

Two-Decade Changes in the Ciliate Assemblage Feeding Pattern Reflect the Reservoir Nutrient Load

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Limnology Variables, Sampling, and Plankton Analysis
2.3. Ciliates
2.4. Data Treatment
3. Results
3.1. Long-Term Trends in the Reservoir
3.1.1. Physical–Chemical Variables
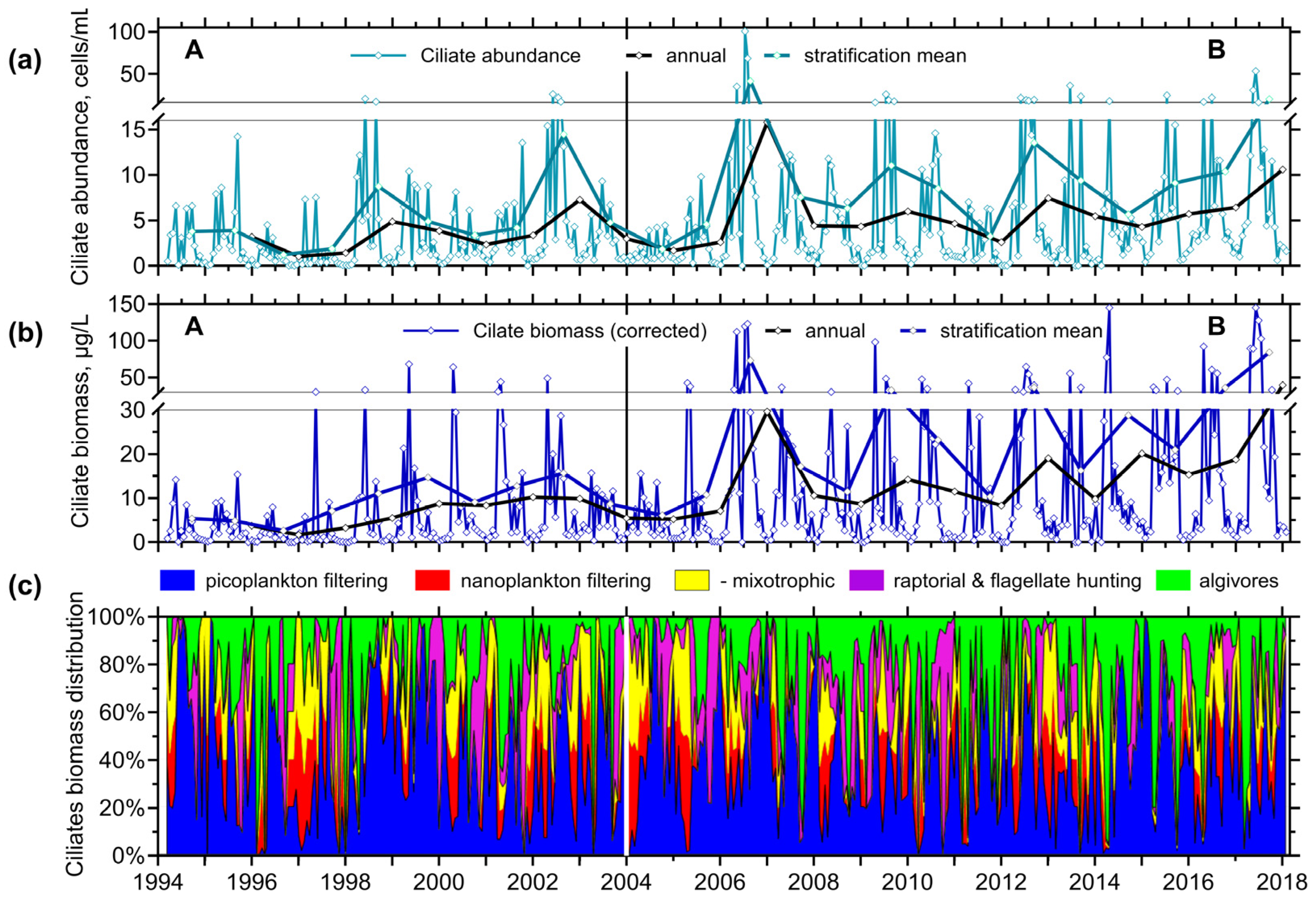
3.1.2. Plankton
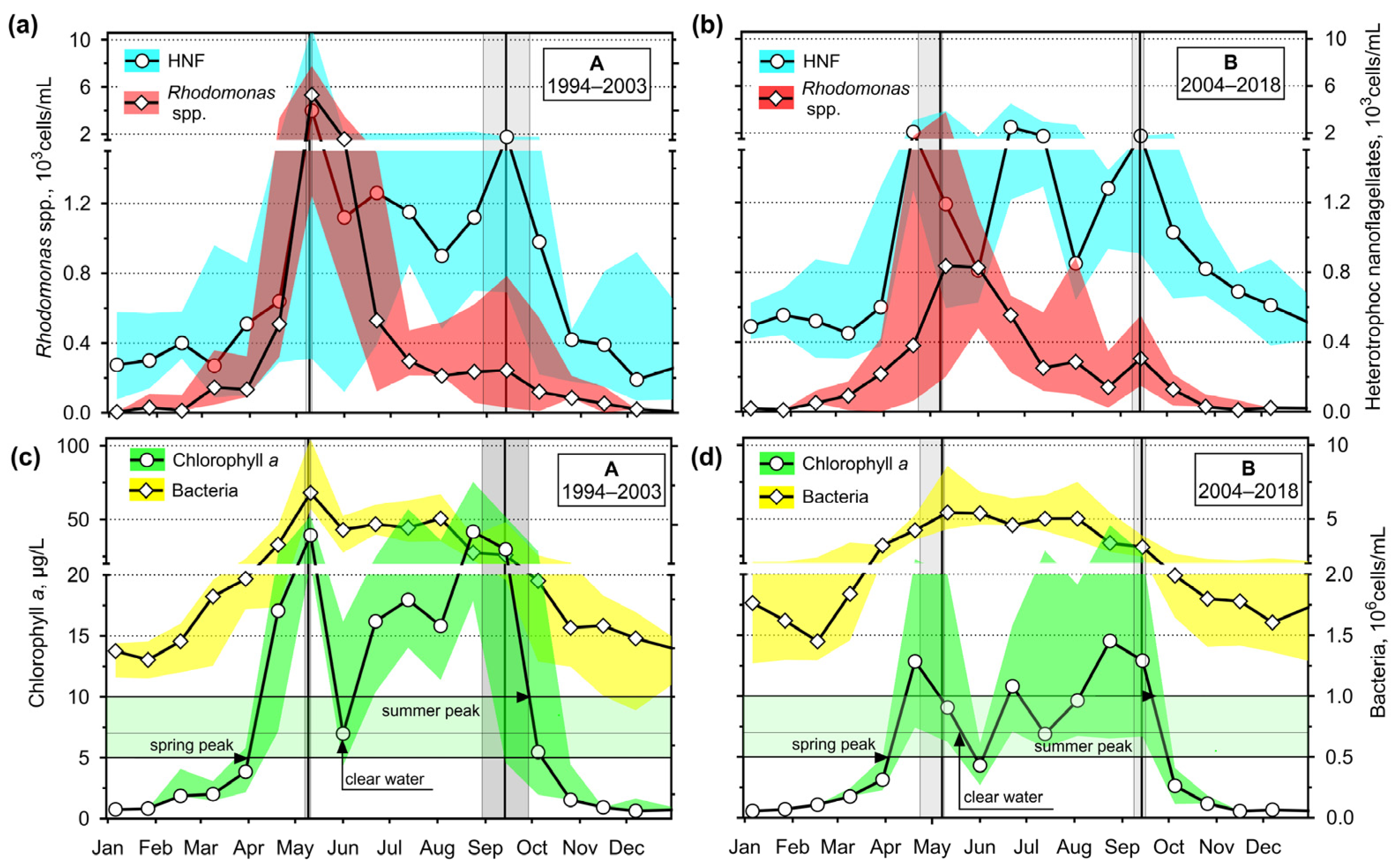
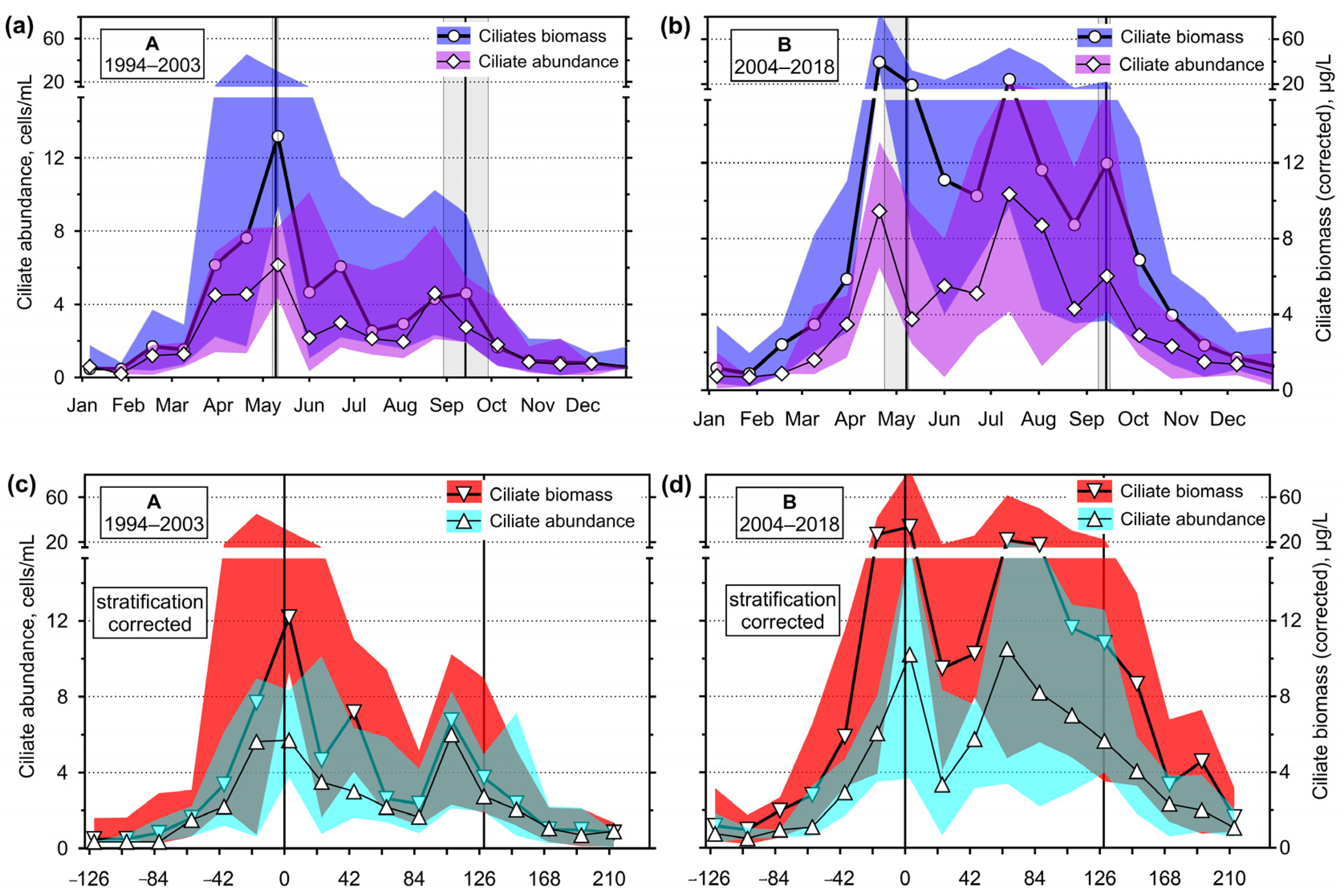
3.2. Seasonality
3.2.1. Physical–Chemical Variables
3.2.2. Chlorophyll a, Bacteria, Rhodomonas spp., and Heterotrophic Nanoflagellates
3.2.3. Ciliates’ Assemblage Abundance and Biomass
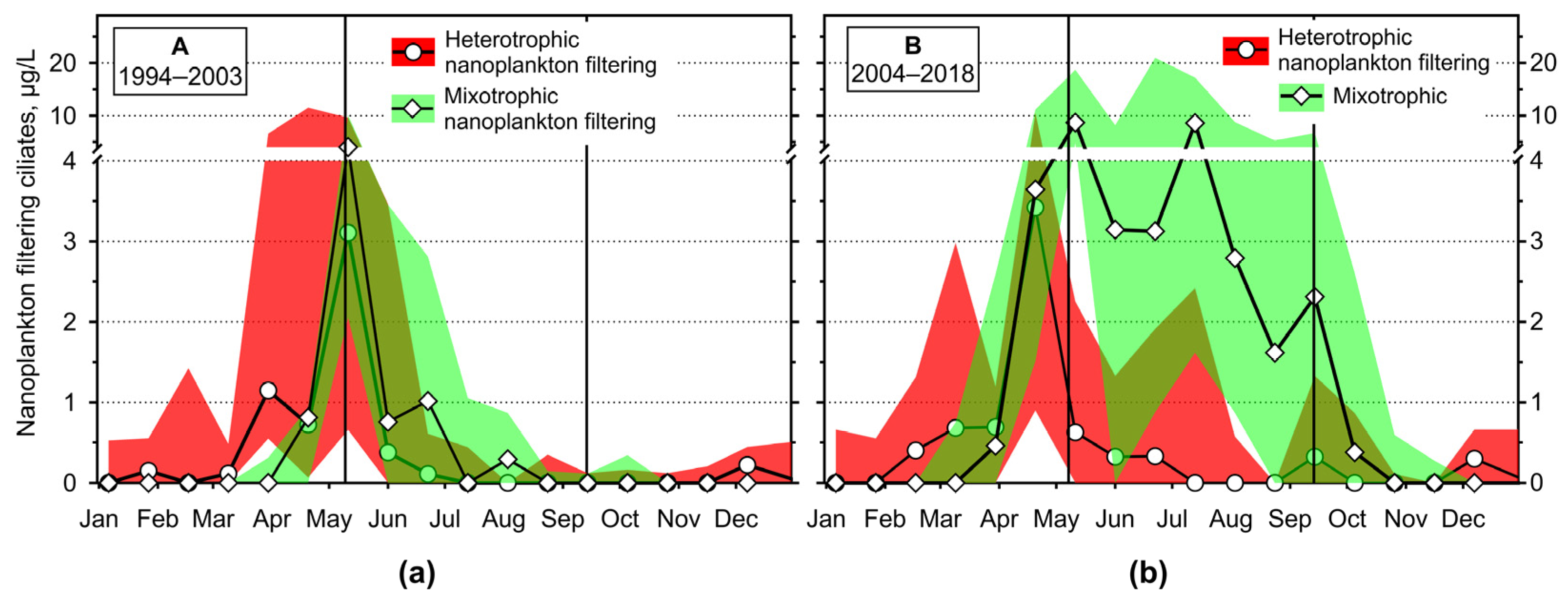
3.2.4. Annual Changes in the Ciliate Feeding-Behaviour Groups
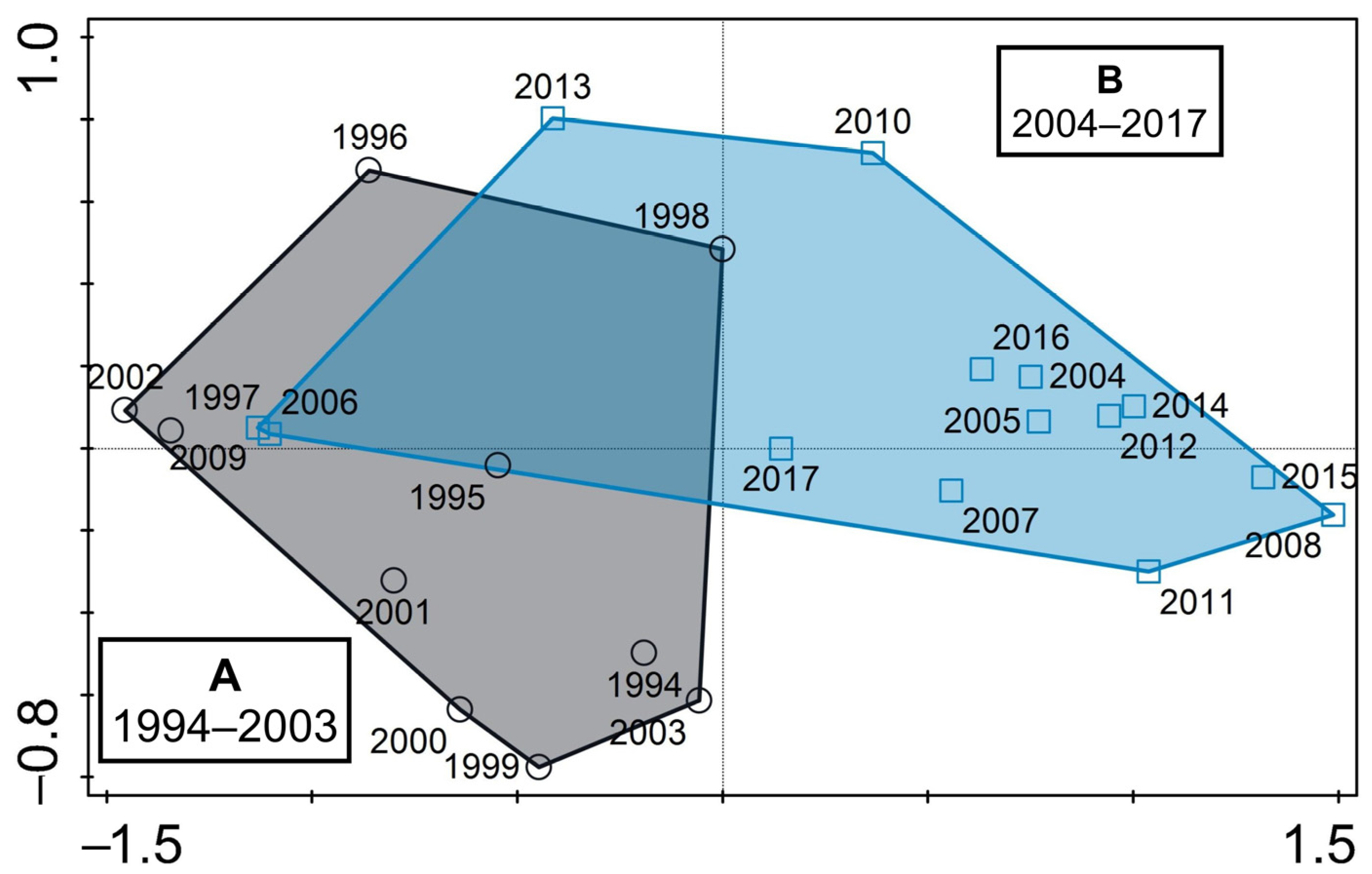
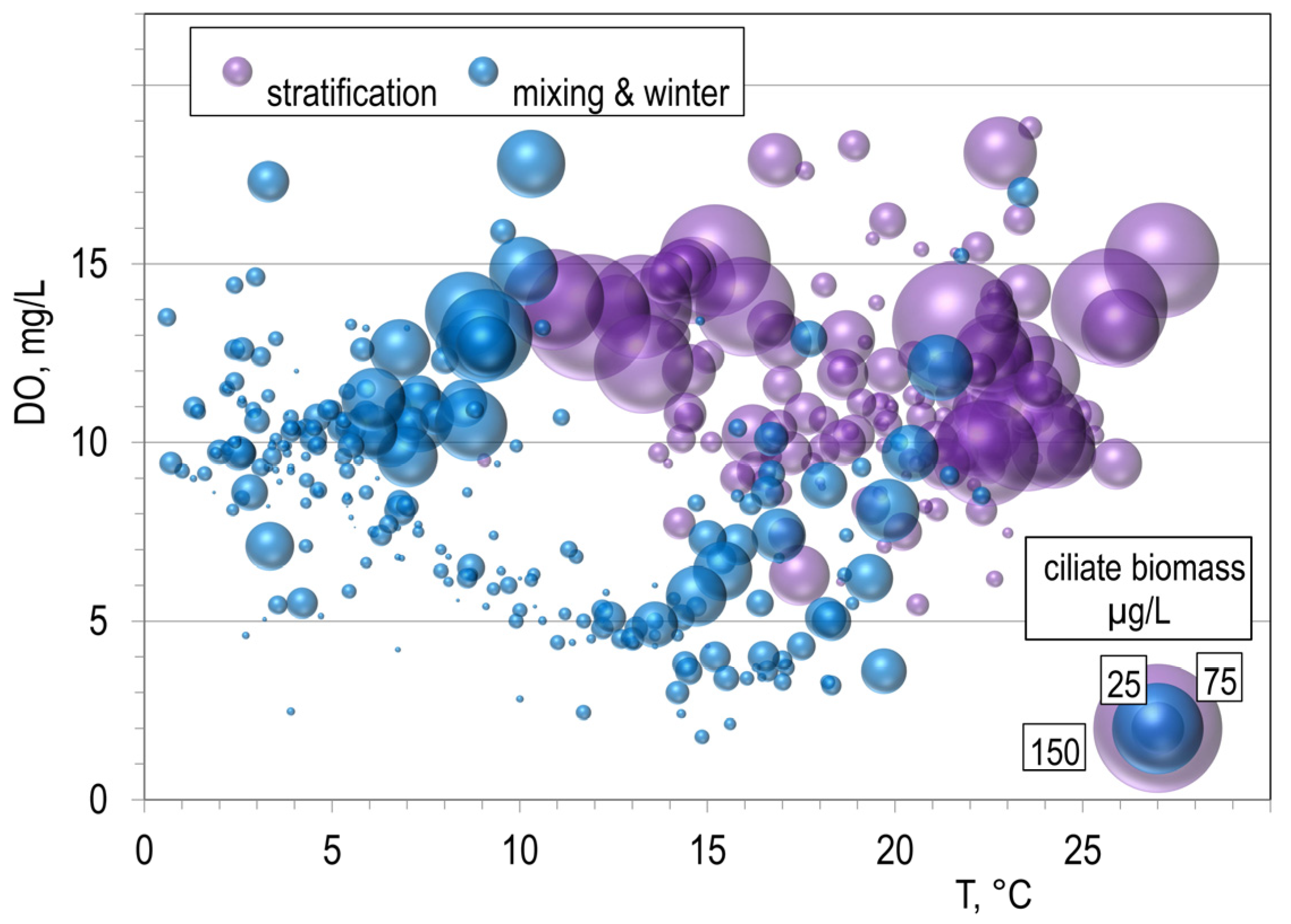
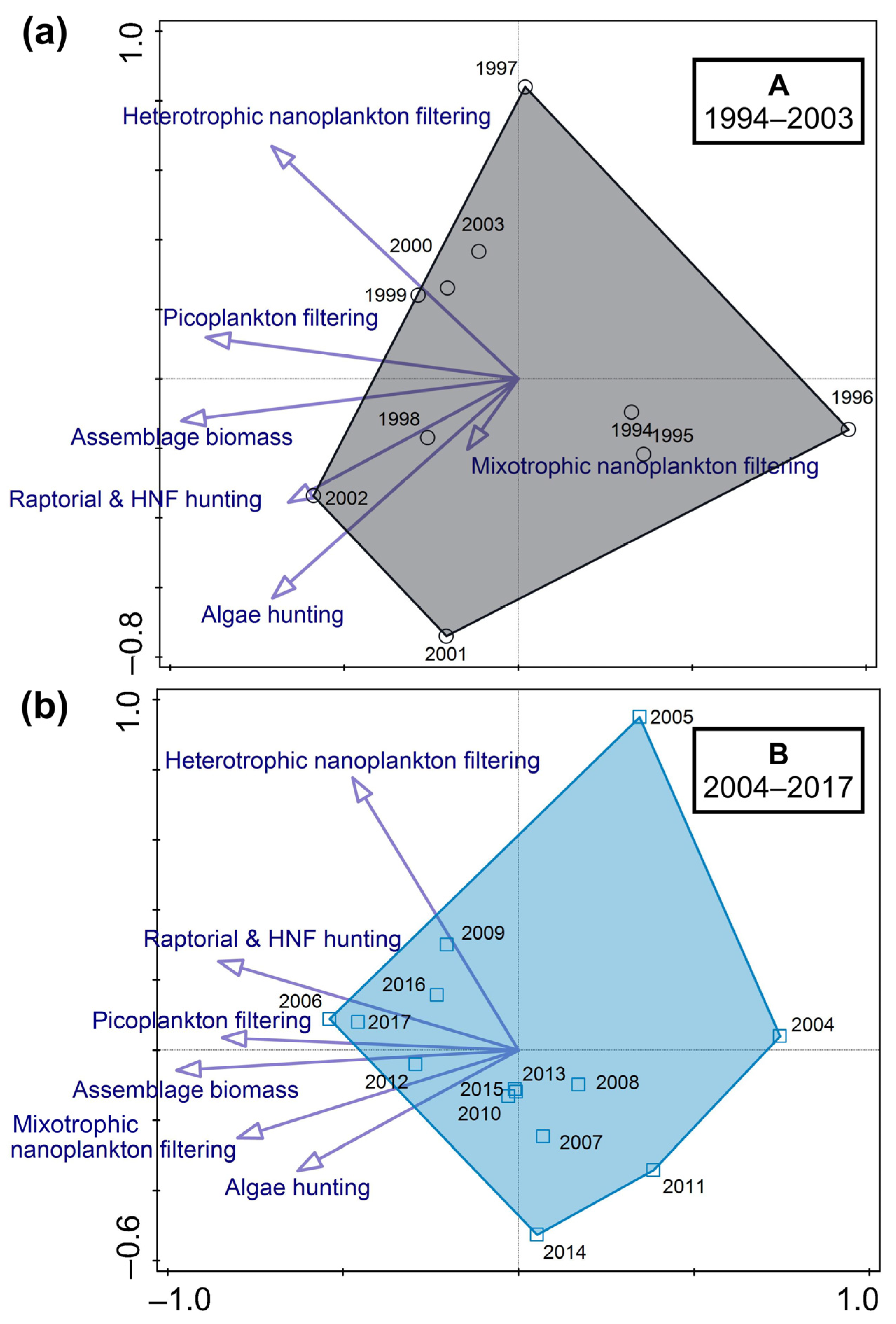
3.3. Predator–Prey Relation and the Importance of Different Feeding-Behaviour Patterns within Ciliates
4. Discussion
4.1. Hydrodynamics of the Surface Layer and a Legacy of Defined Periods A and B
4.2. Environmental Ciliates’ Identification
4.3. Ciliate Ecology in the Reservoir
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sommer, U.; Adrian, R.; De Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lürling, M.; Molinero, J.C.; Mooij, W.M.; et al. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The Use of DAPI for Identifying and Counting Aquatic Microflora1. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.; Gray, J.; Meyer-Reil, L.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Weisse, T.; Montagnes, D.J.S. Ecology of Planktonic Ciliates in a Changing World: Concepts, Methods, and Challenges. J. Eukaryot. Microbiol. 2022, 69, e12879. [Google Scholar] [CrossRef]
- Posch, T.; Eugster, B.; Pomati, F.; Pernthaler, J.; Pitsch, G.; Eckert, E.M. Network of Interactions Between Ciliates and Phytoplankton During Spring. Front. Microbiol. 2015, 6, 1289. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F. Protistan Grazing Rates via Uptake of Fluorescently Labeled Prey. In Handbook of Methods on Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publishers: London, UK, 1993; pp. 695–701. [Google Scholar]
- Sherr, E.B.; Sherr, B.F.; Berman, T.; Hadas, O. High Abundance of Picoplankton-Ingesting Ciliates during Late Fall in Lake Kinneret, Israel. J. Plankton Res. 1991, 13, 789–799. [Google Scholar] [CrossRef]
- Gifford, D.J. Consumption of Protozoa by Copepods Feeding on Natural Microplankton Assemblages. In Handbook of Methods on Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publishers: London, UK, 1993; pp. 723–729. [Google Scholar]
- Stoecker, D.; Pierson, J. Predation on Protozoa: Its Importance to Zooplankton Revisited. J. Plankton Res. 2019, 41, 367–373. [Google Scholar] [CrossRef]
- Hecky, R.E.; Kling, H.J. The Phytoplankton and Protozooplankton of the Euphotic Zone of Lake Tanganyika: Species Composition, Biomass, Chlorophyll Content, and Spatio-temporal Distribution1. Limnol. Oceanogr. 1981, 26, 548–564. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Gifford, D.J.; Putt, M. Preservation of Marine Planktonic Ciliates: Losses and Cell Shrinkage during Fixation. Mar. Ecol. Prog. Ser. 1994, 110, 293–299. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F. Preservation and Storage of Samples for Enumeration of Heterotrophic Protists. In Handbook of Methods on Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publishers: London, UK, 1993; pp. 207–212. [Google Scholar]
- Taylor, W.D.; Heynen, M.L. Seasonal and Vertical Distribution of Ciliophora in Lake Ontario. Can. J. Fish. Aquat. Sci. 1987, 44, 2185–2191. [Google Scholar] [CrossRef]
- Madoni, P. The Ciliated Protozoa of the Monomictic Lake Kinneret (Israel): Species Composition and Distribution during Stratification. Hydrobiologia 1990, 190, 111–120. [Google Scholar] [CrossRef]
- Straškrabová, V.; Callieri, C.; Carrillo, P.; Cruz-Pizarro, L.; Fott, J.; Hartman, P.; Macek, M.; Medina-Sánchez, J.M.; Nedoma, J.; Šimek, K. Investigations on Pelagic Food Webs in Mountain Lakes—Aims and Methods. J. Limnol. 1999, 58, 77–87. [Google Scholar] [CrossRef]
- Felip, M.; Bartumeus, F.; Halac, S.; Catalan, J. Microbial Plankton Assemblages, Composition and Biomass, during Two Ice-Free Periods in a Deep High Mountain Lake (Estany Redó, Pyrenees). J. Limnol. 1999, 58, 193–202. [Google Scholar] [CrossRef]
- Czychewicz, N.; Rychert, K. Seasonal Changes in Ciliate Biomass and Composition of the Ciliate Community in Oligo-Mesotrophic Lake Jasne (Iława Lake District, Poland). Limnol. Rev. 2011, 11, 3–5. [Google Scholar] [CrossRef]
- Müller, H. The Relative Importance of Different Ciliate Taxa in the Pelagic Food Web of Lake Constance. Microb. Ecol. 1989, 18, 261–273. [Google Scholar] [CrossRef]
- Gaedke, U.; Wickham, S.A. Ciliate Dynamics in Response to Changing Biotic and Abiotic Conditions in a Large, Deep Lake (Lake Constance). Aquat. Microb. Ecol. 2004, 34, 247–261. [Google Scholar] [CrossRef]
- Hadas, O.; Berman, T. Seasonal Abundance and Vertical Distribution of Protozoa (Flagellates, Ciliates), and Bacteria in Lake Kinneret, Israel. Aquat. Microb. Ecol. 1998, 14, 161–170. [Google Scholar] [CrossRef]
- Šimek, K.; Bobková, J.; Macek, M.; Nedoma, J.; Psenner, R. Ciliategrazing on Picoplankton in a Eutrophic Reservoir during the Summer Phytoplankton Maximum: A Study at the Species and Community Level. Limnol. Oceanogr. 1995, 40, 1077–1090. [Google Scholar] [CrossRef]
- Peštová, D.; Macek, M.; Martínez-Pérez, M.E. Ciliates and Their Picophytoplankton-Feeding Activity in a High-Altitude Warm-Monomictic Saline Lake. Eur. J. Protistol. 2008, 44, 13–25. [Google Scholar] [CrossRef]
- Bautista-Reyes, F.; Macek, M. Ciliate Food Vacuole Content and Bacterial Community Composition in the Warm-Monomictic Crater Lake Alchichica, México. FEMS Microbiol. Ecol. 2012, 79, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Montagnes, D.J.S.; Lynn, D.H. A Quantitative Protargol Stain (QPS) for Ciliates and Other Protists. In Handbook of Methods on Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publishers: London, UK, 1993; pp. 229–240. [Google Scholar]
- Skibbe, O. An Improved Quantitative Protargol Stain for Ciliates and Other Planktonic Protists. Arch. Hydrobiol. 1994, 130, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Pfister, G.; Sonntag, B.; Posch, T. Comparison of a Direct Live Count and an Improved Quantitative Protargol Stain (QPS) in Determining Abundance and Cell Volumes of Pelagic Freshwater Protozoa. Aquat. Microb. Ecol. 1999, 18, 95–103. [Google Scholar] [CrossRef]
- Sonntag, B.; Posch, T.; Klammer, S.; Griebler, C.; Psenner, R. Protozooplankton in the Deep Oligotrophic Traunsee (Austria) Influenced by Discharges of Soda and Salt Industries. Water Air Soil Pollut. Focus 2002, 2, 211–226. [Google Scholar] [CrossRef]
- Sonntag, B.; Posch, T.; Klammer, S.; Teubner, K.; Psenner, R. Phagotrophic Ciliates and Flagellates in an Oligotrophic, Deep, Alpine Lake: Contrasting Variability with Seasons and Depths. Aquat. Microb. Ecol. 2006, 43, 193–207. [Google Scholar] [CrossRef]
- Macek, M.; Callieri, C.; Šimek, K.; Lugo-Vázquez, A. Seasonal Dynamics, Composition and Feeding Patterns of Ciliate Assemblages in Oligotrophic Lakes Covering a Wide pH Range. Arch. Hydrobiol. 2006, 166, 261–287. [Google Scholar] [CrossRef]
- Sánchez Medina, X.; Macek, M.; Bautista-Reyes, F.; Perz, A.; Bonilla Lemus, P.; Chávez Arteaga, M. Inter-Annual Ciliate Distribution Variation within the Late Stratification Oxycline in a Monomictic Lake, Lake Alchichica (Mexico). J. Limnol. 2016, 75, 179–190. [Google Scholar] [CrossRef]
- Pitsch, G.; Bruni, E.P.; Forster, D.; Qu, Z.; Sonntag, B.; Stoeck, T.; Posch, T. Seasonality of Planktonic Freshwater Ciliates: Are Analyses Based on V9 Regions of the 18S rRNA Gene Correlated with Morphospecies Counts? Front. Microbiol. 2019, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Dirren-Pitsch, G.; Bühler, D.; Salcher, M.M.; Bassin, B.; Le Moigne, A.; Schuler, M.; Pernthaler, J.; Posch, T. FISHing for Ciliates: Catalyzed Reporter Deposition Fluorescence in Situ Hybridization for the Detection of Planktonic Freshwater Ciliates. Front. Microbiol. 2022, 13, 1070232. [Google Scholar] [CrossRef]
- Finlay, B.J.; Esteban, G.F. Planktonic Ciliate Species Diversity as an Integral Component of Ecosystem Function in a Freshwater Pond. Protist 1998, 149, 155–165. [Google Scholar] [CrossRef]
- Macek, M. Distribution of Ciliates in the Římov Reservoir. Ergeb. Limnol. 1994, 40, 137–141. [Google Scholar]
- Qu, Z.; Forster, D.; Bruni, E.P.; Frantal, D.; Kammerlander, B.; Nachbaur, L.; Pitsch, G.; Posch, T.; Pröschold, T.; Teubner, K.; et al. Aquatic Food Webs in Deep Temperate Lakes: Key Species Establish through Their Autecological Versatility. Mol. Ecol. 2021, 30, 1053–1071. [Google Scholar] [CrossRef] [PubMed]
- Abdullah Al, M.; Wang, W.; Jin, L.; Chen, H.; Xue, Y.; Jeppesen, E.; Majaneva, M.; Xu, H.; Yang, J. Planktonic Ciliate Community Driven by Environmental Variables and Cyanobacterial Blooms: A 9-Year Study in Two Subtropical Reservoirs. Sci. Total Environ. 2023, 858, 159866. [Google Scholar] [CrossRef] [PubMed]
- D’Alelio, D.; Libralato, S.; Wyatt, T.; Ribera d’Alcalà, M. Ecological-Network Models Link Diversity, Structure and Function in the Plankton Food-Web. Sci. Rep. 2016, 6, 21806. [Google Scholar] [CrossRef] [PubMed]
- Weisse, T.; Müller, H.; Pinto-Coelho, R.M.; Schweizer, A.; Springmann, D.; Baldringer, G. Response of the Microbial Loop to the Phytoplankton Spring Bloom in a Large Prealpine Lake. Limnol. Oceanogr. 1990, 35, 781–794. [Google Scholar] [CrossRef]
- Tirok, K.; Gaedke, U. Regulation of Planktonic Ciliate Dynamics and Functional Composition during Spring in Lake Constance. Aquat. Microb. Ecol. 2007, 49, 87–100. [Google Scholar] [CrossRef]
- Kerimoglu, O.; Straile, D.; Peeters, F. Modeling the Spring Blooms of Ciliates in a Deep Lake. Hydrobiologia 2014, 731, 173–189. [Google Scholar] [CrossRef]
- Kath, N.J.; Thomas, M.K.; Gaedke, U. Mysterious Ciliates: Seasonally Recurrent and yet Hard to Predict. J. Plankton Res. 2022, 44, 891–910. [Google Scholar] [CrossRef]
- Hadas, O.; Berman, T.; Malinsky-Rushansky, N.; Gal, G. Protozoa (Unicellular Zooplankton): Ciliates and Flagellates. In Lake Kinneret; Zohary, T., Sukenik, A., Berman, T., Nishri, A., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 247–258. ISBN 978-94-017-8943-1. [Google Scholar]
- Nishino, M. Biodiversity of Lake Biwa. In Lake Biwa: Interactions between Nature and People; Kawanabe, H., Nishino, M., Maehata, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 69–257. ISBN 978-3-030-16968-8. [Google Scholar]
- Timoshkin, O.; Grygier, M.J.; Wada, E.; Nakai, K.; Nishino, M.; Genkal, S.I.; Biserov, V.I.; Gagarin, V.G.; Semernoy, V.P.; Jankowski, A.W.; et al. Biodiversity of Lake Biwa: New Discoveries and Future Potential. In Annotirovannyi Spisok Fauny Ozera Baikal i Ego Vodosbornogo Basseina [Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area]; Timoshkin, O., Ed.; Nauka: Novosibirsk, Russia, 2009; Volume 1, pp. 1439–1513. [Google Scholar]
- Macek, M.; Sánchez-Medina, X.; Vilaclara, G.; Lugo-Vázquez, A.; Bautista-Reyes, F.; Valdespino-Castillo, P.M. Protozooplankton. In Lake Alchichica Limnology; Alcocer, J., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 213–236. ISBN 978-3-030-79095-0. [Google Scholar]
- Hejzlar, J.; Vrba, J. Brief History of Long-Term Ecological Research into Aquatic Ecosystems and Their Catchments in the Czech Republic; Znachor, P., Ed.; Biology Centre, Institute of hydrobiology: České Budějovice, Czech Republic, 2016; ISBN 978-80-86668-38-3. [Google Scholar]
- Carrias, J.-F.; Amblard, C.; Bourdier, G.; Sime-Ngando, T. The Importance of Phagotrophic Protists in Lake Pavin. In Lake Pavin; Sime-Ngando, T., Boivin, P., Chapron, E., Jezequel, D., Meybeck, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 307–314. ISBN 978-3-319-39960-7. [Google Scholar]
- Straškraba, M.; Hrbáček, J.; Javornický, P. Effect of an Upstream Reservoir on the Stratification Conditions in Slapy Reservoir. In Hydrobiological Studies 2; Hrbáček, J., Straškraba, M., Eds.; Academia: Prague, Czech Republic, 1973; pp. 7–82. [Google Scholar]
- Kopáček, J.; Hejzlar, J.; Porcal, P.; Posch, M. Trends in Riverine Element Fluxes: A Chronicle of Regional Socio-Economic Changes. Water Res. 2017, 125, 374–383. [Google Scholar] [CrossRef]
- Kopáček, J.; Hejzlar, J.; Porcal, P.; Znachor, P. Biogeochemical Causes of Sixty-Year Trends and Seasonal Variations of River Water Properties in a Large European Basin. Biogeochemistry 2021, 154, 81–98. [Google Scholar] [CrossRef]
- Fott, J.; Nedbalová, L.; Brabec, M.; Kozáková, R.; Řeháková, K.; Hejzlar, J.; Šorf, M.; Vrba, J. Light as a Controlling Factor of Winter Phytoplankton in a Monomictic Reservoir. Limnologica 2022, 95, 125995. [Google Scholar] [CrossRef]
- Lewis, W.M., Jr. A Revised Classification of Lakes Based on Mixing. Can. J. Fish. Aquat. Sci. 1983, 40, 1779–1787. [Google Scholar] [CrossRef]
- Vystavna, Y.; Hejzlar, J.; Kopáček, J. Long-Term Trends of Phosphorus Concentrations in an Artificial Lake: Socio-Economic and Climate Drivers. PLoS ONE 2017, 12, e0186917. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.M.; Wells, S.A. CE-QUAL-W2: A Two-Dimensional, Laterally Averaged, Hydrodynamic and Water Quality Model, version 4.0, Department of Civil and Environmental Engineering, Portland State University, Portland, OR, USA. Available online: https://cee.pdx.edu/w2/ (accessed on 1 April 2024).
- Monsen, N.E.; Cloern, J.E.; Lucas, L.V.; Monismith, S.G. A Comment on the Use of Flushing Time, Residence Time, and Age as Transport Time Scales. Limnol. Oceanogr. 2002, 47, 1545–1553. [Google Scholar] [CrossRef]
- Lorenzen, C.J. Determination of Chlorophyll and Pheo-Pigments: Spectrophotometric Equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Javornický, P. Freshwater Rhodomonads (Cryptophyceae). Arch. Hydrobiol. Algol. Stud. Suppl. Vol. 2001, 102, 93–116. [Google Scholar] [CrossRef]
- Javornický, P. Taxonomic Notes on Some Freshwater Planktonic Cryptophyceae Based on Light Microscopy. Hydrobiologia 2003, 502, 271–283. [Google Scholar] [CrossRef]
- Müller, H.; Schlegel, A. Responses of Three Freshwater Planktonic Ciliates with Different Feeding Modes to Cryptophyte and Diatom Prey. Aquat. Microb. Ecol. 1999, 17, 49–60. [Google Scholar] [CrossRef]
- Jerome, C.A.; Montagnes, D.J.S.; Taylor, F.J.R. The Effect of the Quantitative Protargol Stain and Lugol’s and Bouin’s Fixatives On Cell Size: A More Accurate Estimate of Ciliate Species Biomass. J. Eukaryot. Microbiol. 1993, 40, 254–259. [Google Scholar] [CrossRef]
- Foissner, W.; Berger, H. A User-friendly Guide to the Ciliates (Protozoa, Ciliophora) Commonly Used by Hydrobiologists as Bioindicators in Rivers, Lakes, and Waste Waters, with Notes on Their Ecology. Freshw. Biol. 1996, 35, 375–482. [Google Scholar] [CrossRef]
- Foissner, W.; Berger, H.; Schaumburg, J. Identification and Ecology of Limnetic Plankton Ciliates; Informationsberichte; Informationberichte des Bayerischen Landesamtes Institut für Wasserwirtschaft: Munich, Germany, 1999; ISBN 978-3-930253-79-1. [Google Scholar]
- Lynn, D.H. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature; Springer: New York, NY, USA, 2008; ISBN 978-1-4020-8238-2. [Google Scholar]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef]
- Motulsky, H.J. GraphPad Statistics Guide; GraphPad Software, LLC: San Diego, CA, USA, 2016. [Google Scholar]
- Straškrábová, V. Long-Term Development of Reservoir Ecosystems—Changes in Pelagic Food Webs and Their Microbial Component. Limnetica 2005, 24, 9–20. [Google Scholar] [CrossRef]
- Hyndman, R.J.; Fan, Y. Sample Quantiles in Statistical Packages. Am. Stat. 1996, 50, 361–365. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination; Version 5.1x.; Microcomputer Power: Ithaca, NY, USA, 2018. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- Macek, M.; Jezbera, J.; Sed’a, J.; Straškrábová, V.; Šimek, K.; Han, D. Distribution and Growth Rates of Ciliates along a Longitudinal Profile of the Římov Reservoir. In Proceedings of the Res Lim 2002, 4th International Conference on Reservoir Limnology and Water Quality, České Budějovice, Czech Republic, 12–16 August 2002; Hydrobiological Institute Academy of Sciences of the Czech Republic, Faculty of Biological Sciences University of South Bohemia: České Budějovice, Czech Republic, 2002; pp. 196–199. [Google Scholar]
- Jezbera, J.; Nedoma, J.; Šimek, K. Longitudinal Changes in Protistan Bacterivory and Bacterial Production in Two Canyon-Shaped Reservoirs of Different Trophic Status. Hydrobiologia 2003, 504, 115–130. [Google Scholar] [CrossRef]
- Jeppesen, E.; Sørensen, P.B.; Johansson, L.S.; Søndergaard, M.; Lauridsen, T.L.; Nielsen, A.; Mejlhede, P. Recovery of Lakes from Eutrophication: Changes in Nitrogen Retention Capacity and the Role of Nitrogen Legacy in 10 Danish Lakes Studied over 30 Years. Hydrobiologia 2024, 710, 95–107. [Google Scholar] [CrossRef]
- Pfister, G.; Auer, B.; Arndt, H. Pelagic Ciliates (Protozoa, Ciliophora) of Different Brackish and Freshwater Lakes—A Community Analysis at the Species Level. Limnologica 2002, 32, 147–168. [Google Scholar] [CrossRef]
- Krainer, K.-H. Taxonomische Untersuchungen an neuen und wenig be kannten planktischen Ciliaten (Protozoa: Ciliophora) aus Baggerseen in Österreich. Lauterbornia 1995, 21, 39–68. [Google Scholar]
- Frantal, D.; Agatha, S.; Beisser, D.; Boenigk, J.; Darienko, T.; Dirren-Pitsch, G.; Filker, S.; Gruber, M.; Kammerlander, B.; Nachbaur, L.; et al. Molecular Data Reveal a Cryptic Diversity in the Genus Urotricha (Alveolata, Ciliophora, Prostomatida), a Key Player in Freshwater Lakes, With Remarks on Morphology, Food Preferences, and Distribution. Front. Microbiol. 2022, 12, 787290. [Google Scholar] [CrossRef]
- Sonntag, B.; Frantal, D.; Kammerlander, B.; Darienko, T.; Filker, S.; Stoeck, T.; Gruber, M.; Pröschold, T. Widespread Occurrence of Two Planktonic Ciliate Species (Urotricha, Prostomatida) Originating from High Mountain Lakes. Diversity 2022, 14, 362. [Google Scholar] [CrossRef]
- Humby, P.L.; Lynn, D.H. Taxonomic Description of Six Species of Pelagic, Freshwater Ciliates from the Order Choreotrichida Collected from Lake Ontario, Canada. Aquat. Ecosyst. Health Manag. 2020, 23, 45–57. [Google Scholar] [CrossRef]
- Krainer, K.-H. Contributions to the Morphology, Infraciliature and Ecology of the Planktonic Ciliates Strombidium pelagicum n.sp., Pelagostrombidium mirabile (Penard, 1916) n.g., n. comb., and Pelagostrombidium fallax (Zacharias, 1896) n.g., n. comb. (Ciliophora, Oligotrichida). Eur. J. Protistol. 1991, 27, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, J.; Song, W.; Xu, D.; Warren, A.; Yi, Z.; Gao, S. Multi-Gene-Based Phylogenetic Analysis of Oligotrich Ciliates with Emphasis on Two Dominant Groups: Cyrtostrombidiids and Strombidiids (Protozoa, Ciliophora). Mol. Phylogenet. Evol. 2016, 105, 241–250. [Google Scholar] [CrossRef]
- Ganser, M.H.; Santoferrara, L.F.; Agatha, S. Molecular Signature Characters Complement Taxonomic Diagnoses: A Bioinformatic Approach Exemplified by Ciliated Protists (Ciliophora, Oligotrichea). Mol. Phylogenet. Evol. 2022, 170, 107433. [Google Scholar] [CrossRef] [PubMed]
- Agatha, S.; Weißenbacher, B.; Kirschner, M.; Ganser, M.H. Trichite Features Contribute to the Revision of the Genus Strombidium (Alveolata, Ciliophora, Spirotricha). J. Eukaryot. Microbiol. 2024, 71, e13001. [Google Scholar] [CrossRef]
- Krainer, K.; Foissner, W. Revision of the Genus Askenasia Blochmann, 1895, with Proposal of Two New Species, and Description of Rhabdoaskenasia minima n. g., n. sp. (Ciliophora, Cyclotrichida). J. Protozool. 1990, 37, 414–427. [Google Scholar] [CrossRef]
- Macek, M.; Šimek, K.; Pernthaler, J.; Vyhnálek, V.; Psenner, R. Growth Rates of Dominant Planktonic Ciliates in Two Freshwater Bodies of Different Trophic Degree. J. Plankton Res. 1996, 18, 463–481. [Google Scholar] [CrossRef]
- Küppers, G.C.; Lopretto, E.C.; Claps, M.C. Morphological Aspects and Seasonal Changes of Some Planktonic Ciliates (Protozoa) from a Temporary Pond in Buenos Aires Province, Argentina. Pan-Am. J. Aquat. Sci. 2006, 1, 74–90. [Google Scholar]
- Sommer, F.; Sonntag, B.; Rastl, N.; Summerer, M.; Tartarotti, B. Ciliates in Man-Made Mountain Reservoirs. Front. Environ. Sci. 2022, 10, 903095. [Google Scholar] [CrossRef]
- Lischke, B.; Weithoff, G.; Wickham, S.A.; Attermeyer, K.; Grossart, H.-P.; Scharnweber, K.; HIlt, S.; Gaedke, U. Large Biomass of Small Feeders: Ciliates May Dominate Herbivory in Eutrophic Lakes. J. Plankton Res. 2016, 38, 2–15. [Google Scholar] [CrossRef]
- Šimek, K.; Grujčić, V.; Nedoma, J.; Jezberová, J.; Šorf, M.; Matoušů, A.; Pechar, L.; Posch, T.; Bruni, E.P.; Vrba, J. Microbial Food Webs in Hypertrophic Fishponds: Omnivorous Ciliate Taxa Are Major Protistan Bacterivores. Limnol. Oceanogr. 2019, 64, 2295–2309. [Google Scholar] [CrossRef]
- Tirok, K.; Gaedke, U. Spring Weather Determines the Relative Importance of Ciliates, Rotifers and Crustaceans for the Initiation of the Clear-Water Phase in a Large, Deep Lake. J. Plankton Res. 2006, 28, 361–373. [Google Scholar] [CrossRef]
- Cleven, E. Pelagic Ciliates in a Large Mesotrophic Lake: Seasonal Succession and Taxon-Specific Bacterivory in Lake Constance. Int. Rev. Hydrobiol. 2004, 89, 289–304. [Google Scholar] [CrossRef]
- Müller, H.; Schöne, A.; Pinto-Coelho, R.M.; Schweizer, A.; Weisse, T. Seasonal Succession of Ciliates in Lake Constance. Microb. Ecol. 1991, 21, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Posch, T.; Pitsch, G.; Bruni, E.P. Protists: Ciliates. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2022; pp. 639–649. ISBN 978-0-12-822041-2. [Google Scholar]
- Macek, M.; Šimek, K.; Bittl, T. Conspicuous Peak of Oligotrichous Ciliates Following Winter Stratification in a Bog Lake. J. Plankton Res. 2001, 23, 353–363. [Google Scholar] [CrossRef]
- Šimek, K.; Macek, M.; Pernthaler, J.; Straškrabová, V.; Psenner, R. Can Freshwater Planktonic Ciliates Survive on a Diet of Picoplankton? J. Plankton Res. 1996, 18, 597–613. [Google Scholar] [CrossRef]
- Fenchel, T. Protozoan Filter Feeding. Prog. Protistol. 1986, 1, 65–113. [Google Scholar]
- Bernard, C.; Rassoulzadegan, F. Bacteria or Microflagellates as a Major Food Source for Marine Ciliates: Possible Implications for the Microzooplankton. Mar. Ecol. Prog. Ser. 1990, 64, 147–155. [Google Scholar] [CrossRef]
- Weisse, T. Functional Diversity of Aquatic Ciliates. Eur. J. Protistol. 2017, 61, 331–358. [Google Scholar] [CrossRef]
- Lu, X.; Gao, Y.; Weisse, T. Functional Ecology of Two Contrasting Freshwater Ciliated Protists in Relation to Temperature. J. Eukaryot. Microbiol. 2021, 68, e12823. [Google Scholar] [CrossRef] [PubMed]
- Weisse, T.; Jezberová, J.; Moser, M. Picoplankton Feeding by the Ciliate Vorticella similis in Comparison to Other Peritrichs Emphasizes Their Significance in the Water Purification Process. Ecol. Indic. 2021, 121, 106992. [Google Scholar] [CrossRef]
- Kammerlander, B.; Koinig, K.A.; Rott, E.; Sommaruga, R.; Tartarotti, B.; Trattner, F.; Sonntag, B. Ciliate Community Structure and Interactions within the Planktonic Food Web in Two Alpine Lakes of Contrasting Transparency. Freshw. Biol. 2016, 61, 1950–1965. [Google Scholar] [CrossRef]
- Foissner, W.; Wölfl, S. Revision of the Genus Stentor Oken (Protozoa, Ciliophora) and Description of S. araucanus Nov. Spec, from South American Lakes. J. Plankton Res. 1994, 16, 255–289. [Google Scholar] [CrossRef]
- Modenutti, B.E.; Balseiro, E.G.; Moeller, R. Vertical Distribution and Resistance to Ultraviolet Radiation of a Planktonic Ciliate, Stentor araucanus. SIL Proc. 1922-2010 1998, 26, 1636–1640. [Google Scholar] [CrossRef]
- Woelfl, S. The Distribution of Large Mixotrophic Ciliates (Stentor) in Deep North Patagonian Lakes (Chile): First Results. Limnologica 2007, 37, 28–36. [Google Scholar] [CrossRef]
- Pucciarelli, S.; Buonanno, F.; Pellegrini, G.; Pozzi, S.; Ballarini, P.; Miceli, C. Biomonitoring of Lake Garda: Identification of Ciliate Species and Symbiotic Algae Responsible for the “Black-Spot” Bloom during the Summer of 2004. Environ. Res. 2008, 107, 194–200. [Google Scholar] [CrossRef]
- Mašín, M.; Čuperová, Z.; Hojerová, E.; Salka, I.; Grossart, H.; Koblížek, M. Distribution of Aerobic Anoxygenic Phototrophic Bacteria in Glacial Lakes of Northern Europe. Aquat. Microb. Ecol. 2012, 66, 77–86. [Google Scholar] [CrossRef]
- Müller, H. Laboratory Study of the Life Cycle of a Freshwater Strombidiid Ciliate. Aquat. Microb. Ecol. 2002, 29, 189–197. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Johnson, M.D.; de Vargas, C.; Not, F. Acquired Phototrophy in Aquatic Protists. Aquat. Microb. Ecol. 2009, 57, 279–310. [Google Scholar] [CrossRef]
- Schoener, D.; McManus, G. Plastid Retention, Use, and Replacement in a Kleptoplastidic Ciliate. Aquat. Microb. Ecol. 2012, 67, 177–187. [Google Scholar] [CrossRef]
- Müller, H.; Geller, W. Maximum Growth Rates of Aquatic Ciliated Protozoa: The Dependence on Body Size and Temperature Reconsidered. Arch. Hydrobiol. 1993, 126, 315–327. [Google Scholar] [CrossRef]
- Müller, H.; Weisse, T. Laboratory and Field Observations on the Scuticociliate Histiobalantium from the Pelagic Zone of Lake Constance, FRG. J. Plankton Res. 1994, 16, 391–401. [Google Scholar] [CrossRef]
- Chen, W.-L.; Chiang, K.-P.; Tsai, S.-F. Neglect of Presence of Bacteria Leads to Inaccurate Growth Parameters of the Oligotrich Ciliate Strombidium sp. During Grazing Experiments on Nanoflagellates. Front. Mar. Sci. 2020, 7, 569309. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feeding Behaviour | Family | Genus/Species |
|---|---|---|
| Heterotrophic nanoplankton filtering HF Figure S1 | Strobilidiidae Kahl in Doflein and Reichenow 1929 | Rimostrombidium lacustris (Foissner, Skogstad, and Pratt, 1988) |
| Tintinnidiidae Kofoid and Campbell 1929 | Tintinnidium spp. | |
| Codonellidae Kent, 1881 | Codonella cratera Leidy, 1887 | |
| Incertae sedis Tintinnina Kofoid and Campbell 1929 | Tintinnopsis spp. | |
| Mixotrophic nanoplankton filtering MF Figure S1 | Pelagostrombidiidae Agatha 2004 | Pelagostrombidium spp. Limnostrombidium spp. |
| Strobilidiidae Kahl in Doflein and Reichenow 1929 | Rimostrombidium velox (Faurè-Fremiet, 1924) | |
| Picoplankton filtering PF Figure S2 | Halteriidae Claparède and Lachmann 1858 | Halteria grandinella (Müller, 1773) Halteria—mixotrophic Pelagohalteria viridis (Fromentel, 1876)—mixotrophic |
| Strobilidiidae Kahl in Doflein and Reichenow 1929 | Rimostrombidium brachykinetum Krainer, 1995 Rimostrombidium humile (Penard, 1922) Rimostrombidium—mixotrophic | |
| Vorticellidae Ehrenberg 1838 | Vorticella—minute Vorticella—large Vorticella aqua-dulcis-complex Stokes, 1887 Vorticella mayeri Faure-Fremiet, 1920 | |
| Zoothamniidae Sommer, 1951 | Pseudohaplocaulus sp. | |
| Astylozoidae Kahl 1935 | Astylozoon sp. Hastatella sp. | |
| Trichodinidae Claus 1874 | Trichodina cf. diaptomi Basson and Van As, 1991 | |
| Cinetochilidae Perty 1852 | Cinetochilum margaritaceum Perty, 1852 | |
| Cyclidiidae Ehrenberg 1838 | Cyclidium spp. | |
| other minute scuticociliates | ||
| Algae-hunting AH Figure S3 | Balanionidae Small and Lynn 1985 | Balanion planctonicum (Foissner, Oleksiv and Müller, 1990) |
| Urotrichidae Small and Lynn 1985 | Urotricha~10 µm Urotricha globosa Schewiakoff, 1892 Urotricha furcata Schewiakoff, 1892 Urotricha pseudofurcata Krainer 1995 Urotricha pelagica Kahl, 1932 Urotricha castalia Muñoz, Tellez, and Fernandez-Galiano, 1987 | |
| Histiobalantiidae de Puytorac and Corliss in Corliss 1979 | Histiobalantium spp. | |
| Chilodonellidae Deroux 1970 | Phascolodon vorticella Stein, 1859 Trithigmostoma sp. | |
| Raptorial and flagellate-hunting RH Figure S4 | Colepidae Ehrenberg 1838 | Coleps spp. |
| Actinobolinidae Kahl 1930 | Actinobolina spp. | |
| Enchelyidae Ehrenberg 1838 | Enchelys spp. | |
| Trachelophyllidae Kent 1882 | Lagynophrya spp.—heterotrophic/mixotrophic | |
| Didiniidae Poche 1913 | Monodinium spp.—minute | |
| Incertae sedis CON-threeP | Askenasia chlorelligera Krainer and Foissner, 1990 Askenasia volvox (Penard 1922) Askenasia spp.—minute Askenasia spp.—large Rhabdoaskenasia minima Krainer and Foissner, 1990 | |
| Incertae sedis SAL | Mesodinium spp. Litonotus spp.—minute | |
| Large-cell but low-abundance ciliates LCLA | Epistylididae Kahl 1933 | Epistylis spp.—pelagic, colonial |
| Stentoridae Carus 1863 | Stentor sp. | |
| Holophryidae Perty 1852 | Holophrya spp. | |
| Cyclotrichiidae Jankowski 1980 | Cyclotrichium sp. | |
| Lacrymariidae de Fromentel 1876 | Lacrymaria spp. | |
| Dileptidae Jankowski 1980 | Pelagodileptus spp. | |
| Didiniidae Poche 1913 | Monodinium spp.—large mixotrophic | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macek, M.; Vrba, J.; Hejzlar, J.; Řeháková, K.; Jarošík, J.; Šorf, M.; Šimek, K. Two-Decade Changes in the Ciliate Assemblage Feeding Pattern Reflect the Reservoir Nutrient Load. Diversity 2024, 16, 534. https://doi.org/10.3390/d16090534
Macek M, Vrba J, Hejzlar J, Řeháková K, Jarošík J, Šorf M, Šimek K. Two-Decade Changes in the Ciliate Assemblage Feeding Pattern Reflect the Reservoir Nutrient Load. Diversity. 2024; 16(9):534. https://doi.org/10.3390/d16090534
Chicago/Turabian StyleMacek, Miroslav, Jaroslav Vrba, Josef Hejzlar, Klára Řeháková, Jiří Jarošík, Michal Šorf, and Karel Šimek. 2024. "Two-Decade Changes in the Ciliate Assemblage Feeding Pattern Reflect the Reservoir Nutrient Load" Diversity 16, no. 9: 534. https://doi.org/10.3390/d16090534