1. Introduction
Grass shrimp in the genus
Palaemon occur worldwide in wetlands ranging from freshwater to hypersaline. Several species are present on the east coast of North America, the northern Gulf of Mexico (nGoM), and within the nearby rivers [
1].
Palaemon species play important roles in wetland ecology, contributing to the decomposition of organic material and passing energy to higher trophic levels [
1]. Their annual productivity ranges from 9–16 g dry weight/m
2/year, large enough for them to be ecologically significant [
1]. The daggerblade shrimp,
Palaemon pugio, can reduce the blockages of dead plant matter that form in wetland habitats, maintaining the ecosystem itself [
2].
The family Palaemonidae was previously divided into several subfamilies, with
Palaemon classified in the subfamily Palaemoninae until De Grave et al. [
3] found evidence for paraphyly in the subfamilies and synonymized all of them with Palaemonidae. Similarly,
Palaemon and
Palaemononetes were long considered distinct genera, separated only by the lack of a mandibular palp in
Palaemonetes [
4], but, due to the inconsistency of this character,
Palaemonetes is now considered a junior synonym of
Palaemon [
5]. On the other hand, more recent analysis of
Palaemon detected polyphyly within the genus [
6]. The taxonomy of
Palaemon retains some ambiguities despite extensive research on the subject. Currently,
Palaemon Weber, 1795 contains 95 recognized species [
7].
One ambiguity within
Palaemon exists among
P. pugio (Holthuis, 1949),
P. vulgaris Say, 1818, and
P. mundusnovus De Grave and Ashelby, 2013 (the latter formerly known as
Palaemonetes intermedius). These species have a sympatric distribution in the brackish marshes along the east and Gulf of Mexico coasts of the United States of America [
8]. They are morphologically similar but also display some intraspecific variability [
9]. This analysis includes a fourth species,
Palaemon schmitti (Holthuis, 1950), which has not previously been reported from the nGoM. It is more commonly known from the Tropical Eastern Pacific [
10,
11,
12], the Southern Caribbean, and South America [
13,
14,
15].
Regarding the morphology, we focused on the rostral formula, which is often used for species identification and describes the arrangement of the teeth along the rostrum (
Figure 1). The rostral formula is used here in accordance with [
16] in the following format:
where
a indicates the number of dorsal postorbital teeth on the rostrum,
b the number of preorbital teeth, and
c the number of ventral teeth.
Figure 1.
An example of shrimp rostral formula determination shown on
Palaemon paludosus. For this specimen, the rostral formula would be written as 1)5/3. Images adapted from
https://www.marylandbiodiversity.com/view/19699 (accessed on 1 August 2023).
Figure 1.
An example of shrimp rostral formula determination shown on
Palaemon paludosus. For this specimen, the rostral formula would be written as 1)5/3. Images adapted from
https://www.marylandbiodiversity.com/view/19699 (accessed on 1 August 2023).
Of the species mentioned above,
P. pugio is the most widely studied. It is considered an r-selected species; i.e., it has fast growth, early sexual maturity, frequent reproduction, and high fecundity [
8,
17,
18]. In the coastal wetlands of the southeast United States, it has been referred to as a “core species” [
8].
P. vulgaris generally has smaller population sizes and is considered a “fringe species” in the region [
8]. Further north on the US east coast where both species are common,
P. pugio has a longer reproductive period with three or more spawning cycles per year compared to
P. vulgaris with only two spawning cycles [
19]. Additionally,
P. pugio’s eggs are larger, potentially increasing the offspring’s fitness [
19]. In both species, fecundity decreases with latitude, but the trend is more pronounced in
P. vulgaris [
19]. Comparatively less is known about the life cycle of
P. mundusnovus, but the reproductive period is shorter than in both
P. pugio and
P. vulgaris, and the fecundity is lower than in
P. pugio [
20]. Although this study focuses on the relationships among saltwater
Palaemon species, we also included representatives of several freshwater species (
Table S1) to examine the relationships among the freshwater and saltwater species on the coast of the nGoM.
To provide additional clarity to the relationships within the Palaemon genus, specifically P. pugio, P. vulgaris, and P. mundusnovus, we conducted an analysis of fragments of the mitochondrial cytochrome c oxidase subunit I (COI) and nuclear Histone H3 (H3) genes on specimens from across the northern Gulf of Mexico (GoM) and the US east coast. As the most common “DNA barcoding” sequence for animals, COI is commonly used as a tool for species identification, and a large number of reference sequences from multiple Palaemon species were already publicly available. However, the marker is too variable to provide a solid resolution at deeper taxonomic levels. We therefore supplemented the COI dataset with a more conserved nuclear marker (H3), for which a limited number of reference sequences were also publicly available.
In the GoM, the Mississippi River discharges a massive amount of freshwater and sediment [
21], which may act as a dispersal barrier for coastal species, including grass shrimp. At least 15 marine taxa within the GoM are separated into pairs of genetically distinct species, subspecies, or populations that converge in a vicariance zone near the Mississippi River [
22]. Thus, we hypothesize that the
Palaemon populations on the west and east sides of the Mississippi River would be genetically distinct from each other.
2. Materials and Methods
Live shrimp were collected with dip nets from tidal streams and marshes along the Texas, Florida, and Louisiana coasts. Most of the specimens were found near hard substrates or shaded areas. These collected samples were combined with preserved samples from the Texas A&M Biodiversity Research and Teaching Collections. Shrimp species were identified using dichotomous keys in [
4]. Sampling locations are shown in
Figure 2 (see also
Table S1).
Shrimp were preserved in 95% ethanol, then had their rostral formulas recorded. Abdominal tissue was extracted from each selected shrimp sample for DNA extraction, using the Qiagen Dneasy Blood & Tissue kit, following the manufacturer’s instructions. PCR was then performed to amplify the target gene using the OneTaq kit (New England Biolabs, Ipswich, MA, USA) in 25 microliter (µL) reactions. Each reaction contained 12.5 µL of the OneTaq Master Mix, 1 µL each of the forward and reverse primers at 10 μM, 1 µL of template DNA, and 9.5 µL of water. Multiple COI sequences were collected from each sampling location to define major clades within
Palaemon. Singular Histone H3 sequences were sequenced from each of the sampling locations COI sequences were collected from to resolve deeper branches and increase branch support. COI sequences used the primers LCO1490 and HCO2198 [
23], while Histone H3 sequences used the primers H3aF and H3aR [
24]. The PCR cycle used for COI consisted of (i) 94 °C for 30 s, (ii) denaturation at 94 °C for 30 s, (iii) annealing at 48 °C for 30 s, (iv) extension at 68 °C for 1 min, (v) a total of 30 cycles of steps ii through iv, and (vi) a final extension at 68 °C for 5 min ending with a hold at 4 °C. The PCR cycle for H3 consisted of (i) 94 °C for 7 min, (ii) denaturation at 94 °C for 40 s, (iii) annealing at 57 °C for 40 s, (iv) extension at 72 °C for 1 min, (v) a total of 45 cycles of steps ii through iv, and (vi) a final extension at 72 °C for 7 min ending with a hold at 4 °C.
Successful amplification was confirmed via gel electrophoresis in a 1% agarose gel with ethidium bromide. PCR products were enzymatically cleaned with EXOSAP-IT (ThermoFisher Scientific, Waltham, MA, USA) according to the product’s listed protocol of 15 min at 37 °C followed by 15 min at 80 °C. Product concentration was checked using the Qubit DNA HS protocol, with either 1 or 2 µL of PCR product. Sanger sequencing was performed in both directions at Genewiz/Azenta using the same primers as for the PCR reactions.
Forward and reverse sequences were assembled, followed by trimming of primer sequences in CodonCode Aligner (CodonCode Corporation, Centerville, MA, USA). Resulting consensus sequences were aligned with publicly available
Palaemon sequences in MEGA11 [
25] using the MUSCLE Multiple Sequences Alignment (MSA) program. Four different sequence sets were analyzed: (1) COI sequences from North and Central America only (COI-NCA tree: 124 sequences, length: 585 bp); (2) H3 sequences from North and Central America (H3-NCA tree: 38 sequences, length 286 bp; (3) concatenated COI and H3 sequences from North America (COI + H3 NA tree: 47 sequences, length: 286 bp H3 + 591 bp COI = 877 bp). Only a single H3 sequence was generated for each location in which
Palaemon specimens were collected; all the COI sequences from that location were then combined with the same H3 sequence; and (4) all available COI sequences from global locations (global tree: 231 sequences, length: 539 bp) (
Table S2). Average genetic distances were calculated within and among the North American freshwater and saltwater clades, from in MEGA 11 [
25], using the Kimura-2-Parameter (K2P) model. We chose the K2P model because it is the most commonly used to calculate genetic distances for marine invertebrates, making our values comparable to other studies.
After running Modeltest on the alignments to determine the most appropriate model of evolution, Maximum Likelihood (ML) trees were formed for the multiple sequence alignment (MSA) using 500 bootstrap replicates for the NCA tree and 1000 bootstraps for the global tree. Any duplicate sequences were automatically removed during the tree formation. If the optimal evolution model suggested by Modeltest was available in RaxMLGUI, it was used; if not, the GTR substitution model with Gamma substitution rates with an ML estimate of invariant sites was employed instead. Macrobrachium nipponense was used as an outgroup for rooting purposes for both COI trees. For the COI + H3 tree, the outgroup was Macrobrachium nipponense.
A haplotype network was generated with only the sequences from NCA saltwater species using the minimum spanning network method in PopArt [
26] with an epsilon value of zero. More than 5% of the sites in this analysis contained undefined states and were automatically masked by the PopArt v1.7 for Windows software.
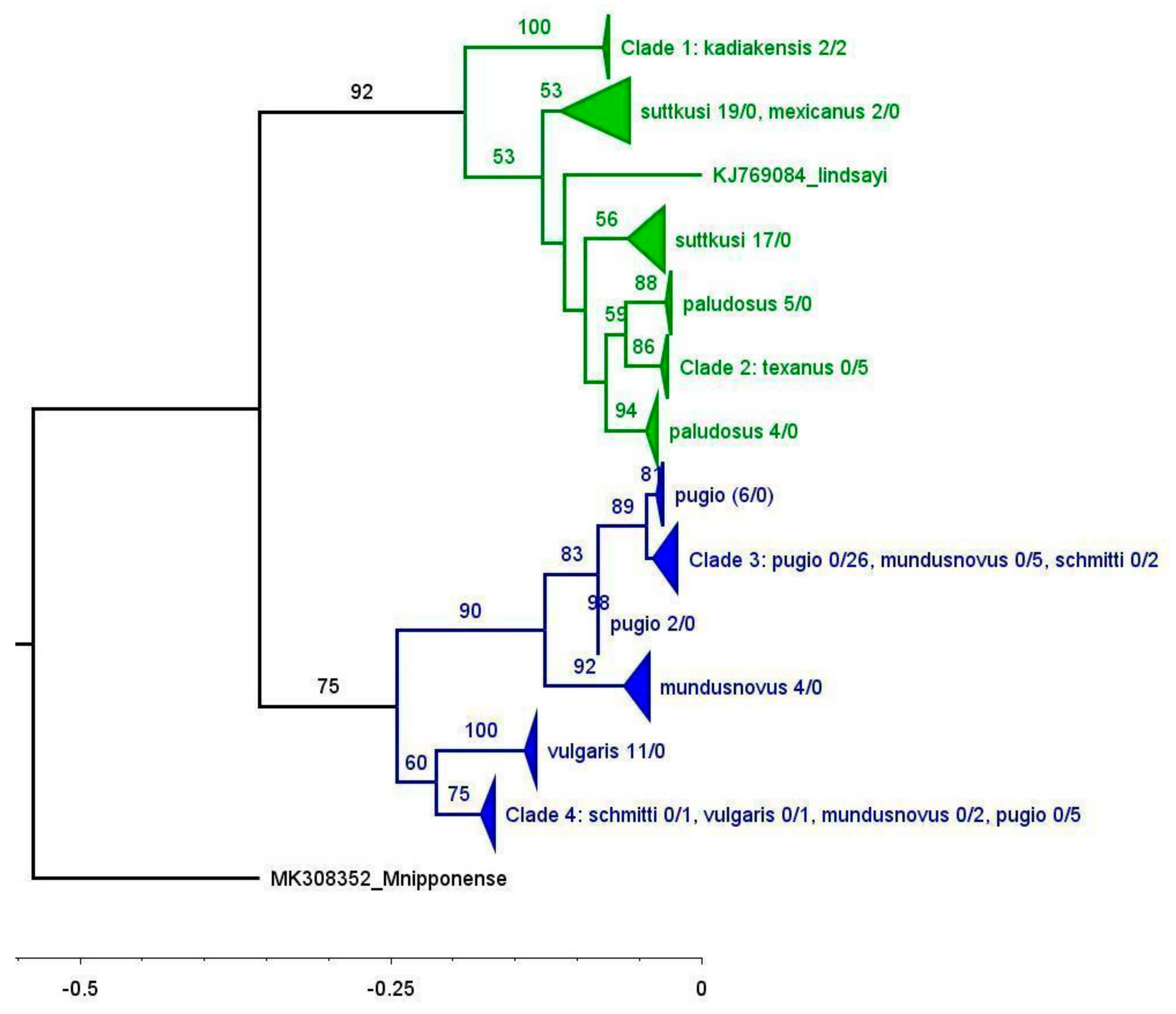
4. Discussion
Our data reveal that the taxonomy of the
Palaemon species from the northern Gulf of Mexico remains poorly resolved, at least when using the taxonomic key in [
4]. As shown in the NCA phylogenetic tree (
Figure 3) and the haplotype network (
Figure 4A), none of the identified saltwater morphospecies from the nGoM form cohesive units.
P. vulgaris from the US east coast forms a well-supported clade, but the single
P. vulgaris from the nGoM shares its haplotype with several
P. pugio individuals (
Figure 3 and
Figure 4A;
Table S1). Similarly, all the
P. mundusnovus samples from the east coast form a monophyletic group (
Figure 3,
Figure 4C and
Figure 5), whereas the nGoM samples of
P. mundusnovus group more closely with
P. vulgaris or
P. pugio.According to [
4],
P. vulgaris and
P. schmitti have two postorbital teeth, whereas only one postorbital tooth is present in
P. pugio and
P. mundusnovus. However, the rostral formula can be highly variable [
9]. Given that Clade 4 appears as the sister group to the east coast
P. vulgaris (
Figure 3) and all its members have two postorbital teeth, it is possible that all the specimens in Clade 4 should actually be classified as
P. vulgaris. Likewise, as Clade 3 is nested within a group of
P. pugio from the east coast, all its members might be correctly classified as
P. pugio. Clade 3 includes two specimens originally identified as
P. schmitti, both of them with two postorbital teeth, but this is possibly a misidentification. The two postorbital teeth in the specimens identified as
P. schmitti may be a further indication for the intraspecific variability of the rostral formula. The incorrect identification is further supported by the fact that the species has not previously been reported from the nGoM, and its type locality is on the Pacific side of the Panama Canal, whereas the other three species (
P. pugio,
P. vulgaris, and
P. mundusnovus) have type localities on the US east coast [
4]. One possible reason why the species identification is challenging is hybridization among the species, a phenomenon that has been documented in other
Palaemon lineages based on mitochondrial/nuclear genomic and karyotype analyses [
27,
28].
The nGoM saltwater samples show no discernible differentiation between the coasts east and west of the Mississippi River Delta (
Figure 4C), indicating high population connectivity along the entire nGoM. Previous studies of
P. pugio found a lack of genetic breaks along the coasts of Georgia [
25] and Texas [
17], but ours is the first to examine the genetic differentiation across the Mississippi River Delta. This finding is remarkable considering the preference of
P. pugio for estuarine habitats, which are more likely to be fragmented than open water, and the fact that
P. pugio reproduces by direct sperm transfer on a seasonal basis rather than broadcast spawning [
26]. It is likely that the population connectivity of
P. pugio is maintained by their larvae, which can tolerate slightly higher salinity levels and have the potential for greater distribution [
18]. Another possible explanation for the high population connectivity is anthropogenic transport through shipping traffic, for example in fouling communities of ships and barges, via the Intracoastal Waterway, which extends along the entire nGoM coast. Assessing the impact of shipping on the distribution of
Palaemon species would require the sequencing of
Palaemon specimens from a vessel’s fouling communities and their putative source and sink populations, as well as knowledge of the vessel’s travel and cleaning schedule.
Palaemon shrimp might be one of many taxa being transported across presumed dispersal barriers in this way. If anthropogenic transport is determined to be a significant factor for the transport of
Palaemon species, assessments of their ecological roles (e.g., interspecific competition and predator-prey interactions) in multiple locations along their range would be warranted.
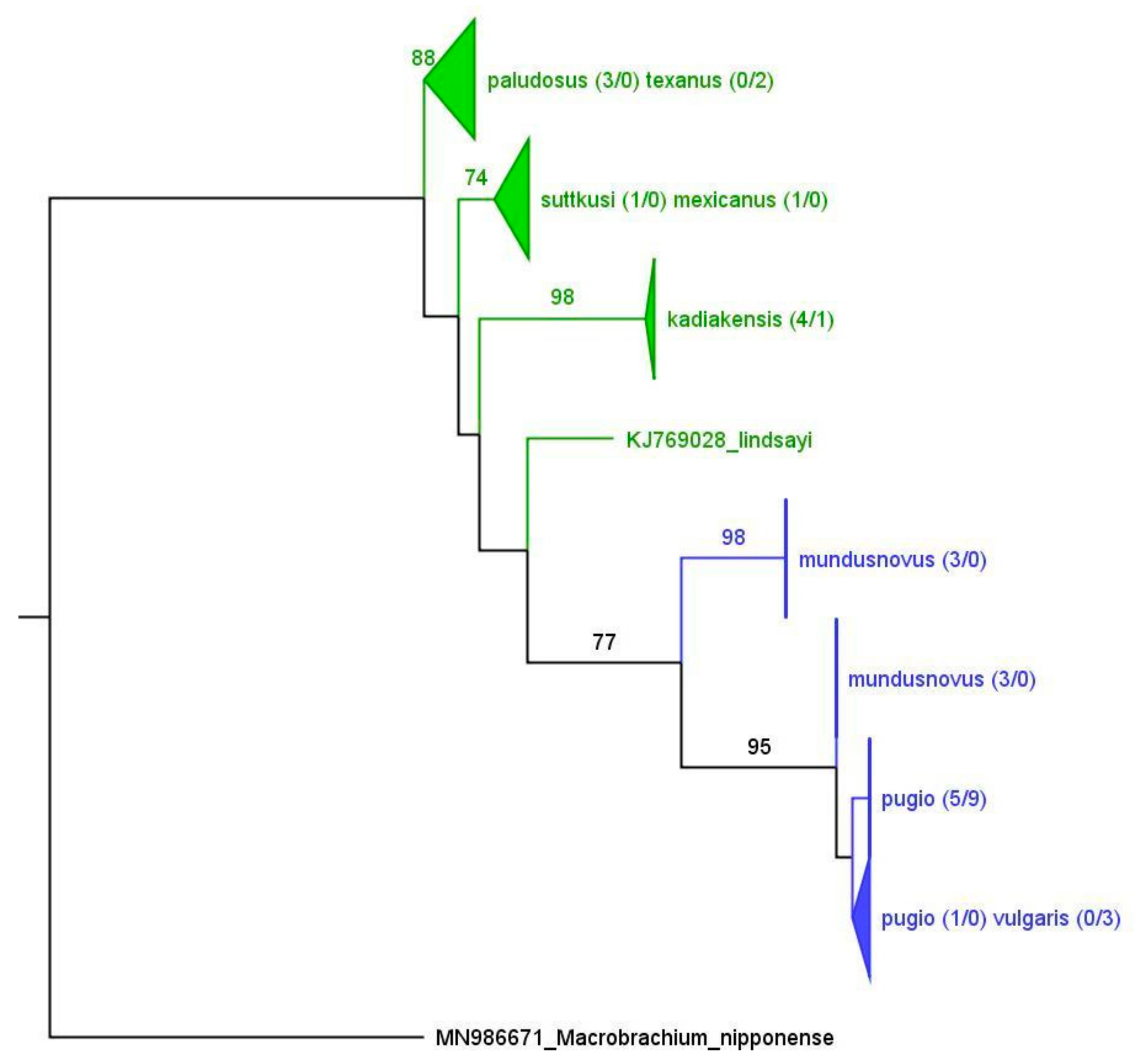
The separation of the saltwater and freshwater
Palaemon species of the nGoM is another interesting finding (
Figure 3,
Figure 5,
Figure 6 and
Figure 7). In fact, the North/Central American freshwater species do not appear to be derived from the saltwater species but rather have colonized the freshwater habitats independently. Our COI datasets revealed average genetic distances of 22.8% between the North/Central American freshwater and saltwater species, supporting their divergence at a deep level within the genus. Based on a wide range of DNA barcoding data for decapods, this value falls well within the range of intrageneric divergence in Palaemonidae [
29]. The relationships among the freshwater species are currently under investigation, with an emphasis on
P. texanus and
P. paludosus.
5. Conclusions
The results of this study demonstrated that a commonly used morphological feature, the rostral formula, is of limited value for the identification of Palaemon species in the nGoM. Given the taxonomic uncertainties within Palaemon, and the history of frequent rearrangements of the genus and the Palaemonidae as a whole, we suggest a shift in focus from the taxonomy to the broader patterns of evolution, population connectivity, morphological, and perhaps physiological and developmental plasticity. Even if careful morphological work may reveal unambiguous species-specific characters, it is unlikely that these will be practical for use without significant investments of time and effort in advanced microscopic methods. Taxonomic names are still the basic units for biodiversity assessments but should be supplemented with additional information about the population characteristics whenever possible.
Our analyses further revealed high population connectivity among the Palaemon populations along the entire nGoM coast, even across the Mississippi River Delta. Whether this high connectivity results from the life history characteristics of grass shrimps or from anthropogenic transport remains to be determined.
Lastly, our data indicate that the saltwater and freshwater species have radiated separately in the region, suggesting that the habitat preferences of Palaemon species are more strongly determined by physiological limits rather than geographic vicinity, even though transitions between saltwater and freshwater have occurred repeatedly within the evolution of the genus. Further research into the environmental tolerances of the different life history stages would lend additional support to this hypothesis.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}