
Fishing for Estuarine Oomycetes

AgroBioInstitute, Agricultural Academy, 8 Dragan Tsankov Blvd., 1164 Sofia, Bulgaria
Diversity 2024, 16(9), 530; https://doi.org/10.3390/d16090530
Submission received: 10 July 2024
/
Revised: 23 August 2024
/
Accepted: 28 August 2024
/
Published: 1 September 2024
(This article belongs to the Special Issue Microbial Diversity and Culture Collections Hotspots in 2024)
Abstract
:Oomycetes are water molds that are frequently isolated during a survey of waterways. Biodiversity of oomycetes in the estuary region of the Veleka River in Bulgaria was investigated in 2021. A total number of 32 isolates were derived using the baiting method. Species identification based on sequencing of the ITS region divided them into four different genera: Phytophthora (P.), Phytopythium (Pp.), Pythium (Py.) and Elongisporangium (E.). The most abundant species in the studied region was P. lacustris (sixteen isolates), followed by P. honggalleglyana (nine isolates). P. bilorbang and P. inundata were represented by only one isolate each and were recognized for the first time in Bulgaria. The genus Phytopythium was presented by two isolates that belong to different species, Pp. litorale and Pp. citrinum. In the obtained collection, the genera Pythium and Elongisporangium were represented by only one species each, Py. angustatum (one isolate) and E. anandrum (two isolates), respectively. Colony morphology of the eight collected oomycete species was characterized by cultivation of selected isolates on three different media. Potential host species of the isolated estuarine oomycetes were estimated by pathogenicity tests conducted with sixteen plants from ten diverse families. P. lacustris and P. honggalleglyana demonstrated a higher aggressiveness among Phytophthora isolates, whereas P. bilorbang and P. inundata showed less ability to infect the tested plant species. Similar pathogenicity and a potential host range for both Phytopythium species were observed. Less aggressive against analyzed plants in this study were Py. angustatum and E. anandrum.
Keywords:
aquatic microorganisms; Phytophthora; Pythium; Phytopythium; Elongisporangium; baiting; ITS; diversity; pathogenicity1. Introduction
A number of oomycetes from the genera Phytophthora, Pythium, Phytopythium and Elongisporangium are important pathogens on agricultural crops and native plants, affecting both crop growing and natural ecosystems [1,2,3]. On the other hand, some oomycete species appear to be saprophytes in aquatic environments [4]. Oomycetes are water molds and are frequently isolated during a survey of waterways. Many of them are associated with wet soil conditions, and their spread is often favored by seasonal floods [5,6,7,8]. Numerous novel species and hybrids derived from riparian ecosystems in various areas of the world have been reported in recent years [8,9,10,11]. Among the most abundantly distributed in water habitats have been found to be Phytophthora species from clades 6 and 9 [12,13,14]. Some members of these groups are strongly associated with host plants from forests and riparian ecosystems [7,13,14,15,16]. However, knowledge on the diversity of oomycetes in estuarine and coastal environments is fragmentary. Out of about 2000 species of oomycetes reported, the highest diversity has been observed in terrestrial plant pathogens, and while some species have been isolated from water sources, only about 60 have been derived from marine environments [17,18]. Some of these species are associated with diseases on seaside and vegetated coastal ecosystems [19,20,21].
The Veleka is a river in the southeastern part of Bulgaria that flows into the Black Sea. The mouth of the river is a unique area with specific features because of its geographic characteristics. The estuary forms a partially enclosed coastal zone (sandbar) causing the formation of a transition area between a river and a marine environment. The fresh water mixes with the sea water at the lower and middle estuary of the river. Occasionally, the mouth is blocked by the sand, which is especially pronounced during rough seas, and leads to flooding of the surrounding area. The region is characterized by transitional Mediterranean climatic conditions with a strong influence by the sea. The vegetation in the area is diverse, as there are forest and woody species like alder, bamboo, hornbeam, elm, ash and others along the banks. Dense emergent and sparse vegetation, as well as grassy meadow species, are widespread in the water and coastal wetland.
The isolation of oomycetes from diseased plants assists in their identification, as some pathogen species are strongly associated with specific host plants. However, when the species are derived from water sources, their pathogenicity and host range are unclear. A large amount of data is collected for some oomycetes, but for the species that are rarely isolated or have been identified recently, further investigations are needed. Frequently a limited number of susceptible plant species are known, and the importance of these oomycetes for the wide variety of potential hosts requires additional studies.
An investigation of oomycete diversity in the estuary region of the Veleka River is presented in this study. A total number of 32 isolates belonging to four different genera, Phytophthora, Phytopythium, Pythium and Elongisporangium, were collected and identified. Their ability to infect 16 different plants from 10 diverse families was evaluated. The most common oomycetes in the studied area and the most aggressive species among them were noted.
2. Materials and Methods
2.1. Sampling Area
The study covers the estuary region of the Veleka River (Figure 1).
Different sites for investigation of oomycete diversity were selected:
- Next to a sandbar between the river and the sea—samples with numbers RVel2021/115 and RVel2021/120 (GPS coordinates: 42.066845, 27.971472). The river bottom in the region is sandy and the main vegetation is common reed (Phragmites australis).
- In the liman area of the river—samples with numbers RVel2021/114 and RVel2021/118 (GPS coordinates: 42.066310, 27.971529) and RVel2021/119 (GPS coordinates: 42.064896, 27.970419). In addition to the reed, there are grassy meadow species and dense riparian vegetation.
- In the forest area of the river—samples with numbers RVel2021/112 (GPS coordinates: 42.060125, 27.966534), RVel2021/113 and RVel2021/116 (GPS coordinates: 42.061555, 27.967234) and RVel2021/117 (GPS coordinates: 42.063026, 27.968246). There is forestry vegetation that includes alder, hornbeam, elm, ash, lianas and creeping plants.
At the time of collecting samples 119 and 120, the mouth of the river was blocked by the sand, and the water did not flow into the sea and spread over a large area of the surrounding region. This allowed zoospores of species that probably are not typical of the river, but rather of the wetlands around it, to be attracted by the baits and isolated.
2.2. Baiting and Isolation of Oomycetes
Baiting of oomycetes was performed in May, July and September of 2021. Zoospores of oomycetes were attracted to detached leaves of Rhododendron, oak and oleander that were placed into meshy baits in the river for a period of 3 days. Only young and healthy leaves were selected for the experiment. After collection of the baits, they were saved in a cooler and transferred to the laboratory.
The leaves from the baits were surface-sterilized, and pieces of developed necrosis with a part of healthy zone were cut out. They were cultivated on selective PARNHB media (carrot agar supplemented with 10 mg Pimaricin, 250 mg Ampicillin, 10 mg Rifampicin, 50 mg Nystatin, 50 mg Hymexazol and 15 mg Benomyl/L) at 20 °C for 5 days. A mycelium plug from each colony with different morphological type was transferred for cultivation on WA (water agar) for a period of 3 days. A hyphal tip from purified colonies was transferred onto a fresh CA medium (carrot agar; 16 g agar, 3 g CaCO3, 100 mL carrot juice/L), and cultivation at 20 °C was performed.
2.3. Species Identification
DNA was isolated from mycelia of 10-day-old cultures using DNeasy Plant Mini Kit (QIAGEN GmbH, Venlo, The Netherlands). PCR amplification of the ITS region was performed with primers ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′). The following PCR program was applied: 96 °C for 2 min, followed by 35 cycles of 96 °C for 1 min, 55 °C for 1 min, 72 °C for 2 min and final elongation at 72 °C for 10 min (Ristaino et al. 1998). The PCR was performed by PuReTaqTM Ready-To-GoTM PCR beads (GE Healthcare Life Sciences, Chicago, IL, USA), according to the manufacturer’s instructions. The resulting PCR products were sequenced at GATC Biotech AG (Constance, Germany). Species identification of all isolates included in the study was performed by searching for the highest-homology ITS sequences within the NCBI database using BLAST.
2.4. Colony Morphology
One isolate from each species collected from the Veleka River was selected for characterization of the colony morphology, as follows: P. lacustris (RVel2021/113d), P. bilorbang (RVel2021/119a), P. inundata (RVel2021/119b), P. honggalleglyana (RVel2021/120c), Pp. citrinum (RVel2021/113a), Pp. litorale (RVel2021/113b), Py. angustatum (RVel2021/114c) and E. anandrum (RVel2021/115d). They were cultivated on three different media: CA, V8A (vegetable agar: 16 g agar, 3 g CaCO3, 100 mL Campbell’s V8 juice/L) and PDA (potato dextrose agar, Difco®) at 20 °C for a period of 10 days for Phytophthora isolates and 5 days for Phytopythium, Pythium and Elongisporangium isolates.
2.5. Testing for Potential Host Plants
Since all collected isolates were derived from the river, not from diseased plants, testing for potential hosts was performed. The isolates that were selected for characterization of the colony morphology were also used for these experiments. Plant species that are dominant in the ecosystem of the Veleka River or species that are among most common in the country were chosen for pathogenicity tests. A total number of sixteen different species from ten diverse families (Fabaceae, Fagaceae, Salicaceae, Rosaceae, Araliaceae, Vitaceae, Ranunculaceae, Asteraceae, Plantaginaceae and Poaceae) were selected. These are representatives of trees, including the Judas tree (Cercis siliquastrum), common oak (Quercus robur), European aspen (Populus tremula) and willow (Salix babylonica); bushes, such as dog rose (Rosa canina) and blackberry (Rubus fruticosus); as well as some perennial and herbaceous plants like common ivy (Hedera helix), five-leaved ivy (Parthenocissus quinquefolia), old man’s beard (Clematis vitalba), alfalfa (Medicago sativa), clover (Trifolium repens), common daisy (Bellis perennis), common yarrow (Achillea millefolium), ribwort plantain (Plantago lanceolata), broadleaf plantain (Plantago major) and green foxtail (Setaria viridis). The pathogenicity tests were performed with leaves and cuttings from the selected potential hosts. They were conducted twice with all tested isolated and plant species.
Detached leaves from healthy plants were selected, and the baiting technique was applied. The leaves were placed into plastic trays containing a mix of sterile distilled water and non-sterile spring water (1:1). Mycelial plugs (10 × 10 mm) of 7-day-old culture from each tested isolate were added into test variants, whereas control trays were prepared with the same water mixture, but without mycelial sample. All leaves were incubated at 22/20 °C (16 h day/8 h night) for a period of 5 to 14 days depending on the development of disease symptoms. Daily monitoring for appearance and expansion of necrotic lesions on tested leaves was performed.
Along with the pathogenicity tests with leaves, young cuttings (6–7 cm) of the selected plant species were arranged on wet paper in plastic trays. Mycelial plugs (3 × 3 mm) of 7-day-old culture from each tested oomycete isolate were placed on the middle part of the cuttings after slight surface injury. Plastic trays were covered with transparent film to maintain a high humidity and were incubated at 22/20 °C (16 h day/8 h night) for 2 weeks. A daily monitoring of the inoculated cuttings for appearance of disease symptoms was performed.
The severity of disease symptoms was evaluated according to the following scale: no symptoms—healthy leaf/cutting (-); small dot necroses—low susceptibility (+); necrotic area about 50% of the leaf/cutting—middle level of sensitivity (++); necrosis covers 90–100% of the leaf/cutting—totally affected (+++).
3. Results
3.1. Diversity of Oomycetes
A total number of 32 isolates were collected from the estuary region of the Veleka River (Table 1). The species identification of the isolates was determined by sequencing of the ITS region and is based on 99–100% homology with corresponding species in the NCBI database. The isolated oomycetes belong to four different genera: Phytophthora, Phytopythium, Pythium and Elongisporangium, represented by 27, two, one and two isolates, respectively.
The genus Phytophthora was represented by four species: P. lacustris, P. honggalleglyana, P. bilorbang and P. inundata. The most abundant species of oomycetes in the study was P. lacustris (sixteen isolates), followed by P. honggalleglyana (nine isolates), corresponding to 50% and 28% of the collected isolates, respectively. The two rarest species from the genus Phytophthora, P. bilorbang and P. inundata, which are represented by only one isolate each (3% of the isolates), were recognized for the first time in Bulgaria. The genus Phytopythium was represented by only two isolates that belong to different species: Pp. litorale and Pp. citrinum (Table 1). The first of them has been found in another river in the northern part of the country previously [22], whereas the second species has not been reported in Bulgaria up to now. The genera Pythium and Elongisporangium were represented by one species each, Py. angustatum (one isolate) and E. anandrum (two isolates), respectively (Table 1).

{kind=link}
{kind=link}
{kind=link}
Table 1.
List of Phytophthora, Phytopythium, Pythium and Elongisporangium isolates included in the study.
Table 1.
List of Phytophthora, Phytopythium, Pythium and Elongisporangium isolates included in the study.
| No. of Isolate | Species | GenBank No. | Type of Bait | Location | Baiting Period |
|---|---|---|---|---|---|
| RVel2021/112a | P. lacustris | PQ107011 | rhododendron | 42.060125, 27.966534 | 2 May 2021 (start) 5 May 2021 (end) |
| RVel2021/112c | P. lacustris | PQ107013 | rhododendron | ||
| RVel2021/112d | P. lacustris | PQ107014 | oak | ||
| RVel2021/112f | P. lacustris | PQ107016 | oleander | ||
| RVel2021/113a | Pp. citrinum | PQ110268 | oak | 42.061555, 27.967234 | |
| RVel2021/113b | Pp. litorale | PQ107594 | oak | ||
| RVel2021/113c | E. anandrum | PQ107604 | oak | ||
| RVel2021/113d | P. lacustris | PQ107017 | oleander | ||
| RVel2021/113e | P. lacustris | PQ107019 | oleander | ||
| RVel2021/114a | P. lacustris | PQ107021 | rhododendron | 42.066310, 27.971529 | |
| RVel2021/114b1 | P. lacustris | PQ107022 | rhododendron | ||
| RVel2021/114b2 | P. lacustris | PQ107024 | rhododendron | ||
| RVel2021/114c | Py. angustatum | PQ107654 | oak | ||
| RVel2021/114d | P. lacustris | PQ107026 | oak | ||
| RVel2021/114e | P. lacustris | PQ107025 | oleander | ||
| RVel2021/114f | P. lacustris | PQ107479 | oleander | ||
| RVel2021/115a | P. lacustris | PQ107487 | rhododendron | 42.066845, 27.971472 | |
| RVel2021/115b | P. lacustris | PQ107488 | rhododendron | ||
| RVel2021/115c | P. lacustris | PQ107490 | oak | ||
| RVel2021/115d | E. anandrum | PQ107632 | oak | ||
| RVel2021/115e | P. lacustris | PQ107492 | oleander | ||
| RVel2021/116a | P. honggalleglyana | PQ107495 | rhododendron | 42.061555, 27.967234 | 16 July 2021 (start) 18 July 2021 (end) |
| RVel2021/116b | P. honggalleglyana | PQ107496 | rhododendron | ||
| RVel2021/117a | P. honggalleglyana | PQ107498 | rhododendron | 42.063026, 27.968246 | |
| RVel2021/117b | P. honggalleglyana | PQ107497 | rhododendron | ||
| RVel2021/118a | P. honggalleglyana | PQ107499 | rhododendron | 42.066310, 27.971529 | |
| RVel2021/118b | P. honggalleglyana | PQ107501 | rhododendron | ||
| RVel2021/119a | P. bilorbang | PQ107517 | rhododendron | 42.064896, 27.970419 | 9 September 2021 (start) 11 September 2021 (end) |
| RVel2021/119b | P. inundata | PQ107518 | rhododendron | ||
| RVel2021/120a | P. honggalleglyana | PQ107500 | rhododendron | 42.066845, 27.971472 | |
| RVel2021/120b | P. honggalleglyana | PQ107504 | rhododendron | ||
| RVel2021/120c | P. honggalleglyana | PQ107514 | rhododendron |
Incubation of the selected isolates from the eight oomycete species on three different media (CA, V8A and PDA) demonstrated diversity in the pattern of colony morphology, growth rate and preferences to nutritional compounds of some species. All Phytophthora isolates showed relatively fast growth on CA and V8A, whereas significant radial development on PDA was observed only for P. inundata, isolate RVel2021/119b (Figure 2). A slight increase in growth of P. lacustris (RVel2021/113d) and P. honggalleglyana (RVel2021/120c) colonies on potato dextrose agar was recorded, but P. bilorbang, isolate RVel2021/119a, was strongly restricted on this medium. Mycelial growth of P. lacustris, isolate RVel2021/113d, was characterized by chrysanthemum pattern and limited aerial mycelium on media CA and V8A (Figure 2A1–A3). P. bilorbang, isolate RVel2021/119a, formed submerged colonies, petaloid on CA and with a stellate morphology pattern on V8A (Figure 2B1–B3). P. inundata, isolate RVel2021/119b, showed petaloid colony morphology on CA and V8A, and a chrysanthemum pattern on PDA with aerial mycelium on all three media (Figure 2C1–C3). P. honggalleglyana, isolate RVel2021/120c, demonstrated a petaloid pattern of colony with aerial growth on all tested media, especially pronounced on CA (Figure 2D1–D3).
Figure 2.
Colony morphology of Phytophthora isolates on CA (first row), V8A (second row) and PDA (third row) after 10 days cultivation at 20 °C. (A1–A3) P. lacustris, isolate RVel2021/113d, (B1–B3) P. bilorbang, isolate RVel2021/119a, (C1–C3) P. inundata, isolate RVel2021/119b, and (D1–D3) P. honggalleglyana, isolate RVel2021/120c.
Figure 2.
Colony morphology of Phytophthora isolates on CA (first row), V8A (second row) and PDA (third row) after 10 days cultivation at 20 °C. (A1–A3) P. lacustris, isolate RVel2021/113d, (B1–B3) P. bilorbang, isolate RVel2021/119a, (C1–C3) P. inundata, isolate RVel2021/119b, and (D1–D3) P. honggalleglyana, isolate RVel2021/120c.
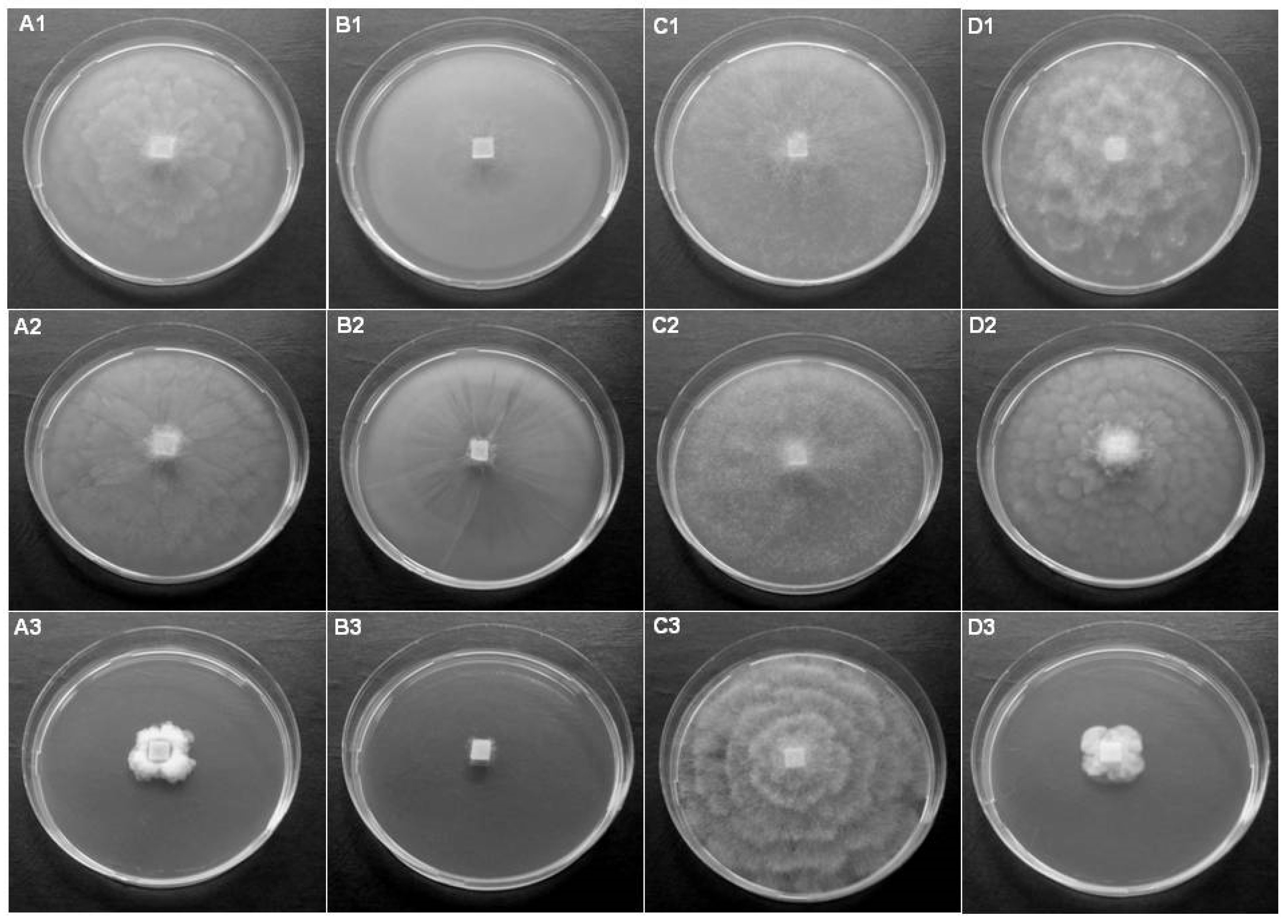
Figure 3.
Colony morphology of Phytopythium, Pythium and Elongisporangium isolates on CA (first row), V8A (second row) and PDA (third row) after 5 days cultivation at 20 °C. (A1–A3) Pp. citrinum, isolate RVel2021/113a, (B1–B3) Pp. litorale, isolate RVel2021/113b, (C1–C3) Py. angustatum, isolate RVel2021/114c, and (D1–D3) E. anandrum, isolate RVel2021/115d.
Figure 3.
Colony morphology of Phytopythium, Pythium and Elongisporangium isolates on CA (first row), V8A (second row) and PDA (third row) after 5 days cultivation at 20 °C. (A1–A3) Pp. citrinum, isolate RVel2021/113a, (B1–B3) Pp. litorale, isolate RVel2021/113b, (C1–C3) Py. angustatum, isolate RVel2021/114c, and (D1–D3) E. anandrum, isolate RVel2021/115d.

An extremely rapid mycelial growth on all tested media of the studied isolates from the genera Phytopythium, Pythium and Elongisporangium was observed (Figure 3). Typical for all isolates was formation of aerial mycelium on all three media used. Similar morphology for both Phytopythium species was monitored. Pp. citrinum, isolate RVel2021/113a, and Pp. litorale, isolate RVel2021/113b, formed petaloid to rosaceous colonies on CA and V8A, and a rosette pattern of mycelial growth on PDA (Figure 3A1–A3,B1–B3). Py. angustatum, isolate RVel2021/114c, also showed petaloid to rosaceous colonies (Figure 3C1–C3). E. anandrum, isolate RVel2021/115d, demonstrated stellate mycelial growth on CA and V8A, and a rosette pattern on PDA with profuse aerial mycelium (Figure 3D1–D3).
3.2. Investigation of Potential Host Plants
In order to determine potential host species of isolates, a number of pathogenicity tests with detached leaves and cuttings from various plants were conducted (Table 2).
P. lacustris (RVel2021/113d) and P. honggalleglyana (RVel2021/120c) demonstrated higher aggressiveness among Phytophthora isolates, especially to the tree species, such as C. siliquastrum, Q. robur and P. tremula, as well as to some other plants such as R. canina and P. quinquefolia. Significant sensitivity of herbaceous plants M. sativa and T. repens to P. honggalleglyana was also observed. Moderate pathogenicity by P. lacustris against S. babylonica, C. vitalba, B. perennis, A. millefolium and S. viridis was detected. A similar effect to H. helix, P. quinquefolia, C. vitalba and B. perennis after inoculation with P. honggalleglyana was also observed. In contrast to both dominant Phytophthora species, P. bilorbang (RVel2021/119a) and P. inundata (RVel2021/119b) showed less ability to infect the tested plant species. More sensitive to P. bilorbang were various species including P. tremula, P. lanceolata and B. perennis, whereas P. inundata affected mainly the herbaceous plants P. quinquefolia, T. repens, P. lanceolata and A. millefolium. A total inability of the studied Phytophthora species to infect R. fruticosus was established. Relatively resistant to infection were also H. helix and P. major.
Both Phytopythium isolates (RVel2021/113a and RVel2021/113b) demonstrated comparable pathogenicity and potential host species. Similar to Phytophthora isolates, higher aggressiveness against C. siliquastrum, Q. robur, P. tremula, P. quinquefolia and R. canina was monitored. Moderate ability to infect C. vitalba, M. sativa and A. millefolium by Pp. citrinum (RVel2021/113a) and Pp. litorale (RVel2021/113b) was observed. Herbaceous plants R. fruticosus, H. helix and P. major were resistant to the tested Phytopythium species. These results showed similar potential host plants for the pathogens from the genus Phytophthora and the genus Phytopythium among tested tree species, bushes and herbaceous plants.
Py. angustatum (RVel2021/114c) showed increased aggressiveness against Q. robur and the herbaceous plants P. lanceolata, B. perennis and A. millefolium. The species failed to infect most of the tested plants (nine out of 16). Similarly, eight plant species were not affected by E. anandrum (RVel2021/115d). The studied isolates demonstrated pathogenicity against Q. robur, P. tremula, H. helix and T. repens.
4. Discussion
A number of investigations on the diversity of the genus Phytophthora in Bulgaria were published in the last few years and some species, such as P. pseudosyringae, P. gallica and P. polonica, have been reported for the first time in the country [23,24,25], as well as a new hybrid species P. × sansomeana [11]. In contrast, information on the isolation and identification of the representatives from the genera Phytopythium, Pythium and Elongisporangium is limited [22,26,27,28]. Fishing for estuarine oomycetes in the firth region of the Veleka River resulted in accumulation of new data for species diversity in the country. Eight different species including P. lacustris, P. bilorbang, P. inundata, P. honggalleglyana, Pp. litorale, Pp. citrinum, Py. angustatum and E. anandrum were detected based on sequencing of the ITS region. Additional information for the collected oomycetes by characterization of colony morphology confirmed the species identification according to compliance with the main features described in the corresponding databases (IDphy, 1PPYTG, 1PYTHG).
In the present study, P. lacustris and P. honggalleglyana were determined as the most distributed species from the genus Phytophthora in the firth of the Veleka River. In contrast, two other members of the genus, P. bilorbang and P. inundata, were represented by only one isolate each and were identified for the first time in Bulgaria. The results also showed that clade 6 (P. lacustris, P. bilorbang and P. inundata) dominates in terms of species diversity and in number of Phytophthora isolates obtained (18 out of 27; 67%), whereas clade 9 is represented by nine isolates (33%) that belong to only one species (P. honggalleglyana). No species from the remaining nine Phytophthora clades were isolated at the time of this survey. These results confirm the statement that the Phytophthora species from clades 6 and 9 are among the most abundantly distributed in water ecosystems [12,13,14].
The species P. lacustris is widespread in Europe and has been reported as a weak to moderate pathogen to Alnus, Prunus and Salix [29]. According to a large-scale investigation of the potential hosts, the species was not able to infect a number of crop plants such as pumpkin (Cucurbita spp.), onion (Allium cepa), chili pepper (Capsicum annuum), alfalfa (M. sativa), corn (Zea mays), oats (Avena sativa) and barley (Hordeum vulgare) [15]. The results of our tests confirmed the moderate aggressiveness of P. lacustris to S. babylonica, as well as its pathogenicity to other forest species and a number of perennial and herbaceous plants. In addition, the resistance of M. sativa to the species reported previously [15] was also proved in this study.
The oomycete P. honggalleglyana is associated mainly with water ecosystems such as rivers, streams and irrigation sources, and has been reported in several European countries, including Austria, Spain and Italy [7,16,30,31]. The species has been isolated for the first time as a pathogen on ornamental plants [32]. Lately, P. honggalleglyana has been associated with alder decline across Europe, including Spain [16], Italy [31] and more recently in Bulgaria [25]. The current study presented a variety of potential host plants, including trees and perennial and herbaceous species. It is notable that all P. honggalleglyana isolates were obtained in the summer season, when the water temperature is the highest, while in the spring, this species was not detected in any of the collected samples. This is most likely due to the fact that species from clade 9, to which P. honggalleglyana belongs, tolerate high temperatures and can grow even at 35–40 °C, in addition to their ability to develop quickly [32].
The species P. bilorbang has been recently designated as an exotic pathogen for European countries [33]. It has been isolated in Italy from the rhizosphere soil of Platanus orientalis by baiting [33] and has been also determined as a pathogen on trees from the Oleaceae family, including Olea europaea and Phyllirea latifolia [34,35]. Although P. bilorbang has been represented for the first time as a causal agent on declining Rubus anglocandicans in Australia [36], it appears that most hosts reported to date are woody plants. The results of the conducted pathogenicity tests showed that the isolated in Bulgaria P. bilorbang (RVel2021/119a) could affect tree species like P. tremula and C. siliquastrum, as well as herbaceous perennial plants such as P. lanceolata and B. perennis. The lack of sufficient information indicates that the identification of the natural hosts of this pathogen is an issue requiring further investigations.
The species P. inundata has been isolated from various plants and water environments in different countries in Europe (UK, France, Denmark, Spain, Italy, Turkey), as well as in some other continents [37,38,39,40,41]. Typical habitats of P. inundata are riparian ecosystems and vegetation areas after flooding. Similarly, in the present investigation, the species was derived after a large-scale spill of water in the area around the estuary of the Veleka River. P. inundata has been associated with diseases of woody plants and shrubs, including genera Aesculus, Olea, Salix, Prunus and Vitis [37]. It has been reported as a causal agent of olive tree decline, and root and collar rot of pomegranate in orchards in Turkey [40,41]. A former investigation of different potential hosts of P. inundata demonstrated that the species is capable of infecting roots of Prunus amygdalus and Beta vulgaris [39]. Additional pathogenicity tests conducted by the same authors with detached twigs of different tree species indicated several dormant hosts including C. siliquastrum, Juglans regia, Magnolia grandiflora, Prunus domestica, Prunus domestica and R. canina. The results of the present study showed that the isolated in Bulgaria P. inundata (RVel2021/119b) induced necrosis on cuttings of C. siliquastrum and R. canina, which confirms the potential of the species to infect these plants.
The species from the genus Phytopythium are mainly described as saprophytic organisms linked to water and soil habitats. However, some members of the group have been reported as dangerous plant pathogens. Pp. litorale has been derived from various types of water ecosystems such as rivers and streams in Austria, Slovakia and the Czech Republic [7] and mountain streams in Vietnam [8], as well as irrigation water tanks in the USA [42]. It has also been identified as a pathogenic species on diverse plant hosts, such as soya bean, almond and oriental plane trees Platanus orientalis [43,44,45]. Similar to previous reports, Pp. litorale (RVel2021/113b) derived from the Veleka River demonstrated aggressiveness to different forest species and perennial and herbaceous plants. Most sensitive were C. siliquastrum, Q. robur, P. tremula, P. quinquefolia and R. canina, which are therefore determined as potential host plants for the pathogen. Similar to Pp. litorale, Pp. citrinum has been isolated from various sources. The species has been identified in streams in the Czech Republic [7], nurseries in Sweden [46] and oak stands in Poland [47,48]. As presented in this study, Pp. citrinum (RVel2021/113a) showed moderate to high potential to infect a number of selected plants and was determined as a pathogen with a wide host range. The collected data suggest that both Phytopythium species are a potential threat for forest ecosystems, gardens and parks.
In this survey, less aggressive against the tested plants were Py. angustatum (RVel2021/114c) and E. anandrum (RVel2021/115d). Py. angustatum is associated with aquatic ecosystems and has been isolated from green algae, freshwater wetland soils invaded by European common reed (P. australis), a river in Ukraine and water courses in Poland [49,50,51,52]. Occurrence of E. anandrum in Europe is most often associated with oak stands, and it has been reported in Germany, Sweden, Austria, Turkey and Poland [48,53,54,55,56,57]. Although E. anandrum (RVel2021/115d) was isolated from the Veleka River by baiting with oak leaves, a low infestation of Q. robur was monitored. A moderate aggressiveness was established only toward herbaceous plants H. helix and T. repens. In addition, Py. angustatum (RVel2021/114c) was weakly pathogenic to Q. robur, as well as most of the other tested plants. However, a moderate pathogenicity against P. lanceolata, B. perennis and A. millefolium was detected. These results indicate that Py. angustatum and E. anandrum are weak pathogens with various potential hosts.
5. Conclusions
Fishing for estuarine oomycetes in the firth region of the Veleka River in Bulgaria resulted in the isolation and identification of eight different species, including P. lacustris, P. bilorbang, P. inundata, P. honggalleglyana, Pp. litorale, Pp. citrinum, Py. angustatum and E. anandrum. A small collection of oomycetes from the genera Phytophthora, Phytopythium, Pythium and Elongisporangium derived from this specific habitat was established. Although a number of Phytophthora species have been reported for the first time in the country during the last few years, two other members of the genus (P. bilorbang and P. inundata) were identified, indicating that surveying various environments leads to the accumulation of new knowledge about oomycete diversity in Bulgaria.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
This study was supported by the project “Characteristics of microorganisms establishing pathogenic and other relationships with plant species from forest and agricultural ecosystems” (2020–2022, contract OL5) of the AgroBioInstitute (Agricultural Academy, Bulgaria). The author gratefully acknowledge R. Batchvarova for her valuable remarks, which significantly contributed to the quality of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schroeder, K.L.; Martin, F.N.; De Cock, A.; Lévesque, C.A.; Spies, C.F.J.; Okubara, P.A.; Paulitz, T.C. Molecular detection and quantification of Pythium species: Evolving taxonomy, new tools, and challenges. Plant Dis. 2013, 97, 4–20. [Google Scholar] [CrossRef]
- Kageyama, K. Molecular taxonomy and its application to ecological studies of Pythium species. J. Gen. Plant Pathol. 2014, 80, 314–326. [Google Scholar] [CrossRef]
- Parke, J.L.; Redekar, N.R.; Eberhart, J.L.; Funahashi, F. Hazard analysis for Phytophthora species in container nurseries: Three Case Studies. HortTechnology 2019, 29, 745–755. [Google Scholar] [CrossRef]
- Thines, M. Phylogeny and evolution of plant pathogenic oomycetes—A global overview. Eur. J. Plant Pathol. 2014, 138, 431–447. [Google Scholar] [CrossRef]
- Oh, E.; Gryzenhout, M.; Wingfield, B.D.; Wingfield, M.J.; Burgess, T.I. Surveys of soil and water reveal a goldmine of Phytophthora diversity in South African natural ecosystems. IMA Fungus 2013, 4, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Nowakowska, J.A.; Oszako, T.; Sikora, K.; Corcobado, T.; Jung, T. Isolation and pathogenicity of Phytophthora species from poplar plantations in Serbia. Forests 2018, 9, 330. [Google Scholar] [CrossRef]
- Corcobado, T.; Cech, T.L.; Daxer, A.; Datkova, H.; Janousek, J.; Patra, S.; Jahn, D.; Huettler, C.; Milenkovic, I.; Tomsovsky, M.; et al. Phytophthora, Nothophytophthora and Halophytophthora diversity in rivers, streams and riparian alder ecosystems of Central Europe. Mycol. Prog. 2023, 22, 50. [Google Scholar] [CrossRef]
- Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenković, I.; Corcobado, T.; Tomšovský, M.; Pánek, M.; Bakonyi, J.; Maia, C.; et al. A Survey in Natural Forest Ecosystems of Vietnam Reveals High Diversity of both New and Described Phytophthora Taxa including P. ramorum. Forests 2020, 11, 93. [Google Scholar] [CrossRef]
- Nagel, J.H.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J.; Hardy, G.E.S.J.; Stukely, M.J.C.; Burgess, T.I. Characterization of Phytophthora hybrids from ITS clade 6 associated with riparian ecosystems in South Africa and Australia. Fungal Biol. 2013, 117, 329–347. [Google Scholar] [CrossRef]
- Yang, X.; Hong, C.X. Phytophthora virginiana sp. nov., a high-temperature tolerant species from irrigation water in Virginia. Mycotaxon 2013, 126, 167–176. [Google Scholar] [CrossRef]
- Christova, P.; Kostov, K.; Lyubenova, A.; Slavov, S. A new hybrid of Phytophthora from Southeast Europe. Mycologia 2021, 113, 734–747. [Google Scholar] [CrossRef]
- Jung, T.; Stukely, M.J.C.; Hardy, G.E.S.J.; White, D.; Paap, T.; Dunstan, W.A.; Burgess, T.I. Multiple new Phytophthora species from ITS Clade 6 associated with natural ecosystems. Persoonia 2011, 26, 13–39. [Google Scholar] [CrossRef] [PubMed]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gallegly, M.E.; Hong, C. A high-temperature tolerant species in clade 9 of the genus Phytophthora: P. hydrogena sp. nov. Mycologia 2014, 106, 57–65. [Google Scholar] [CrossRef]
- Stamler, R.A.; Sanogo, S.; Goldberg, N.P.; Randall, J.J. Phytophthora Species in Rivers and Streams of the Southwestern United States. Appl. Environ. Microbiol. 2016, 82, 4696–4704. [Google Scholar] [CrossRef]
- Pintos, C.; Rial, C.; Aguín, O.; Ferreiroa, V.; Mansilla, J.P. First report of Phytophthora hydropathica in river water associated with riparian alder in Spain. New Dis. Rep. 2016, 33, 25. [Google Scholar] [CrossRef]
- Thines, M.; Kamoun, S. Oomycete-plant coevolution: Recent advances and future prospects. Curr. Opin. Plant Biol. 2010, 13, 1e7. [Google Scholar] [CrossRef]
- Redekar, N.R.; Eberhart, J.L.; Parke, J.L. Diversity of Phytophthora, Pythium, and Phytopythium Species in Recycled Irrigation Water in a Container Nursery. Phytobiomes J. 2019, 3, 31–45. [Google Scholar] [CrossRef]
- Govers, L.L.; Man in ‘t Veld, W.A.; Meffert, J.P.; Bouma, T.J.; van Rijswick, P.C.J.; Heusinkveld, J.H.T.; Orth, R.J.; van Katwijk, M.M.; van der Heide, T. Marine Phytophthora species can hamper conservation and restoration of vegetated coastal ecosystems. Proc. R. Soc. B 2016, 283, 20160812. [Google Scholar] [CrossRef]
- Bennett, R.M.; Thines, M. Confirmation that Phytophthora insolita (Peronosporaceae) is present as a marine saprotroph on mangrove leaves and first report of the species for the Philippines. Nova Hedwig. Band 2017, 105, 185–196. [Google Scholar] [CrossRef]
- Bennett, R.M.; Thines, M. An overview on Philippine estuarine oomycetes. Philipp. J. Syst. Biol. 2020, 14, 1–14. [Google Scholar]
- Kostov, K.; Christova, P.; Lyubenova, A.; Slavov, S. Diversity of oomycete species in the riparian ecosystem of Iskar gorge. Annual of Sofia University “St. Kliment Ohridski”. In Book 4—Scientific Sessions of the Faculty of Biology; Sofia University: Sofia, Bulgaria, 2019; Volume 104, pp. 90–99. [Google Scholar]
- Christova, P.K.; Lyubenova, A.B.; Kostov, K.V.; Slavov, S.B. First report of Phytophthora pseudosyringae recovered from aquatic ecosystems in Bulgaria. For. Pathol. 2019, 49, e12505. [Google Scholar] [CrossRef]
- Christova, P.K. Detection of Phytophthora gallica in Bulgaria and co-existence with other Phytophthora species in a small river. J. Plant Dis. Prot. 2022, 129, 1377–1387. [Google Scholar] [CrossRef]
- Christova, P.K. Phytophthora polonica and Phytophthora hydropathica from clade 9 associated with alder decline in Bulgaria. Life 2024, 14, 720. [Google Scholar] [CrossRef] [PubMed]
- Slavov, S.; Lubenova, A.; Kostov, K.; Christova, P. Oomycotes from genus Pythium found in water and soil samples in Bulgaria. J. Mt. Agric. Balk. 2018, 21, 161–171. [Google Scholar]
- Yordanova, E. First report of root rot caused by Pythium aphanidermatum on industrial hemp (Cannabis sativa L.) in Bulgaria. Agric. Sci. 2023, 15, 83–88. [Google Scholar] [CrossRef]
- Vatchev, T.D.; Bistrichanov, S.M.; Yanashkov, I.T. First report of root rot caused by Pythium aphanidermatum and Pythium F-group on hydroponically grown pepper in Bulgaria. Bulg. J. Agric. Sci. 2016, 22, 794–795. [Google Scholar]
- Nechwatal, J.; Bakonyi, J.; Cacciola, S.O.; Cooke, D.E.L.; Jung, T.; Nagy, Z.A.; Vannini, A.; Vettraino, A.M.; Brasier, C.M. The morphology, behavior and molecular phylogeny of Phytophthora taxon Salixsoil and its redestination as Phytophthora lacustris sp. nov. Plant Pathol. 2013, 62, 355–369. [Google Scholar] [CrossRef]
- Vitale, S.; Luongo, L.; Galli, M.; Belisario, A. A First Report of Phytophthora hydropathica Causing Wilting and Shoot Dieback on Viburnum in Italy. Plant Dis. 2014, 98, 1582. [Google Scholar] [CrossRef]
- Bregant, C.; Sanna, G.P.; Bottos, A.; Maddau, L.; Montecchio, L.; Linaldeddu, B.T. Diversity and pathogenicity of Phytophthora species associated with declining alder trees in Italy and description of Phytophthora alpina sp. nov. Forests 2020, 11, 84. [Google Scholar] [CrossRef]
- Hong, C.X.; Gallegly, M.E.; Richardson, P.A.; Kong, P.; Moorman, G.W.; Lea-Cox, J.D.; Ross, D.S. Phytophthora hydropathica, a new pathogen identified from irrigation water, Rhododendron catawbiense and Kalmia latifolia. Plant Pathol. 2010, 59, 913–921. [Google Scholar] [CrossRef]
- La Spada, F.; Cock, P.J.A.; Randall, E.; Pane, A.; Cooke, D.E.L.; Cacciola, S.O. DNA Metabarcoding and Isolation by Baiting Complement Each Other in Revealing Phytophthora Diversity in Anthropized and Natural Ecosystems. J. Fungi 2022, 8, 330. [Google Scholar] [CrossRef]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First Report of Root Rot Caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef] [PubMed]
- Bregant, C.; Mulas, A.A.; Rossetto, G.; Deidda, A.; Maddau, L.; Piras, G.; Linaldeddu, B.T. Phytophthora mediterranea sp. nov., a New Species Closely Related to Phytophthora cinnamomi from Nursery Plants of Myrtus communis in Italy. Forests 2021, 12, 682. [Google Scholar] [CrossRef]
- Aghighi, S.; Hardy, G.E.S.J.; Scott, J.K.; Burgess, T.I. Phytophthora bilorbang sp. nov., a new species associated with the decline of Rubus anglocandicans (European blackberry) in Western Australia. Eur. J. Plant Pathol. 2012, 133, 841–855. [Google Scholar] [CrossRef]
- Brasier, C.M.; Sanchez-Hernandez, E.; Kirk, S.A. Phytophthora inundata sp. nov., a part heterothallic pathogen of trees and shrubs in wet or flooded soils. Mycol. Res. 2003, 107, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.H.; Hong, C.X.; Erwin, D.C. Phytophthora inundata isolated from diseased alfalfa roots in southern California. Mycotaxon 2006, 97, 349–358. [Google Scholar]
- Safaiefarahani, B.; Mostowfizadeh-Ghalamfarsa, R.; Cooke, D.E.L. Characterisation of Phytophthora inundata according to host range, morphological variation and multigene molecular phylogeny. Phytopathol. Mediterr. 2013, 52, 46–65. [Google Scholar]
- Kurbetli, İ.; Sülü, G.; Taştekin, E.; Polat, İ. First report of Phytophthora inundata causing olive tree decline in Turkey. Can. J. Plant Pathol. 2016, 38, 254–257. [Google Scholar] [CrossRef]
- Kurbetli, İ.; Karaca, G.; Aydoğdu, M.; Sülü, G. Phytophthora Species Causing Root and Collar Rot of Pomegranate in Turkey. Eur. J. Plant Pathol. 2020, 157, 485–496. [Google Scholar] [CrossRef]
- Choudhary, C.E.; Burgos-Garay, M.L.; Moorman, G.W.; Hong, C. Pythium and Phytopythium species in two Pennsylvania greenhouse irrigation water tanks. Plant Dis. 2016, 100, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Radmer, L.; Anderson, G.; Malvick, D.M.; Kurle, J.E.; Rendahl, A.; Mallik, A. Pythium, Phytophthora, and Phytopythium spp. associated with soybean in Minnesota, their relative aggressiveness on soybean and corn, and their sensitivity to seed treatment fungicides. Plant Dis. 2017, 101, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Javadi, N.; Sharifnabi, B. Phytopythium litorale, the causal agent of almond root and crown rot in Iran. In Proceedings of the 22nd Iranian Plant Protection Congress, College of Agriculture and Natural Resources, University of Tehran, Karaj, Iran, 27–30 August 2016. [Google Scholar]
- Derviş, S.; Turkolmez, Ş.; Ciftci, O.; Ozer, G.; Serce, C.U.; Dikilitas, M. Phytopythium littorale: A novel killer pathogen of plane (Platanus orientalis) causing canker stain and root and collar rot. Plant Dis. 2020, 104, 2642–2648. [Google Scholar] [CrossRef] [PubMed]
- Cleary, M.; Blomquist, M.; Marchand, M.; Witzell, J. Oomycetes in rhizosphere soil of ornamental plants from retail nurseries in Southern Sweden. For. Pathol. 2021, 51, e12673. [Google Scholar] [CrossRef]
- Cordier, T.; Belbahri, L.; Calmin, G.; Oszako, T.; Nowakowska, J.; Lefort, F. Emerging Phytophthora and Pythium species in Polish declining forests. Established and emerging Phytophthora: Increasing threats to woodland and forest ecosystems in Europe. In Proceedings of the First Working Groups Meeting, Nový Smokovec, Slovakia, 16–19 April 2009. [Google Scholar]
- Jankowiak, R.; Stępniewska, H.; Bilański, P. Notes on some Phytopythium and Pythium species occurring in oak forests in southern Poland. Acta Mycologyca 2015, 50, 1052. [Google Scholar] [CrossRef]
- Sparrow, F.K. Two new species of Pythium parasitic in green algae. Ann. Bot. 1931, 45, 257–277. [Google Scholar] [CrossRef]
- Crocker, E.V.; Karp, M.A.; Nelson, E.B. Virulence of oomycete pathogens from Phragmites australis-invaded and noninvaded soils to seedlings of wetland plant species. Ecol. Evol. 2015, 5, 2127–2139. [Google Scholar] [CrossRef]
- Matsiakh, I.; Oszako, T.; Kramarets, V.; Nowakowska, J.A. Phytophthora and Pythium Species Detected in Rivers of the Polish-Ukrainian Border Areas. Balt. For. 2016, 22, 230–238. [Google Scholar]
- Oszako, T.; Sikora, K.; Belbahr, L.; Nowakowska, J.A. Molecular detection of oomycetes species in water courses. Folia For. Pol. Ser. A For. 2016, 58, 246–251. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phythophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Jönsson, U.; Lundberg, L.; Sonesson, K.; Jung, T. First records of soilborne Phytophthora species in Swedish oak forests. For. Pathol. 2003, 33, 175–179. [Google Scholar] [CrossRef]
- Balcì, Y.; Halmschlager, E. Incidence of Phytophthora species in oak forests in Austria and their possible involvement in oak decline. For. Pathol. 2003, 33, 157–174. [Google Scholar] [CrossRef]
- Balcì, Y.; Halmschlager, E. Phytophthora species in oak ecosystems in Turkey and their association with declining oak trees. Plant Pathololy 2003, 52, 694–702. [Google Scholar] [CrossRef]
- Akıllı, S.; Serçe, Ç.U.; Katırcıoğlu, Y.Z.; Maden, S. Does Pythium anandrum contribute to the dieback of sessile oak (Quercus petraea) in Turkey? For. Pathol. 2013, 43, 505–508. [Google Scholar] [CrossRef]
Figure 1.
Sampling area of the Veleka River: (A) map of the region and the studied sites; (B) photo of the river.
Figure 1.
Sampling area of the Veleka River: (A) map of the region and the studied sites; (B) photo of the river.

Table 2.
Pathogenicity of the tested isolates Phytophthora, Phytopythium, Pythium and Elongisporangium on selected plant species.
Table 2.
Pathogenicity of the tested isolates Phytophthora, Phytopythium, Pythium and Elongisporangium on selected plant species.
| Plant Species | Phytophthora | Phytopythium | Pythium | Elongisporangium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 113d/PL | 119a/PB | 119b/PI | 120c/PH | 113a/PpC | 113b/PpL | 114c/PyAg | 115d/EAn | |||||||||
| Leaves/Cuttings | L | C | L | C | L | C | L | C | L | C | L | C | L | C | L | C |
| C. siliquastrum | +++ | +++ | + | ++ | - | + | +++ | +++ | + | +++ | ++ | +++ | - | + | - | + |
| Q. robur | +++ | +++ | + | + | + | + | +++ | +++ | ++ | +++ | +++ | ++ | + | + | + | + |
| P. tremula | +++ | +++ | ++ | ++ | - | - | +++ | +++ | +++ | +++ | +++ | +++ | - | - | + | + |
| S. babylonica | +++ | - | - | + | - | - | + | - | + | - | - | + | - | - | - | - |
| R. canina | +++ | +++ | - | - | - | + | +++ | +++ | + | ++ | ++ | +++ | - | - | + | - |
| R. fruticosus | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| H. helix | - | - | - | - | - | - | ++ | - | - | - | - | - | - | - | ++ | - |
| P. quinquefolia | +++ | + | + | - | ++ | - | ++ | ++ | +++ | ++ | +++ | + | + | - | - | - |
| C. vitalba | ++ | + | - | - | + | - | ++ | - | ++ | - | ++ | - | - | - | - | - |
| M. sativa | - | - | - | - | - | - | +++ | ++ | ++ | - | ++ | - | + | - | + | - |
| T. repens | + | na | + | na | ++ | na | +++ | na | - | na | ++ | na | - | na | ++ | na |
| P. lanceolata | - | na | ++ | na | +++ | na | + | na | ++ | na | - | na | ++ | na | - | na |
| P. major | - | na | - | na | - | na | + | na | - | na | - | na | - | na | - | na |
| B. perennis | ++ | na | ++ | na | + | na | ++ | na | - | na | ++ | na | ++ | na | - | na |
| A. millefolium | ++ | na | + | na | ++ | na | + | na | ++ | na | ++ | na | ++ | na | - | na |
| S. viridis | ++ | na | - | na | + | na | - | na | ++ | na | + | na | - | na | + | na |
PL—P. lacustris; PB—P. bilorbang; PI—P. inundata; PH—P. honggalleglyana; PpC—Pp. citrinum; PpL—Pp. litorale; PyAg—Py. angustatum; EAn—E. anandrum. Disease symptoms severity scale: - no symptoms; + small dot necroses; ++ necrotic area about 50% of the leaf/cutting; +++ necrosis covers 90–100% of the leaf/cutting; na—not applicable. L—leaves; C—cuttings.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Christova, P.K. Fishing for Estuarine Oomycetes. Diversity 2024, 16, 530. https://doi.org/10.3390/d16090530
AMA Style
Christova PK. Fishing for Estuarine Oomycetes. Diversity. 2024; 16(9):530. https://doi.org/10.3390/d16090530
Chicago/Turabian StyleChristova, Petya Koeva. 2024. "Fishing for Estuarine Oomycetes" Diversity 16, no. 9: 530. https://doi.org/10.3390/d16090530
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.