull. Inst. Sci. Nat. Belg. Entomol. 2000, 70, 33–70. [Google Scholar]
- Glowska, E. New quill mites (Cheyletoidea: Syringophilidae) parasitizing tyrannid birds (Passeriformes: Tyrannidae) in Peru. Zootaxa 2014, 3814, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Glowska, E.; Skoracki, M. New species of quill mites (Acari, Cheyletoidea, Syringophilidae) and the first record of male for the genus Stibarokris. Zootaxa 2011, 2817, 63–68. [Google Scholar] [CrossRef]
- Glowska, E.; Laniecka, I.; Milensky, C.M. Two new picobiin mite species (Acari: Cheyletoidea: Syringophilidae) parasitizing passerine birds in Guyana. Acta Parasitol. 2015, 60, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Glowska, E.; Romanowska, K.; Schmidt, B.K.; Dabert, M. Combined description (morphology with DNA barcodedata) of a new quill mite Torotrogla paenae n. sp. (Acariformes: Syringophilidae) parasitising the Kalahari scrub-robin Cercotrichas paena (Smith) (Passeriformes: Muscicapidae) in Namibia. Syst. Parasitol. 2018, 95, 863–869. [Google Scholar] [CrossRef]
- Kethley, J.B. A revision of the family Syringophilidae (Prostigmata: Acarina). Contrib. Am. Entomol. Inst. 1970, 5, 1–76. [Google Scholar]
- Klimovičová, M.; Skoracki, M.; Njoroge, P.; Hromada, M. Two new species of the family Syringophilidae (Prostigmata: Syringophilidae) parasitising bushshrikes (Passeriformes: Malaconotidae). J. Parasitol. 2016, 102, 187–192. [Google Scholar] [CrossRef]
- Nattress, B.; Skoracki, M. Quill mites of the family Syringophilidae Lavoipierre, 1953 (Acari: Cheyletoidea) parasitic on birds in England. Genus 2000, 18, 139–145. [Google Scholar]
- Sikora, B.; Fajfer, M.; Skoracki, M. Quill mites (Acari: Syringophilidae) from mimid birds (Aves: Mimidae). Zootaxa 2011, 3027, 29–38. [Google Scholar] [CrossRef]
- Sikora, B.; Kaszewska, K.; Skoracki, M. Two new quill mites of the family Syringophilidae (Acari: Prostigmata) parasitising the tapaculos (Passeriformes: Rhinocriptidae) in South America. Zootaxa 2014, 3895, 419–426. [Google Scholar] [CrossRef]
- Sikora, B.; Unsöld, M.; Skoracki, M. Aulonastus paridus spec. nov.—A parasite of two bird species of the genus Melaniparus in Kenya and Tanzania. Spixiana 2016, 39, 149–152. [Google Scholar]
- Sikora, B.; Mahamoud-Issa, M.; Unsoeld, M.; Hromada, M.; Skoracki, M. Species composition of parasitic mites of the subfamily Picobiinae (Acariformes: Syringophilidae) associated with African Barbets (Piciformes: Lybiidae). Animals 2023, 13, 2007. [Google Scholar] [CrossRef]
- Sikora, B.; Unsoeld, M.; Melzer, R.R.; Friedrich, S.; Skoracki, M. First records of Picobiine mites associated with Birds-of-Paradise: Can interspecific sexual behaviour of hosts play a role in the distribution of Quill Mite parasites? Animals 2023, 13, 1509. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B.; Marciniak, N.; Zmudzinski, M. Syringophiloidus bucerotidus sp. nov. (Acari: Syringophilidae), a new quill mite species parasitizing hornbills (Aves: Bucerotidae) in the Sub-Saharan Africa. Int. J. Acarol. 2017, 43, 39–43. [Google Scholar] [CrossRef]
- Skoracki, M. New genus and species of Syringophilidae from Eurasian reed-warbler, Acrocephalus scirpaceus (Sylviidae: Passeriformes) (Acari: Prostigmata). Genus 1999, 10, 155–162. [Google Scholar]
- Skoracki, M. Three new species of quill mites of the genus Aulonastus Kethley, 1970 (Acari, Prostigmata, Syringophilidae) from passerine birds. Acta Parasitol. 2002, 47, 300–305. [Google Scholar]
- Skoracki, M. Three new species of the ectoparasitic mites of the genus Syringophiloidus Kethley, 1970 (Acari: Syringophilidae) from passeriform birds from Slovakia. Folia Parasit. 2002, 49, 305–313. [Google Scholar] [CrossRef]
- Skoracki, M. A review of quill mites of the genus Syringophiloidus Kethley, 1970 parasitizing quills of passeriform birds, with descriptions of four new species (Acari: Prostigmata: Syringophilidae). Genus 2004, 15, 281–300. [Google Scholar]
- Skoracki, M. Quill mites (Acariformes: Syringophilidae) associated with birds of Mexico. Zootaxa 2017, 4282, 179–191. [Google Scholar] [CrossRef]
- Skoracki, M.; Bochkov, A.V. Syringophilid mites from Kazakhstan. Zootaxa 2010, 2546, 52–68. [Google Scholar] [CrossRef]
- Skoracki, M.; Dabert, M. A new species of the genus Syringophilopsis Kethley, 1970 (Acari: Prostigmata: Syringophilidae) from the tree pipit Anthus trivialis (Passeriformes: Motacillidae). Acarina 1999, 7, 89–92. [Google Scholar]
- Skoracki, M.; Dabert, M. Syringophilopsis albicollisi sp.n., a new species of the quill mite of the family Syringophilidae (Acari: Prostigmata). Acarina 2000, 8, 59–63. [Google Scholar]
- Skoracki, M.; Dabert, M. The quill mites of the genus Syringophilopsis Kethley, 1970 (Acari: Syringophilidae) from African birds. Acarina 2001, 9, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Dabert, M. Two new species of the parasitic mites of the genus Aulobia Kethley, 1970 (Acari, Syringophilidae) from African birds. Acta Parasitol. 2001, 46, 208–215. [Google Scholar]
- Skoracki, M.; Dabert, M. A review of parasitic mites of the family Syringophilidae (Acari, Prostigmata) from African birds, with descriptions of four new species. Acta Parasitol. 2002, 47, 137–146. [Google Scholar]
- Skoracki, M.; Dabert, M.; Ehrnsberger, R. A new quill mite (Acari: Syringophilidae) from the blackbird. Osnabrucker Naturwissenschaftliche Mitteilungen 2000, 26, 191–198. [Google Scholar]
- Skoracki, M.; Flannery, M.E.; Spicer, G.S. Quill mites of the genus Syringophilopsis Kethley, 1970 (Acari: Syringophilidae) from North American birds. Folia Parasit. 2008, 55, 291–300. [Google Scholar] [CrossRef]
- Skoracki, M.; Glowska, E. Two new species of the quill mite genus Aulobia Kethley, 1970 (Acari, Syringophilidae) associated with sunbirds (Passeriformes, Nectariniidae). Acta Parasitol. 2008, 53, 179–185. [Google Scholar] [CrossRef]
- Skoracki, M.; Glowska, E. Two new species of the genus Picobia Haller (Acari: Syringophilidae) from Australian and Indonesian passeriform birds. N. Z. J. Zool. 2008, 35, 281–286. [Google Scholar] [CrossRef]
- Skoracki, M.; Glowska, E.; Sikora, B. Four new species of the quill mite genus Picobia Haller (Acari: Syringophilidae) parasitizing birds in the Australian region. Zootaxa 2008, 1961, 58–68. [Google Scholar] [CrossRef]
- Skoracki, M.; Hendricks, S.; Spicer, G.S. Systematics of the ectoparasitic quill mites of the genus Aulobia Kethley, 1970 (Acari: Syringophilidae) with the description of a new species. Zootaxa 2010, 2399, 31–41. [Google Scholar] [CrossRef]
- Skoracki, M.; Hendricks, S.; Spicer, G.S. Four new species of Aulonastus Kethley, 1970 (Acari: Syringophilidae) from North American passerines. Syst. Parasitol. 2010, 76, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Hendricks, S.; Spicer, G.S. New species of parasitic quill mites of the genus Picobia (Acari: Syringophilidae: Picobiinae) from North American birds. J. Med. Entomol. 2010, 47, 727–742. [Google Scholar] [CrossRef]
- Skoracki, M.; Hendricks, S.; Spicer, G.S. Systematics of the genus Syringophilopsis Kethley, 1970 (Acari: Prostigmata: Syringophilidae) with description of three new species from North American passerines. Zootaxa 2011, 2793, 1–22. [Google Scholar] [CrossRef]
- Skoracki, M.; Hromada, M. A review of picobiine mites (Acari: Syringophilidae: Picobiinae) parasitising African birds. Folia Parasit. 2013, 60, 192–212. [Google Scholar] [CrossRef]
- Skoracki, M.; Hromada, M.; Kuczynski, L. Torotrogla lullulae, a new species of the family Syringophilidae Lavoipierre, 1953 (Acari: Prostigmata: Cheyletoidea). Genus 2001, 12, 87–92. [Google Scholar]
- Skoracki, M.; Hromada, M.; Unsoeld, M. Three new quill mite species of the genus Neoaulonastus Skoracki (Acari: Syringophilidae) parasitizing passerines in Tanzania. Zootaxa 2013, 3616, 367–377. [Google Scholar] [CrossRef]
- Skoracki, M.; Klimovičová, M.; Muchai, M.; Hromada, M. New taxa of the family Syringophilidae (Acari: Prostigmata) from the African barbets and woodpeckers (Piciformes: Lybiidae, Picidae). Zootaxa 2014, 3768, 178–188. [Google Scholar] [CrossRef]
- Skoracki, M.; Magowski, W. Two new species of quill mites of the genus Picobia (Acari: Prostigmata: Syringophilidae) from passerine birds of Poland. Acarina 2001, 9, 113–119. [Google Scholar]
- Skoracki, M.; Mironov, S.V. New species and records of quill mites of the family Syringophilidae (Acari: Prostigmata) from the passerines (Aves: Passeriformes) from the Russian Far East. Zootaxa 2013, 3641, 554–564. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Skoracki, M.; Mironov, S.V.; Unsoeld, M. The first records of quill mites of the family Syringophilidae (Acariformes: Prostigmata: Cheyletoidea) from trogoniform birds (Aves: Trogoniformes). Zootaxa 2013, 3701, 291–297. [Google Scholar] [CrossRef]
- Skoracki, M.; Møller, A.P.; Tryjanowski, P. A new species of parasitic mites of the genus Syringophiloidus Kethley, 1970 (Acari: Syringophilidea) from the barn swallow Hirundo rustica Linnaeus, 1758. Parasite 2003, 10, 17–20. [Google Scholar] [CrossRef][Green Version]
- Skoracki, M.; OConnor, B.M.; Goodman, S.M.; Marciniak, N.; Sikora, B. New species and records of syringophilid mites (Acariformes: Syringophilidae) associated with Malagasy birds. Syst. Appl. Acarol. 2016, 21, 1534–1546. [Google Scholar] [CrossRef]
- Skoracki, M.; O’Connor, B. New taxa of quill mites (Acari: Cheyletoidea: Syringophilidae). Zootaxa 2010, 2341, 1–32. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B. Quill mites (Acari: Prostigmata: Syringophilidae) from African passeriform birds. Zootaxa 2003, 129, 1–10. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B. Two new quill mite species of the family Syringophilidae (Acari: Prostigmata) parasitising the house sparrow Passer domesticus (L.) (Aves: Passeriformes). Zootaxa 2014, 3765, 194–200. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B.; Spicer, G.S. A review of the subfamily Picobiinae Johnston and Kethley, 1973 (Acariformes: Prostigmata: Syringophilidae). Zootaxa 2016, 4113, 1–95. [Google Scholar] [CrossRef]
- Skoracki, M.; Solarczyk, P.; Sikora, B. Three new species of picobiine mites (Acari: Syringophilidae) parasitizing African flycatchers (Aves: Muscicapidae). Syst. Parasitol. 2012, 83, 123–135. [Google Scholar] [CrossRef]
- Skoracki, M.; Spicer, G.S.; OConnor, B.M. A systematic review of the subfamily Syringophilinae (Acari: Syringophilidae) of the Nearctic region. Part 1: Quill mites associated with passerines (Aves: Passeriformes). Zootaxa 2016, 4084, 451–494. [Google Scholar] [CrossRef]
- Skoracki, M.; Tryjanowski, P.; Hromada, M. Two new species of the genus Syringophilopsis Kethley, 1970 (Acari: Syringophilidae) parasitizing quills of true shrikes (Aves: Laniidae). Parasite 2002, 9, 11–16. [Google Scholar] [CrossRef][Green Version]
- Skoracki, M.; Unsoeld, M.; Kavetska, K.; Kaszewska, K. Quill mites of the subfamily Picobiinae (Acari: Syringophilidae) associated with woodpeckers (Aves: Piciformes: Picidae). Acta Parasitol. 2014, 59, 68–79. [Google Scholar] [CrossRef]
- Skoracki, M.; Glowska, E.; Lontkowski, J.; Stawarczyk, T. Picobia ictericus sp. n., an ectoparasite of two icterid bird species from Brazil (Acari: Prostigmata: Syringophilidae). Genus 2010, 21, 143–148. [Google Scholar]
- Skoracki, M.; Hromada, M.; Zmudzinski, M.; Unsoeld, M.; Sikora, B. Parasitic quill mites of the family Syringophilidae (Acariformes: Prostigmata) associated with sub-Saharan sunbirds (Passeriformes: Nectariniidae): Species composition and host-parasite relationships. J. Med. Entomol. 2018, 55, 1464–1477. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M. New Data on Systematics of the Quill Mites of the genus Torotrogla Kethley, 1970 (Acari, Syringophilidae). Belg. J. Entomol. 2004, 6, 303–314. [Google Scholar]
- Skoracki, M. Quill Mites of the genus Syringophilopsis (Acari, Syringophilidae) from passeriform birds of Poland with descriptions of five new species. Acta Parasitol. 2004, 49, 45–62. [Google Scholar]
- Skoracki, M.; Hromada, M.; Tryjanowski, P. Description of a new species of Quill Mite Syringophiloidus sp. n. (Acari, Prostigmata, Syringophilidae) from Great Grey Shrike Lanius excubitor. Acta Parasitol. 2001, 46, 30–34. [Google Scholar]
- Skoracki, M.; Spicer, G.S.; OConnor, B.M. A review of mites of the subfamily Picobiinae Johnston & Kethley, 1973 (Prostigmata: Syringophilidae) from North American birds. Syst. Parasitol. 2014, 87, 99–110. [Google Scholar] [PubMed]
- Skoracki, M.; Unsoeld, M.; Kosicki, J.Z.; Melzer, R.R.; Friedrich, S.; Sikora, B. Enigmatic host-mite relationships: Unraveling the distribution of quill mites on Birds-of-Paradise. Int. J. Parasitol. 2024, 54, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Unsöld, M.; Sikora, B. Redescription of Syringophilioidus glandarii (Fritsch, 1958) with new records of hosts and localities. Spixiana 2022, 45, 39–44. [Google Scholar]
- Skoracki, M.; Patan, M.; Unsoeld, M.; Hromada, M.; Kwiecinski, Z.; Marcisova, I. Diversity of Quill Mites of the family Syringophilidae (Acariformes: Prostigmata) parasitizing Starlings of the genus Lamprotornis (Passeriformes: Sturnidae). Diversity 2024, 16, 51. [Google Scholar] [CrossRef]
- Dunning, J.B. CRC Handbook of Avian Body Masses, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Featherbase. Available online: https://www.featherbase.info/en/home (accessed on 30 June 2024).
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Fahrenholz, H. Ectoparasiten und Abstammungslehre. Zool. Anz. 1913, 41, 371–374. [Google Scholar]
- Page, R.D. (Ed.) Tangled Trees: Phylogeny, Cospeciation, and Coevolution; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Hendricks, S.A.; Flannery, M.E.; Spicer, G.S. Cophylogeny of Quill Mites from the genus Syringophilopsis (Acari: Syringophilidae) and their North American passerine hosts. J. Parasitol. 2013, 99, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.D.; Krasnov, B.R.; Poulin, R.; Nakagawa, S. A tale of two phylogenies: Comparative analyses of ecological interactions. Am. Nat. 2014, 183, 174–187. [Google Scholar] [CrossRef]
- Poulin, R. Functional biogeography of parasite traits: Hypotheses and evidence. Philos. Trans. R. Soc. B 2021, 376, 20200365. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).