Abstract
Urbanization is one of the leading causes of habitat loss, which has increased significantly in tropical regions in recent years, leading to the loss of species, their ecological functions, and evolutionary history. To determine the effect of urbanization on the diversity of bat communities in urban and suburban environments, we analyzed the α and β taxonomic, functional, and phylogenetic diversities at four sites along urbanization gradients surrounding a rapidly expanding city (Oaxaca City) in southern Mexico. We recorded bats using conventional techniques such as mist nets and acoustic monitoring. We calculated the diversity of bats in four sites with different urbanization conditions: urban (1), suburban (1), and rural (2). To assess the degree of total differentiation and components of bat turnover and nestedness between sites, we calculated the β taxonomic, functional, and phylogenetic diversities. A total of 33 bat species were recorded. The highest taxonomic and functional diversity was observed in the Center of Oaxaca (the site with the highest level of urbanization). In contrast, the highest phylogenetic diversity was found in the West (the site with the lowest level of urbanization). The total β taxonomic diversity was higher than the functional and phylogenetic diversity. Regarding the contributions of turnover and nestedness, turnover made a more significant contribution than nestedness to the taxonomic and phylogenetic β diversity. In contrast, functional nestedness contributed more to the functional β diversity than turnover. Tadarida brasiliensis, Desmodus rotundus, Sturnira hondurensis, and S. parvidens were recorded in all three urbanization conditions. In the most urbanized site, four Myotis species were recorded: M. fortidens, M. keaysi, M. thysanodes, and M. velifer. We suggest that the analysis of different dimensions of diversity is essential and should be considered to strengthen conservation strategies; moreover, we suggest the preservation of native vegetation mosaics and water bodies within the city to maintain bat diversity.
1. Introduction
Human population growth has increased exponentially in recent years, leading medium-sized cities to expand into large cities with over a million inhabitants [1]. Consequently, rural or semi-rural areas surrounding these cities have transformed into urban zones [2]. The urbanization process alters landscapes, causing habitat loss and fragmentation. Human activity is considered one of the greatest threats to global biodiversity [3].
In urban landscapes, biological communities interact in response to factors such as city size, climatic context, availability of green spaces, and proximity to native vegetation [4]. Among the most studied biological groups affected by urbanization are flying vertebrates, including bats, where it has been observed that their species richness and abundance decrease in sites with varying degrees of urbanization [5]. However, it has also been noted that results vary by species, as some can adapt to or benefit from some urban regions [1,6].
Bats are a highly diverse and abundant group with a relatively well-known natural and taxonomic history. Additionally, they exhibit a wide variety of diets, feeding strategies, body sizes, flight capabilities, and degrees of habitat specialization [7,8]. Due to these characteristics, bats perform several critical ecological functions, making them an excellent group for studying the impact of urbanization on species diversity.
One of the rapidly growing cities in southern Mexico is Oaxaca de Juárez, which is in the state of Oaxaca. As of 2020, it had an approximate population of four million inhabitants. It is also one of the most visited sites in Mexico by national and international tourists. Within the urban area of Oaxaca de Juárez, 33 bat species have been identified [9], representing 34% of the species found in the state of Oaxaca. However, urban growth may be affecting the presence of these species, making it essential to monitor these organisms continually.
Alpha (α) diversity (species richness) and beta (β) diversity (differences in species composition between communities) are among the most used biodiversity metrics. However, these metrics do not consider the ecological characteristics and evolutionary history of species [10,11]. Recently, various studies have emphasized the importance of considering different facets of communities to better characterize biodiversity, especially in urban areas [12,13,14]. This is because species occupying the same habitats may exhibit different levels of phylogenetic relationships and functional traits [15,16].
Functional diversity has received attention in recent years, as it reflects the variation among species in ecological attributes associated with how they respond to environmental variation and their effects on ecosystem processes and services [17,18]. On the other hand, phylogenetic diversity quantifies evolutionary diversity within communities, describing the evolutionary heritage or relationship of all species in each community [19,20].
Understanding the impact of native ecosystem conversion on bat diversity in cities like Oaxaca is crucial because bats provide a wide range of ecosystem services. Therefore, due to the city’s rapid expansion with suburban areas it is necessary to analyze the effect of urbanization on wildlife, particularly bats. The main objective of this study is to evaluate the α and β taxonomic, functional, and phylogenetic diversities of bat communities at sites with different urbanization gradients in Oaxaca de Juárez and surrounding suburban cities, providing new insights into bat conservation in urban and suburban areas in southern Mexico. We tested whether sites with different urbanization gradients (human population density, green areas, light pollution, and impervious surfaces) affect bat communities’ taxonomic, functional, and phylogenetic diversities.
We expect that sites with high urbanization levels will exhibit lower diversity (species richness, functional, and phylogenetic diversity) while sites with lower urbanization levels will exhibit higher diversity (species richness, functional, and phylogenetic diversity).
This study aims to provide information on how landscape fragmentation in urban areas affects bat communities in the Neotropical region. Moreover, the key mechanisms shaping bat community structure in urbanized sites can be identified by analyzing diversity from a multidimensional perspective (taxonomic, functional, and phylogenetic) in response to habitat conversion. The results of this study can inform conservation policies and habitat management in the Neotropical region to protect biological diversity and promote species coexistence in landscapes altered by human activities.
2. Materials and Methods
2.1. Study Area
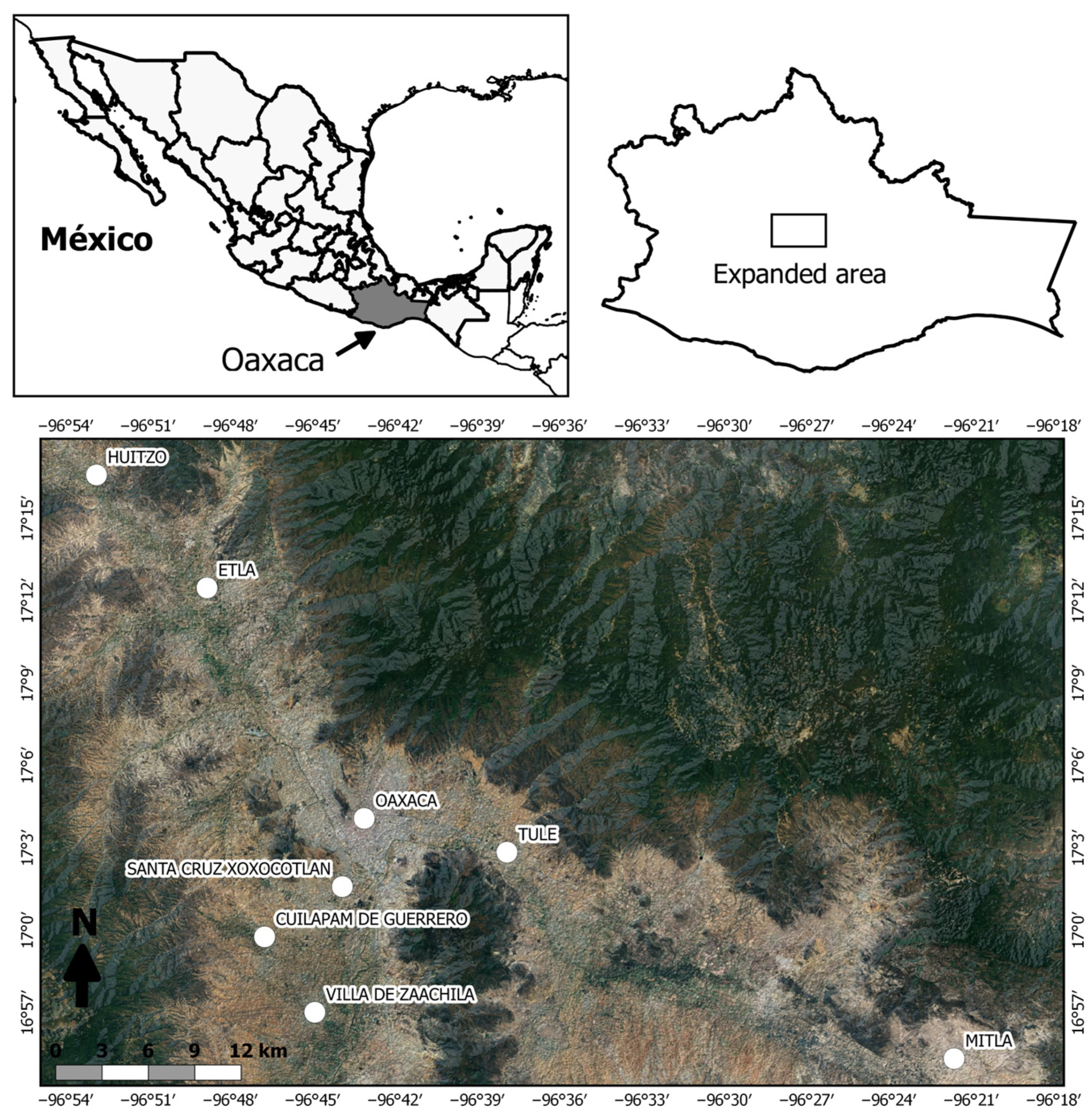
In southern Mexico, within the state of Oaxaca, lies the physiographic subprovince known as the Central Valleys of Oaxaca, located in the central portion of the state (Figure 1). This region is characterized by a stepped system of plains [21], covering an area of 2267.42 km2, with diverse types of vegetation such as tropical deciduous forests, xerophilous scrub, oak forests, pine forests, and pine–oak forests [22]. The climate ranges from arid to warm subhumid, with an annual mean temperature of 18 to 22 °C and an average annual precipitation of 500 to 800 mm [23].
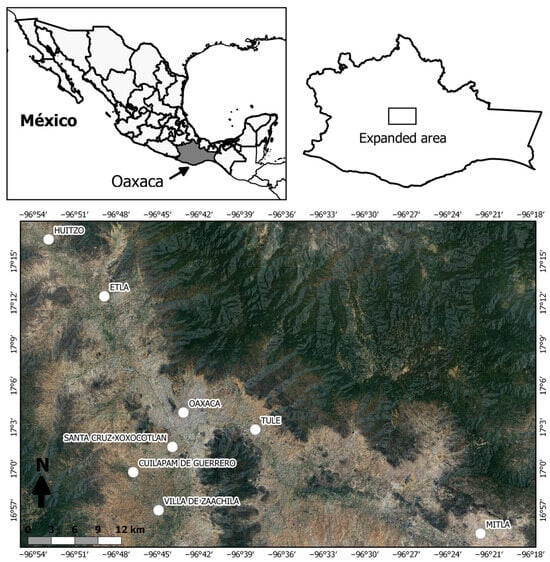
Figure 1.
Geographic location of the study area in the Central Valleys of Oaxaca. The analyzed localities are indicated.
For the study, the material collection places were divided into four sites according to urbanization characteristics. The sites were named a priori according to their geographic location, considering Oaxaca City as the central site: (1) Central Site (Urban Site: Oaxaca City and Santa Cruz Xoxocotlán), (2) South Site (Semi-Urban Site: Cuilápam de Guerrero and Villa de Zaachila), (3) East Site (Rural Site: Santa María del Tule and San Pablo Villa de Mitla), and (4) West Site (Rural Site: San Pablo Huitzo and Villa de Etla) (Figure 1, Table 1).

Table 1.
Urbanization characteristics of the four analyzed sites in the Central Valleys of Oaxaca, Mexico.
Central Site; Oaxaca City: The total area is 86.69 km2, with an average altitude of 1550 m and an annual mean temperature of 22 °C. Land use is distributed as follows: 25.9% agriculture, 0.9% pasture, 34.9% secondary vegetation, and 38.3% urban areas [24,25]. Santa Cruz Xoxocotlán: The total area is 48.09 km2, with an annual mean temperature of 20.4 °C and an average altitude of 1530 m. Land use is as follows: 63.79% agriculture and 20.27% pasture [24,25].
South Site; Cuilápam de Guerrero: The total area is 48.66 km2, with an annual mean temperature of 20.4 °C and an average altitude of 1560 m. Land use is distributed as follows: 58.98% agriculture, 17.42% induced pasture, 6.97% tropical deciduous forest, and 16.63% urban area [24,25]. Villa de Zaachila: The total area is 81.69 km2, with an annual mean temperature of 20 °C and an average altitude of 1552 m. Land use is as follows: 62.95% agriculture, 23.03% pasture, 11.42% tropical deciduous forest, 2.60% bare soil, and 12.79% urban area [24,25].
East Site; Santa María del Tule: The total area is 25.52 km2, with a temperate climate ranging from 18 to 22 °C and an average altitude of 1560 m. Land use is distributed as follows: 58.61% agriculture, 8.28% induced pasture, and 33.11% urban area [24,25]. San Pablo Villa de Mitla: The total area is 82.93 km2, with temperatures ranging from 14 to 22 °C and an altitude between 1300 and 2800 m. Land use is as follows: 20.93% agriculture, 2.46% urban area, 60.11% tropical deciduous forest, and 16.50% induced pasture [24,25].
West Site; San Pablo Huitzo: The total area is 63.79 km2, with an elevation of 1700 m and an average temperature of 26 °C. Land use is distributed as follows: 9.70% agriculture, 7.14% urban area, 59.9% coniferous and oak forest, and 23.26% induced pasture [24,25]. Villa de Etla: The total area is 8.47 km2, with a semi-warm subhumid climate and temperatures ranging from 18 to 20 °C. The elevation is 1660 m. Land use is as follows: 55.92% agriculture, 18.73% urban area, and 25.35% induced pasture [24,25].
2.2. Data Collection
In the four sites, monthly visits were conducted from January to August 2018, with two nights per site. The sampling covered the dry season (January–April) and the rainy season (May–September). Two techniques were employed for bat recording: mist net captures and acoustic monitoring.
For bat capture, six mist nets sized 12 × 2.6 m with a mesh opening of 2 × 2 cm were used. The nets were placed near trees, paths, roads, urban parks, and buildings, avoiding full moon days. The nets were open from twilight until 03:00 h and checked hourly. Captured bats were measured for total length, forearm length, ear length, leg length, and tail length. Age was determined based on phalangeal ossification, and reproductive condition in males was assessed by testicle position (scrotal for active, inguinal for inactive); females were evaluated for pregnancy, lactation, or post-lactation by palpation of the abdomen [26]. Taxonomic identification followed identification guides [27], and the taxonomic arrangement was based on Ramírez-Pulido et al. [28]. Capture was conducted under permit SGPA/DGVS/07292/17 from the Ministry of Environment and Natural Resources. Specimens were handled following guidelines from the American Society of Mammologists (ASM) for wildlife research. Most specimens were released at capture sites, while others were voucher specimens deposited in the Mammalogy Collection of the Interdisciplinary Research Center for Integral Regional Development, Oaxaca Unit, National Polytechnic Institute (OAX.MA.026.0497).
Acoustic monitoring sessions were conducted using passive recording to comprehensively record species presence using the Avisoft Ultrasound Gate116 Hm detector. This detector was active for ten minutes each hour from 18:00 to 03:00 h, set to high sensitivity, and recordings were made under favorable environmental conditions (rain-free nights with minimal wind). Recordings were analyzed using BatSound (version 4.2), with vocalizations separated for identification using spectrograms from García-Luis and Briones-Salas [9] and Ortega et al. [29].
2.3. Selection of Functional Traits
A functional trait is a measurable property that strongly influences an organism’s performance [30] and is linked to ecosystem processes, ecosystem stability, biological interactions, and/or habitat modification [31]. Ecological and morphological functional traits were selected for 33 bat species, as bat morphology relates to movement capacity, diet, and habitat use, while ecological traits represent species requirements and relate to ecosystem services [15,32,33]. The functional trait values were sourced from Martin-Regalado and Briones-Salas [34] (Table 2, Supplementary Material S1).
Table 2.
Functional traits used to quantify the functional diversity of bats in urban and suburban environments in the Central Valleys of Oaxaca.
2.4. Calculation of α Taxonomic, Functional, and Phylogenetic Diversities
Taxonomic diversity was obtained by considering species richness, i.e., the number of bat species recorded at each site in this study.
For functional diversity, the functional diversity index (FD) by Petchey and Gaston [18,35] was calculated. This index measures the total branch length connecting all species in a functional dendrogram (a multivariate classification analysis based on functional traits). Gower distance was used, recommended for datasets containing both qualitative and quantitative data. The Unweighted Pair Group Method with Arithmetic Mean (UPGMA) was employed to create the distance matrix and construct the dendrogram. The FD index typically correlates positively with species richness. To account for this, a null model approach [36] was used to standardize FD and obtain functional diversity appropriately. The null model randomly selected species but maintained species richness for each site. The Standardized Effect Size (SES) of FD was calculated using the formula: SES.FD = (Meanobs−Meannull)/sdnull, where Meanobs is the mean of observed measurements, Meannull is the mean of 99 iterations under the null model, and sdnull is the standard deviation of null model iterations. Analyses were conducted in R (version 3.3.3) using the FD package [37] and vegan package [38].
Phylogenetic diversity was estimated for each site using the index proposed by Faith [39,40]. This index measures the total branch lengths of a phylogenetic tree and quantifies phylogenetic richness as the total amount of evolutionary history among all species at each site. Phylogeny from Upham et al. [41] was used for calculations (Supplementary Material S2). Some indices of phylogenetic diversity show a positive correlation with species richness. To counteract this effect, null models were performed, and Standardized Effect Size of Phylogenetic Diversity (SES.PD) was calculated similarly to SES.FD. SES.PD was calculated using the “independent swap” null model, which randomizes the data matrix for each site while maintaining species. Analyses were conducted in R (version 3.3.3) using the picante package [42].
2.5. Urbanization vs. Taxonomic, Functional, and Phylogenetic Diversities
To assess the potential relationships between the levels of urbanization and taxonomic, functional, and phylogenetic diversities of bat communities, correlations were conducted between these variables (urbanization vs. diversity). Urbanization values at the four study sites were categorized a priori into three classes (9, 6, and 3; Table 1), based on the following characteristics: percentage of asphalted areas, percentage of green areas, population, housing units, and urbanization level, where 9 represents the most urbanized site and 3 the least urbanized.
2.6. Calculating Taxonomic, Functional, and Phylogenetic β Diversities
To determine the degree of total differentiation and the components of turnover and nestedness of bats among four sites (Center, South, East, and West), according to the Jaccard index [43], we calculated the following: (1) total taxonomic β diversity (Taxoβjaccard); (2) taxonomic β diversity due to turnover (Taxoβjturnover); (3) taxonomic β density due to nestedness (Taxoβjnestedness); (4) total functional β diversity (Funcβjaccard); (5) functional β diversity due to turnover (Funβjturnover); (6) functional β diversity due to nestedness (Funβnestedness), (7) total phylogenetic β diversity (Phyloβjaccard); (8) phylogenetic β diversity due to turnover (Phyloβjturnover); and (9) phylogenetic β diversity due to nestedness (Phyloβjnestedness). Taxonomic, functional, and phylogenetic β diversities were calculated using the function beta.multi for multiples samples and beta.pair for pairwise values. Functional β diversity refers to the variation in functional traits among species from two or more sites [44], and in this study it is based on the volume of convex hull intersections in a multidimensional functional space using four quantitative functional traits (total length, tail length, ear length, and weight). On the other hand, phylogenetic or evolutionary β diversity (phylobeta) measures the phylogenetic distance or dissimilarity between sites (i.e., how phylogenetic relationships change over space or time) [45], and in this study are based on Faith’s phylogenetic diversity [40].
To graphically represent the dissimilarity between pairs of sites in terms of their taxonomic β diversity, functional β diversity, and phylogenetic β diversity, we conducted a cluster analysis based on UPGMA. All analyses were conducted using the “betapart” package [43] in R (version 3.3.3).
3. Results
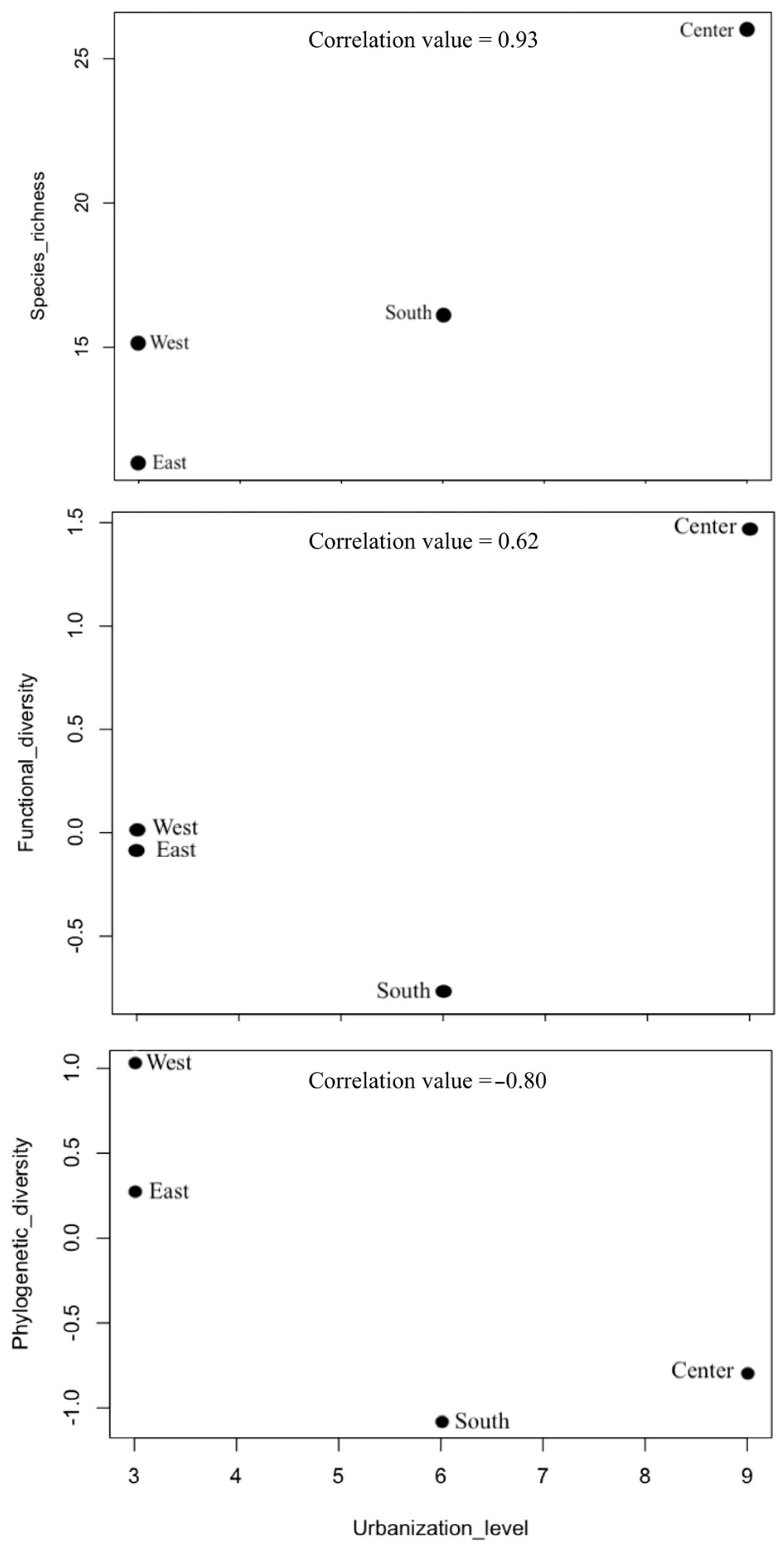
For the entire region, we recorded 33 species of bats. Species richness was highest at the Central Site of Oaxaca with 26 species, followed by the South, West, and East with 16, 15, and 11 species, respectively (Table 3, Figure 2). Functional diversity was highest at the Central Site (1.461), followed by the West and East; the South exhibited the lowest functional diversity (−0.787). Phylogenetic diversity was highest at the West Site (1.02), followed by the East and Central Sites, while the South had the lowest phylogenetic diversity (−1.09) (Figure 2).
Table 3.
Bats species recorded at the four sites in the Central Valleys of Oaxaca. Sites: Center (Oaxaca City and Santa Cruz Xoxocotlán), South (Cuilápam de Guerrero and Villa de Zaachila), East (Santa María el Tule and San Pablo Villa de Mitla), West (San Pablo Huitzo and Villa de Etla). MX = Endemic to Mexico.
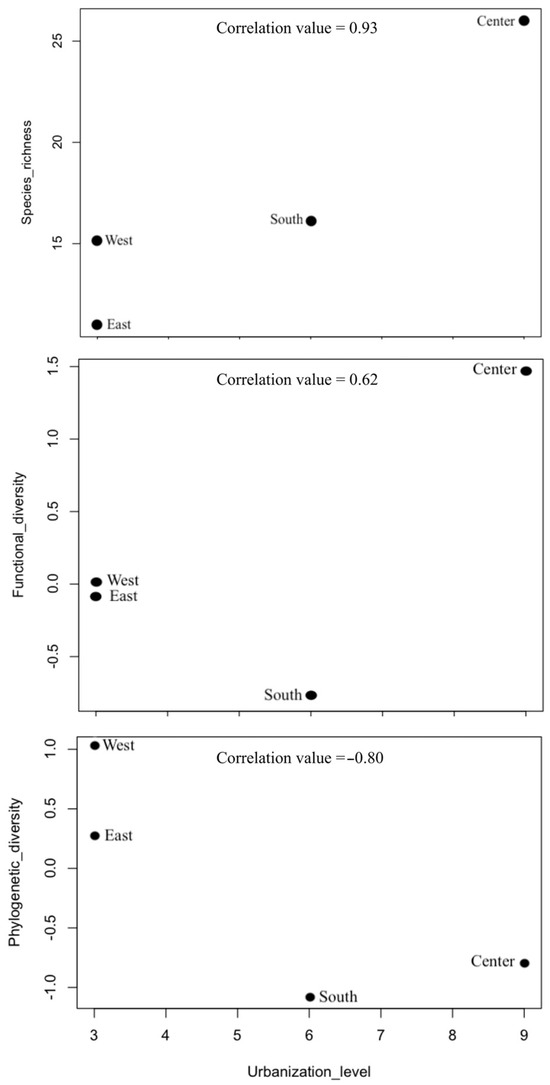
Figure 2.
Relationship between α taxonomic, functional, and phylogenetic diversities and the level of urbanization at four sites in the Central Valleys of Oaxaca, Mexico.
We found a high positive correlation between urbanization and bat species richness (0.93), while functional diversity and urbanization showed a moderate positive correlation (0.62). Finally, phylogenetic diversity and urbanization exhibited a high negative correlation (−0.80) (Figure 2).
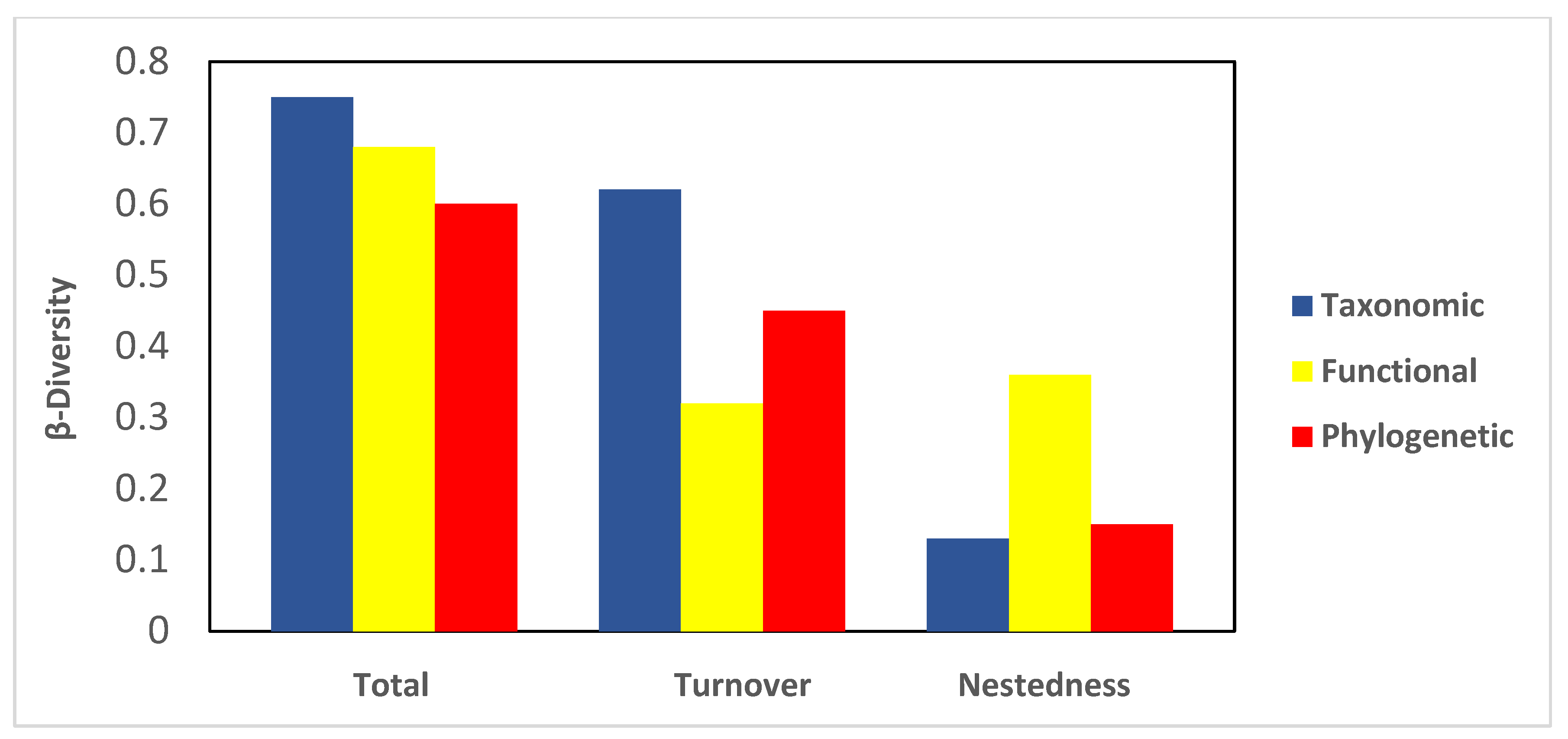
Total taxonomic β diversity (0.75) was higher than functional (0.68) and phylogenetic diversity (0.60) (Figure 3). Regarding the contributions of turnover and nestedness, turnover made a more significant contribution (0.62) than nestedness to taxonomic β diversity; a similar pattern was observed for phylogenetic β diversity (0.45). In contrast, functional nestedness contributed more (0.36) to functional β diversity than turnover (Figure 3).
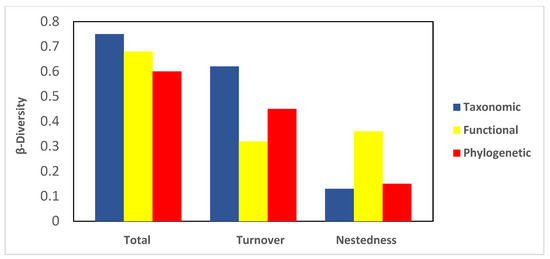
Figure 3.
Phylogenetic, functional, and taxonomic bats β diversities (i.e., total, turnover, and nestedness) in four sites in the Central Valleys of Oaxaca, México.
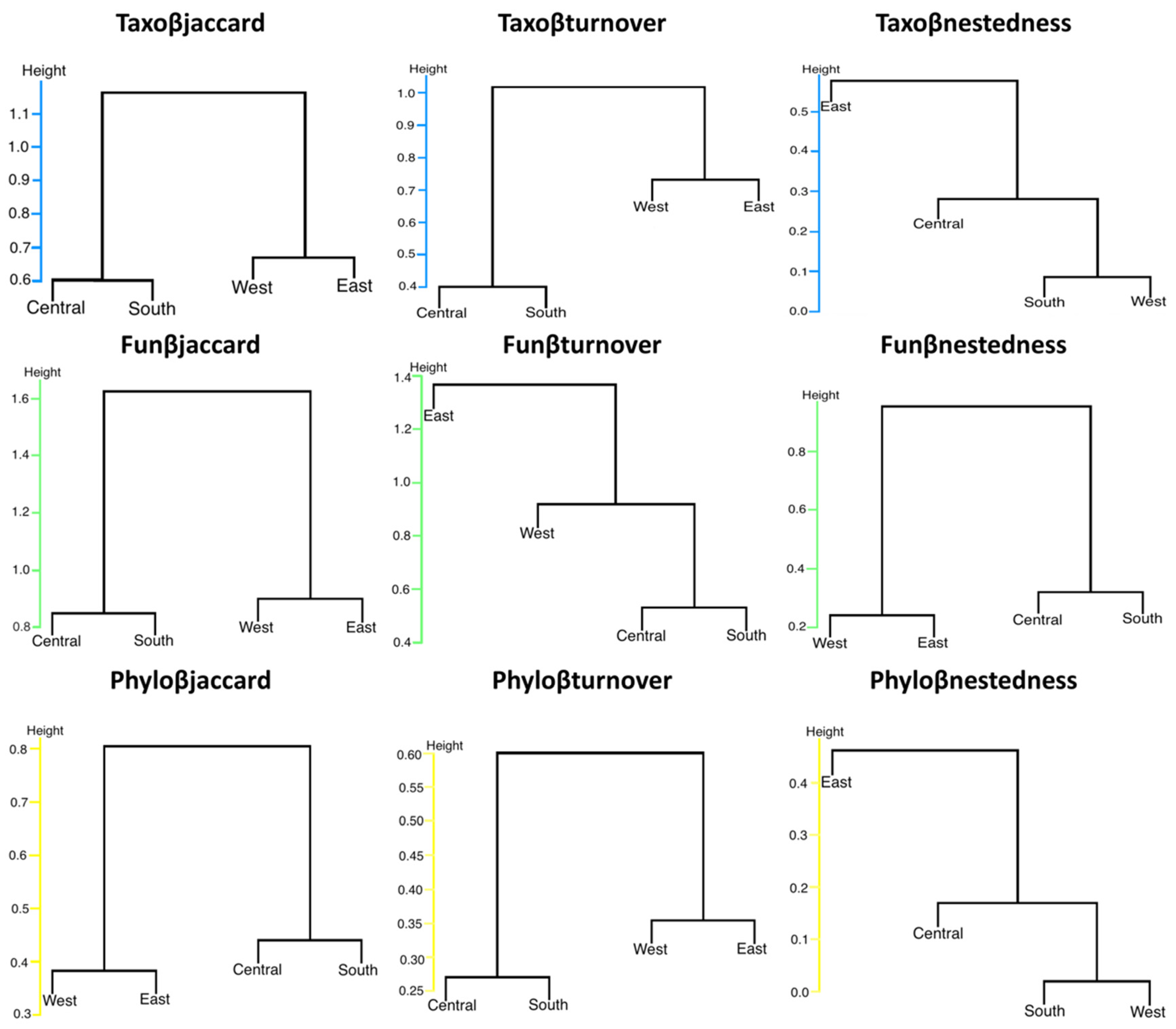
Cluster analyses using UPGMA based on taxonomic dissimilarity (Taxoβjaccard), functional dissimilarity (Funβjaccard), and phylogenetic dissimilarity (Phyloβjaccard) of bats in urban and suburban environments in southern Mexico formed the same groups (Central–South and West–East) based on their taxonomic, functional, and phylogenetic composition. However, for the turnover component of species, Taxoβturnover and Phyloβturnover showed similar clustering patterns. In contrast, Funβturnover exhibited a different pattern where the East Site did not cluster with the other sites. For the nestedness component, Taxoβnestedness and Phyloβnestedness showed similar clustering patterns, whereas Funβnestedness grouped differently (Figure 4).
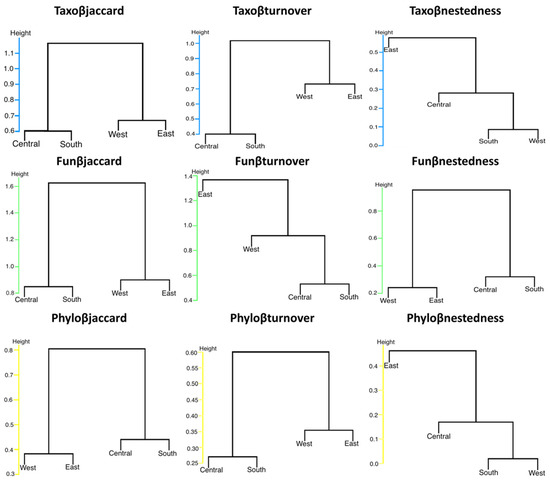
Figure 4.
UPGMA cluster analysis. The groups represent total taxonomic diversity (Taxoβjaccard), functional diversity (Funβjaccard), and phylogenetic diversity (Phyloβjaccard) of bats at four sites in the Central Valleys of Oaxaca, Mexico, based on Jaccard dissimilarity index and its turnover components (Taxoβturnover, Funβturnover, Phyloβturnover) and nestedness components (Taxoβnestedness, Funβnestedness, Phyloβnestedness).
4. Discussion
Some studies have demonstrated that cities are not favorable habitats for certain bat species, with many being unable to tolerate urban environments [46]. These studies suggest that urbanization can generally be detrimental to the diversity and abundance of some aerial insectivore bat species [47,48]. However, we recorded a high number of species, nearly 34% of the bat species reported for the State of Oaxaca (97 species; [49]), and almost double the number found in other studies on bat diversity in cities across Mexico (e.g., [5,50,51]).
The predominant species in our study belong to insectivore bats of the Family Vespertilionidae and frugivore bats of the Family Phyllostomidae, which are commonly found in urban areas [46] and appear to benefit from resources available in urban and suburban environments [52]. The variety of guilds and the high number of bat species recorded in the Central Valleys of Oaxaca may be attributed to the diverse habitats in the region (e.g., gardens, parks, riparian vegetation, agave crops), which provide adequate resources for bats.
Among the analyzed sites, the highest number of bat species was found in the Central Site, which has the largest territorial extent and the highest percentage of urbanization (70% of asphalt-covered areas; Table 1, Figure 2). This contrasts with our predictions and the findings reported by other authors such as Gutiérrez-Granados and Rodríguez-Zuñiga [5] in a study conducted in Mexico City. However, in some European sites, greater species richness and changes in species-specific activity levels were observed relatively close to the urban center in the transition between urban and peri-urban areas [53].
A wide range of bats in the site with the highest percentage of urbanization could be related to the availability of food resources (e.g., fruits, flower nectar, and insects). Saldaña-Vázquez et al. [54] mention that bats inhabit urban ecosystems not only due to year-round availability of food but also due to the abundance of roosting sites found in cities (e.g., abandoned buildings, sewers, bridges, and large trees). Additionally, it has been documented that habitat complexity in urban areas positively influences bat species richness and foraging activity [5]. Another factor that may affect species richness at these sites is the proximity of mountains to the north of the city, where Benito Juárez National Park is just a few kilometers away, providing a variety of roosting sites and a foraging habitat. Some species, such as Myotis or Mormoops megalophylla, may feed in urban sites and roost in the forests of the adjacent mountains [55]. In this context, it becomes clear that urban-sensitive species decrease while urban-tolerant species, or synurbic species, increase. We recorded only two species at sites with lower urbanization, Pteronotus parnelli and Rhogeessa gracilis, both at the West Site, which could be considered urban-sensitive. In contrast, six species were exclusive to the highly urbanized Central Site (M. megalophylla, D. azteca, M. fortidens, M. keaysi, M. thysanodes, and M. velifer), which could be considered synurbic [1].
In contrast, the East Site, corresponding to a rural area (28% asphalt-covered areas; Table 1, Figure 2), had the lowest number of bat species. This may be attributed to the homogeneity of the landscape and the limited availability of food. Most of the bats recorded at this site are frugivores (e.g., Artibeus lituratus and Sturnira parvidens), likely due to the abundance of cultivated (e.g., guavas, zapotes, mangoes, and plums) and native fruit trees, which serve as the food sources for these species. These animals contribute to the maintaining and restoring of spaces within urban fragments through seed dispersal [46].
Functional diversity varied among sites and was significantly higher in the Central Site, suggesting that bat species in this urban area do not share many functional traits. Interestingly, four species of the genus Myotis were recorded in this highly urbanized site. Despite sharing similar diets and litter sizes, these species exhibit different morphological measures (e.g., total length and ear length). A study revealed significant associations between morphological characteristics and feeding styles within this group of organisms [56]. For instance, aerial feeders like M. velifer and M. fortidens, which feed on moths, flying ants, and small beetles, have tiny hind feet, short calcars, short ears, and narrow tragi [56]. Moreover, these species typically roost in caves, tunnels, and abandoned structures often found in urban and suburban areas. The functional traits of these dominant species provide essential ecosystem services, such as pest control and disease vector regulation [57]. This underscores the importance of incorporating ecological and morphological traits in functional diversity studies [58].
The lowest functional diversity was observed at the South Site (Cuilápam de Guerrero and Villa de Zaachila), a semi-urban area (51% asphalt-covered areas; Table 1), where several frugivore species of the genera Artibeus and Sturnira are present. This suggests significant ecological redundancy in the area, resulting in lower functional diversity, likely due to the abundant availability of fruit resources.
The highest phylogenetic diversity was found at the West Site (San Pablo Huitzo and Villa de Etla), with the smallest territorial extent and lowest percentage of urbanization (12% asphalt-covered areas; Table 1, Figure 2). These findings indicate that the 15 bat species found at this site are more distantly related (i.e., they exhibit greater phylogenetic distances) than those found at other sites. Species such as Leptonycteris nivalis, Pteronotus parnellii, and Rhogeessa gracilis were recorded exclusively at the West Site, potentially providing unique evolutionary information for that location. These species exhibit different feeding habits; for example, L. nivalis feeds on pollen and nectar, probably from the agaves that exist at this site and are used for mezcal production. In contrast, P. parnelli and R. gracilis feed on arthropods at low heights, primarily Lepidoptera and Coleoptera. Gordon et al. [59] mention that the current diversity of foraging strategies in bats, which includes both aerial insectivores and foragers, suggests that different bat lineages have developed specializations in response to their particular ecological context.
Conversely, the South Site exhibited the lowest phylogenetic diversity, suggesting that some bat species in this urban site share similar evolutionary histories (i.e., closely related), which may increase their vulnerability to diseases and environmental changes. These phylogenetic results reflect not only the evolutionary history, genetic diversity, and ecosystem functioning but also how increased urbanization can reduce phylogenetic richness, which sometimes parallels species loss [39,60,61,62]. This loss could be concerning if the most vulnerable species are evolutionary distinct or unique. Under this scenario, the homogenization of the physical environment (i.e., more urbanized sites) may be creating filters that determine which lineages can persist in these environments, leading to phylogenetic clustering and low phylogenetic diversity (i.e., phylogenetic homogenization) [63], which would reduce the ability of bat communities to cope with environmental change. Our results confirm this expectation, revealing that bat phylogenetic diversity was lower in more urbanized sites (South and Central).
We recorded four species shared across four sites: Tadarida brasiliensis, Desmodus rotundus, Sturnira hondurensis, and S. parvidens. The genus Sturnira appears to be highly ecologically plastic, capable of feeding on both wild and cultivated plants, as mentioned in previous studies in this region of Oaxaca [64]. Their ability to exploit a wide variety of fruits enables them to adapt to urbanization and maintain healthy populations [65]. On the other hand, T. brasiliensis, with their high dispersal capability, can travel long distances in search food, and are notable for their ability to adapt to urban environments, influenced by their foraging behavior and ability to adjust to altered landscapes [52,66,67]. The presence of insectivore bats such as T. brasiliensis and Myotis is significant because their diet can include insect pests, providing an ecosystem service with economic benefits [68]. Only one sanguinivore species was recorded, D. rotundus, which has been noted to find roosts in caves in the Sierra Norte of Oaxaca [69] and descend to feed on goats, sheep, and cattle in the Central Valleys of Oaxaca. However, this species did not appear to pose a problem for the livestock in the visited localities (pers. obser.)
In contrast, six exclusive species, five of which are arthropodivore, were recorded at the site with the highest urbanization. Four exclusive species were recorded at the South Site (semi-urban). The opposite was observed at the rural sites, with three species exclusive to the West Site, while no exclusive species were present at the East Site. These findings contradict our predictions of the highest species richness, and in this case, exclusive species were recorded at the more urbanized sites.
Total taxonomic β diversity was higher than functional and phylogenetic diversity, indicating more significant variability in species composition among different sites. Recently, β diversity has been subdivided into nestedness and turnover components [70], which can provide insights into species turnover and variation in species richness. For taxonomic β diversity, the turnover component made the most significant contribution, a pattern also reported in studies of other vertebrates such as amphibians (e.g., [71]) and birds (e.g., [72]).
Contrary to the previous findings, the nestedness component better explained functional β diversity. This suggests that while sites differ in species richness, they share bat species with similar functional traits, leading to reduced turnover in functional traits. This result contrasts with that obtained by Carvalho et al. [73], who studied bats in a conserved area (Amazon biome) and found that differences in species richness contributed slightly more to total functional β diversity than turnover. They observed a similar pattern in total phylogenetic β diversity of bats, whereas in this study, the turnover component made the greatest contribution to phylogenetic β diversity. This inconsistency in results may be influenced by the fact that few bat species can exploit and adapt to urban areas. As mentioned earlier, urban growth has been documented to reduce phylogenetic diversity in addition to species loss [39,46,60,61,62].
Cluster analyses based on taxonomic, functional, and phylogenetic differences revealed two patterns: (1) closely clustered sites (Central–South) based on the similarity of their taxonomic, functional, and phylogenetic compositions. This similarity pattern correlated with geographic distance has been reported in Oaxaca by Calderón-Patrón et al. [74] and Cornejo-Latorre et al. [75], likely due to similar environmental conditions and levels of urbanization these sites; and (2) distant clustered sites (East–West), possibly because, despite the greater geographic distance between them, the environmental and urbanized conditions at both sites are more similar. Additionally, it is important to consider that some bats (e.g., insectivores) can travel approximately 40 km on average per night in search of food [76].
This information is important because not all components of bat diversity are equally affected by urban development. Our findings provide a framework for focusing on the main effects of urban vegetation, building density, and structure, and the potential impact of acoustic and light pollution on bat species assemblages that could be used in urban planning to enhance the resilience of the urban–nature matrix.
In this study, we followed a multidimensional approach to characterizing bat communities, recognizing that the complexity of ecological systems is best described when focusing on different facets of diversity.
We found bat communities that were phylogenetically related. This type of phylogenetic homogenization could explain the structure and assembly of urban bat communities. According to Darwin’s hypothesis, we can expect that the level of competition in communities with more closely related species would be higher [77]. Darwin speculated that niche overlap among more closely related species would make their coexistence more difficult [78], and there is evidence that interspecific competition with urbanized species prevents less urbanized and closely related species from colonizing cities [79]. However, although evolutionary relationships could be related to interspecific competition and niche differences, they may also be related to differences in average fitness between species [77].
5. Conclusions
Urbanization has altered natural ecosystems, making the study of organisms of interest for biodiversity conservation, such as bats, which provide essential ecosystem services, a priority. Maintaining adequate bat diversity levels within urban and suburban settlements can help enhance the potential resilience of these ecosystems exposed to rapid and continuous vegetation changes. Finally, we consider that our study, the first of its kind in Oaxaca, paves the way for future research and provides valuable information that could be used by local and regional governments and stakeholders worldwide as recommendations for green space planning.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d16090527/s1. Table S1: Functional traits of bats species recorded at the four sites in the city of Oaxaca surrounding municipalities; Figure S1: Phylogeny used to estimate the phylogenetic diversity of bats in southern Mexico.
Author Contributions
M.B.-S. contributed to the design and writing of the document and funding acquisition; G.E.M.-C., fieldwork, writing and editing the document; C.N.M.-R. designed the work, performed the analyses and wrote the document. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by SIP: 20230797 of the National Polytechnic Institute.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We thank E. Galindo and M. Peralta for reviewing this document. We also thank M. C. Lavariega for preparing the study are map. M.B.-S. thanks the Commission for the Operation and Promotion of Academic Activities (COFAA) and the Researcher Performance Incentive Program (EDI) at the National Polytechnic Institute for their support. C.N.M.-R. acknowledges the Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT) for the postdoctoral fellowship (CVU 504386). C.N.M.-R. and M.B.-S. acknowledge the Sistema Nacional de Investigadores (SNI) for its recognition and support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mammal. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Newbold, T.; Scharlemann, J.P.; Butchart, S.H.; Şekercioğlu, Ç.H.; Alkemade, R.; Booth, H.; Purves, D.W. Ecological traits affect the response of tropical forest bird species to land-use intensity. Proc. R. Soc. B Biol. Sci. 2013, 280, 2012–2131. [Google Scholar] [CrossRef] [PubMed]
- Sala, O.E.; Stuart Chapin, F.I.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Téllez-Hernández, E.; Domínguez-Vega, H.; Zuria, I.; Marín-Togo, M.C.; Gómez-Ortiz, Y. Functional and ecological diversity of urban birds: Conservation and redesign of biocultural landscapes. J. Nat. Conserv. 2023, 73, 126395. [Google Scholar] [CrossRef]
- Gutiérrez-Granados, G.; Rodríguez-Zúñiga, M.T. Bats as indicators of ecological resilience in a megacity. Urban Ecosyst. 2024, 27, 479–489. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Law, B.; Banks, P.B. Sensitivity of insectivorous bats to urbanization: Implications for suburban conservation planning. Biol. Conserv. 2012, 146, 41–52. [Google Scholar] [CrossRef]
- Gorresen, P.M.; Willig, M.R.; Strauss, R.E. Multivariate analysis of scale dependent associations between bats and landscape structure. Ecol. Appl. 2005, 15, 2126–2136. [Google Scholar] [CrossRef]
- García-Morales, R.; Moreno, C.E.; Badano, E.I.; Zuria, I.; Galindo-González, J.; Rojas-Martínez, A.E.; Ávila-Gómez, E.S. Deforestation impacts on bat functional diversity in tropical landscapes. PLoS ONE 2016, 11, e0166765. [Google Scholar] [CrossRef]
- García-Luis, M.; Briones-Salas, M. Composición y actividad de la comunidad de murciélagos artropodívoros en parques eólicos del trópico mexicano. Rev. Mex. Biodiv. 2017, 88, 888–898. [Google Scholar] [CrossRef]
- Moreno, C.E.; Calderón-Patrón, J.M.; Arroyo-Rodríguez, V.; Barragán, F.; Escobar, F.; Gómez-Ortiz, Y.; Zuria, I. Measuring biodiversity in the Anthropocene: A simple guide to helpful methods. Biodivers. Conserv. 2017, 26, 2993–2998. [Google Scholar] [CrossRef]
- Moreno, C.E.; Calderón-Patrón, J.M.; Martín-Regalado, N.; Martínez-Falcón, A.P.; Ortega-Martínez, I.J.; Rios-Díaz, C.L.; Rosas, F. Measuring species diversity in the tropics: A review of methodological approaches and framework for future studies. Biotropica 2018, 50, 929–941. [Google Scholar] [CrossRef]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.C.; Moura, N.G. Taxonomic, phylogenetic and functional diversity of an urban Amazonian avifauna. Urban Ecosyst. 2017, 20, 1019–1025. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Su, T.; Zhou, B.; Moravec, D.; Šímová, P.; Liang, W. Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing’s urban parks: Effects of land use and vegetation structure. Urban For. Urban Green. 2017, 23, 84–92. [Google Scholar] [CrossRef]
- Castillo-Figueroa, D.; Pérez-Torres, J. On the development of a trait-based approach for studying Neotropical bats. Pap. Avulsos Zool. 2021, 61, e20216124. [Google Scholar] [CrossRef]
- Cisneros, L.; Fagan, M.E.; Willig, M.R. Effects of human-modified landscapes on taxonomic, functional and phylogenetic dimensions of bats biodiversity. Divers. Distrib. 2015, 21, 523–533. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.L. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef]
- Laity, T.; Laffan, S.W.; González-Orozco, C.E.; Faith, D.P.; Rosauer, D.F.; Byrne, M.; Miller, J.T.; Crayn, D.; Costion, C.; Moritz, C.C.; et al. Phylodiversity to inform conservation policy: An Australian example. Sci. Total Environ. 2015, 534, 131–143. [Google Scholar] [CrossRef]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Davies, T.J.; Ferrier, S.; Fritz, S.A.; Mazel, F. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 2017, 92, 698–715. [Google Scholar] [CrossRef]
- Ortiz, P.M.A.; Hernández, J.R.S.; Figueroa Mah-Eng, J.M. Reconocimiento fisiográfico y geomorfológico. In Biodiversidad de Oaxaca; García-Mendoza, A.J., Ordónez, M.J., Briones-Salas, M., Eds.; Instituto de Biología, Universidad Nacional Autónoma de México-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Found: Oaxaca, Mexico, 2004; pp. 43–45. [Google Scholar]
- Torres, C.T. Tipos de vegetación. In Biodiversidad de Oaxaca; García-Mendoza, A.J., Ordónez, M.J., Briones-Salas, M., Eds.; Instituto de Biología, Universidad Nacional Autónoma de México-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Found: Oaxaca, Mexico, 2004; pp. 105–117. [Google Scholar]
- Trejo, I. Clima. In Biodiversidad de Oaxaca; García-Mendoza, A.J., Ordónez, M.J., Briones-Salas, M., Eds.; Instituto de Biología, Universidad Nacional Autónoma de México-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Found: Oaxaca, Mexico, 2004; pp. 67–85. [Google Scholar]
- INEGI (Instituto Nacional de Estadística y Geografía). Censo Nacional de Vivienda; Instituto Nacional de Estadística y Geografía: San Luis Potosí, México, 2010. [Google Scholar]
- INEGI (Instituto Nacional de Estadística y Geografía). Uso de Suelo y Vegetación serie V, Escala 1:250,000; Instituto Nacional de Estadística y Geografía: San Luis Potosí, México, 2013. [Google Scholar]
- Vargas-Contreras, J.A.; Escalona-Segura, G.; Cú-Vizcarra, J.D.; Arroyo-Cabrales, J.; Medellín, R.A. Estructura y Diversidad de los Ensambles de Murciélagos en el Centro y sur de Campeche, México; Avances en el estudio de los mamíferos de México: Ciudad de México, México, 2008; pp. 551–577. [Google Scholar]
- Medellín, R.A.; Arita, H.T.; Sánchez, Ó. Identificación de los Murciélagos de México: Clave de Campo; Instituto de Ecología, UNAM: Ciudad de México, México, 2008; pp. 28–63. [Google Scholar]
- Ramírez-Pulido, J.; González-Ruiz, N.; Gardner, A.L.; Arroyo-Cabrales, J. List of Recent Land Mammals of Mexico; Museum of Texas Tech University: Lubbock, TX, USA, 2014; pp. 6–16. [Google Scholar]
- Ortega, J.; MacSwiney, M.C.; Zamora, V. Compendio de los Llamados de Ecolocalización de los Murciélagos Insectívoros Mexicanos; Asociación Mexicana de Mastozoología, A.C. y CONABIO: Ciudad de México, México, 2022; pp. 24–217. [Google Scholar]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.F.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Safi, K.; Cianciaruso, M.V.; Loyola, R.D.; Brito, D.; Armour-Marshall, K.; Diniz-Filho, J.A.F. Understanding global patterns of mammalian functional and phylogenetic diversity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2536–2544. [Google Scholar] [CrossRef]
- Martín-Regalado, C.N.; Briones-Salas, M. Rasgos funcionales para mamíferos terrestres del sur de México. Caldasia 2024, 46, 1–29. [Google Scholar]
- Petchey, O.L.; Gaston, K.J. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Gotelli, N.J. Null model analysis of species co-occurrence patterns. Ecology 2000, 81, 2606–2621. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M.E. Measuring functional diversity from multiple traits, and other tools for functional ecology. In R Package FD; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Oksanen, J. Vegan: Community Ecology Package. 2010. Available online: http://vegan.r-forge.r-project.org/ (accessed on 10 June 2024).
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Faith, D.P. The PD phylogenetic diversity framework: Linking evolutionary history to feature diversity for biodiversity conservation. In Biodiversity Conservation and Phylogenetic Systematics; Springer: Berlin/Heidelberg, Germany, 2016; pp. 39–56. [Google Scholar]
- Upham, N.S.; Esselstyn, J.A.; Jetz, W. Inferring the mammal tree: Species- level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS Biol. 2019, 17, e3000494. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Baselga, A.D.; Orme, S.; Villeger, J.; De Bortoli, F. Leprieur. Betapart: Partitioning Beta Diversity into Turnover and Nestedness Components. R Package Version 1.5.1. Available online: https://CRAN.R-project.org/package=betapart. (accessed on 19 April 2023).
- Ricotta, C.; Burrascano, S. Beta diversity for functional ecology. Preslia 2008, 80, 61–71. [Google Scholar]
- Graham, C.H.; Fine, P.V.A. Phylogenetic beta diversity: Linking ecological and evolutionary processes across space in time. Ecol. Lett. 2008, 11, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Espinal-Palomino, R.; Ibarra-Cerdeña, C.N. Siluetas en el cielo: Murciélagos en mi ciudad. Therya Ixmana 2022, 1, 42–44. [Google Scholar] [CrossRef]
- Pierson, E.D. Tall trees, deep holes, and scarred landscapes: Conservation biology of North American bats. In Bat Biology and Conservation; Kunz, T.H., Racey, P.A., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 309–325. [Google Scholar]
- Palheta, L.R.; Urbieta, G.L.; Brasil, L.S.; Dias-Silva, K.; Da Silva, J.B.; Graciolli, G.; Aguilar, L.M.S.; Vieira, T.B. The effect of urbanization on bats and communities of bat flies (Diptera: Nycteribiidae and Streblidae) in the Amazon, northern Brazil. Acta Chiropt. 2020, 22, 403–416. [Google Scholar] [CrossRef]
- Briones-Salas, M.; Cortés-Marcial, M.; Lavariega, M.C. Diversidad y distribución geográfica de los mamíferos terrestres del estado de Oaxaca, México. Rev. Mex. Biodiv. 2015, 8, 685–710. [Google Scholar] [CrossRef]
- Gurrusquieta-Navarro, M.C. Dieta de Murciélagos Frugívoros en la Zona Urbana de Cuernavaca Morelos. Bachelor Thesis, Universidad Autónoma de Morelos, Morelos, Mexico, 2015. [Google Scholar]
- Ferreyra-García, D.; Saldaña-Vázquez, R.A.; Mendoza, E. Macrotus waterhousii en Morelia, México. Boletín Red Latinoam. Para Conserv. Murciélagos 2017, 8, 20–23. [Google Scholar]
- Jung, K.; Kalko, E.K. Adaptability and vulnerability of high flying Neotropical aerial insectivorous bats to urbanization. Divers. Distrib. 2011, 17, 262–274. [Google Scholar] [CrossRef]
- Starik, N.; Gygax, L.; Göttert, T. Unexpected bat community changes along an urban-rural gradient in the Berlin-Brandenburg metropolitan area. Sci. Rep. 2024, 14, 10552. [Google Scholar] [CrossRef]
- Saldaña-Vázquez, R.A.; Ferreyra-García, D.; Vázquez-Domínguez, G. ¿Murciélagos en mi ciudad? Consejos y reflexiones para convivir con ellos. Digit. Cienc.@ UAQRO 2019, 12, 28–32. [Google Scholar]
- Rodríguez-Aguilar, G.; Orozco-Lugo, C.L.; Vleut, I.; Vazquez, L.B. Influence of urbanization on the occurrence and activity of aerial insectivorous bats. Urban Ecosyst. 2017, 20, 477–488. [Google Scholar] [CrossRef]
- Fenton, M.B.; Bogdanowicz, W. Relationships between external morphology and foraging behaviour: Bats in the genus Myotis. Can. J. Zool. 2002, 80, 1004–1013. [Google Scholar] [CrossRef]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Tsianou, M.A.; Kallimanis, A.S. Different species traits produce diverse spatial functional diversity patterns of amphibians. Biodivers. Conserv. 2016, 25, 117–132. [Google Scholar] [CrossRef]
- Gordon, R.; Ivens, S.; Ammerman, L.; Fenton, M.; Littlefair, J.; Ratcliffe, J.; Clare, E. Molecular diet analysis finds an insectivorous desert bat community dominated by resource sharing despite diverse echolocation and foraging strategies. Ecol. Evol. 2019, 9, 3117–3129. [Google Scholar] [CrossRef]
- Nee, S.; May, R.M. Extinction and the Loss of Evolutionary History. Science 1997, 278, 692–694. [Google Scholar] [CrossRef]
- Purvis, A.; Agapow, P.M.; Gittleman, J.L.; Mace, G.M. Nonrandom extinction and the loss of evolutionary history. Science 2000, 288, 328–330. [Google Scholar] [CrossRef]
- Huang, S.; Davies, T.J.; Gittleman, J.L. How global extinctions impact regional biodiversity in mammals. Biol. Lett. 2011, 8, 222–225. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- García-Luis, M.; Briones-Salas, M.; Lavariega, M.C. Bat species richness in the region of the Central Valleys of Oaxaca, México. Arxius Misc. Zool. 2019, 17, 1–11. [Google Scholar] [CrossRef]
- Vissoto, M.; Schneiberg, I.; Varassin, I.G.; de Araujo, A.C.; Maruyama, P.K.; Vizentin-Bugoni, J. Frugivory and seed dispersal in tropical urban areas: A review. In Ecology of Tropical Cities: Natural and Social Sciencies; Angeoletto, F., Tryjanowski, P., Fellowes, M., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2023; pp. 1–33. [Google Scholar]
- Poma, M.G.G.; Poma, F.C.G. Métodos de exclusión de murciélagos hematófagos en edificaciones humanas. Boletín Malariol. Salud Ambient. 2022, 62, 1363–1370. [Google Scholar] [CrossRef]
- Cerón-Hernández, J.A.; Serna-Lagunes, R.; Torres-Cantú, G.B.; Llarena-Hernández, C.R.; Mora-Collado, N.; García-Martínez, M.Á. Diversidad, tipos de dieta de murciélagos y su respuesta a bordes de bosque mesófilo de montaña, Veracruz, México. Ecosis. Recur. Agropec. 2022, 9, 1–13. [Google Scholar] [CrossRef]
- Tuttle, M.D.; Moreno, A. Murciélagos Cavernícolas del Norte de México. Su Importancia y Problemas de Conservación; Bat Conservation International Inc.: Austin, TX, USA, 2005. [Google Scholar]
- Calderón-Patrón, J.; Briones-Salas, M.; Moreno, C.E. Diversidad de murciélagos en cuatro tipos de bosque de la Sierra Norte de Oaxaca, México. Therya 2013, 4, 121–137. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Calderón-Patrón, J.M.; Peña-Joya, K.E.; Téllez-López, J.; Canales-Gómez, E.P. Dissimilarity among Species and Higher Taxa of Amphibians in a Hotspot of Biodiversity and Endemism in the Neotropics. Diversity 2024, 16, 224. [Google Scholar] [CrossRef]
- Zeng, C.; Li, W.; Ding, P.; Peng, Y.; Wu, L.; Si, X. A landscape level analysis of bird taxonomic, functional and phylogenetic β-diversity in habitat island systems. J. Biogeogr. 2022, 49, 1162–1175. [Google Scholar] [CrossRef]
- Carvalho, W.D.; Fluck, I.E.; de Castro, I.J.; Hilário, R.R.; Martins, A.C.M.; de Toledo, J.J.; Bobrowiec, P.E. Elevation drives taxonomic, functional and phylogenetic β-diversity of phyllostomid bats in the Amazon biome. J. Biogeogr. 2023, 50, 70–85. [Google Scholar] [CrossRef]
- Calderón-Patrón, J.M.; Moreno, C.E.; Pineda-López, R.; Sánchez-Rojas, G.; Zuria, I. Vertebrate dissimilarity due to turnover and richness differences in a highly beta-diverse region: The role of spatial grain size, dispersal ability and distance. PLoS ONE 2013, 8, e82905. [Google Scholar] [CrossRef]
- Cornejo-Latorre, C.; Moreno, C.E.; Martín-Regalado, C.N.; Briones-Salas, M. Taxonomic and phylogenetic beta diversity of cricetid rodents in Oaxaca, southern Mexico. J. Mammal. 2020, 101, 1451–1462. [Google Scholar] [CrossRef]
- Gutiérrez-Granados, G. Murciélagos viajeros: El misterio de las migraciones nocturnas. Therya Ixmana 2023, 2, 62–63. [Google Scholar] [CrossRef]
- Godoy, O.; Kraft, N.J.; Levine, J.M. Phylogenetic relatedness and the determinants of competitive outcomes. Ecol. Lett. 2014, 17, 836–844. [Google Scholar] [CrossRef]
- Darwin, C. The Origin of Species, 1859–1959. Bios 1959, 30, 67–72. [Google Scholar]
- Møller, A.P.; Díaz, M. Avian preference for close proximity to human habitation and its ecological consequences. Curr. Zool. 2018, 64, 623–630. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).