
A 50-Year Perspective on Changes in a Pacific Northwest Breeding Forest Bird Community Reveals General Stability of Abundances



Abstract
:1. Introduction
2. Materials and Methods
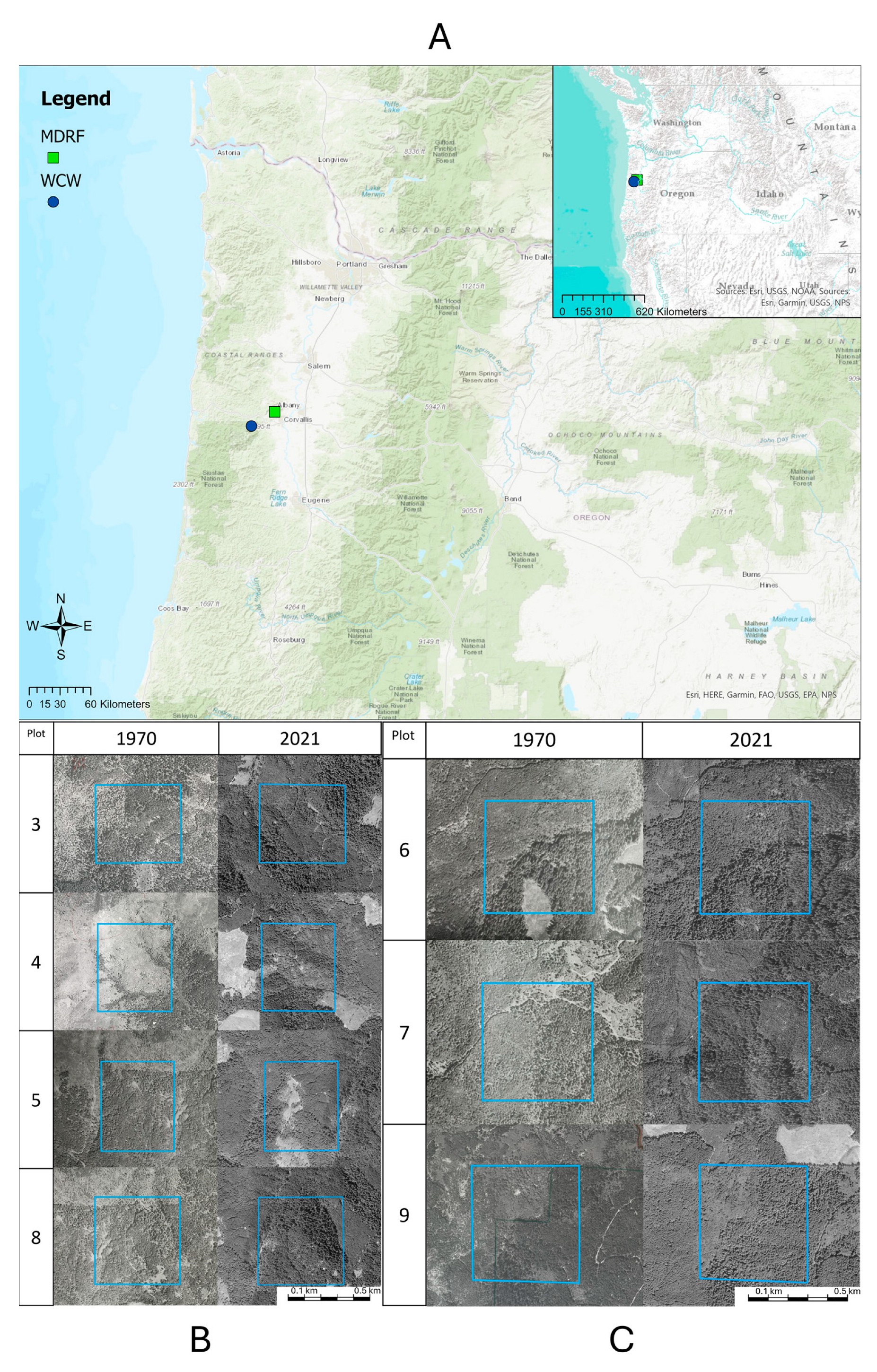
2.1. Study Site and Historical Surveys
2.2. Modern Study Site and Surveys
2.3. Landscape and Habitat Comparison
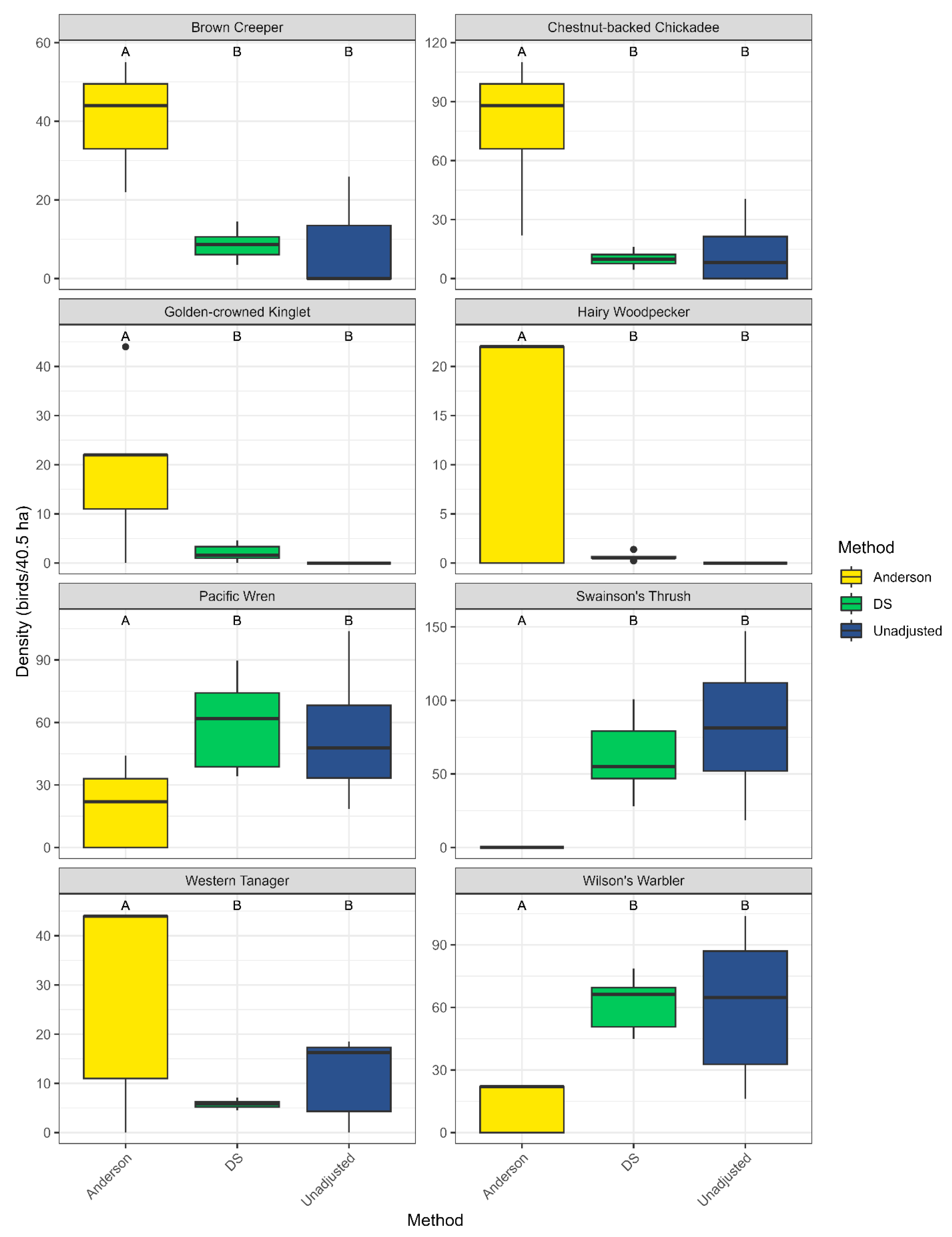
2.4. Density Estimation and Comparison
3. Results
3.1. Avian Communities
3.2. Comparisons with BBS Data
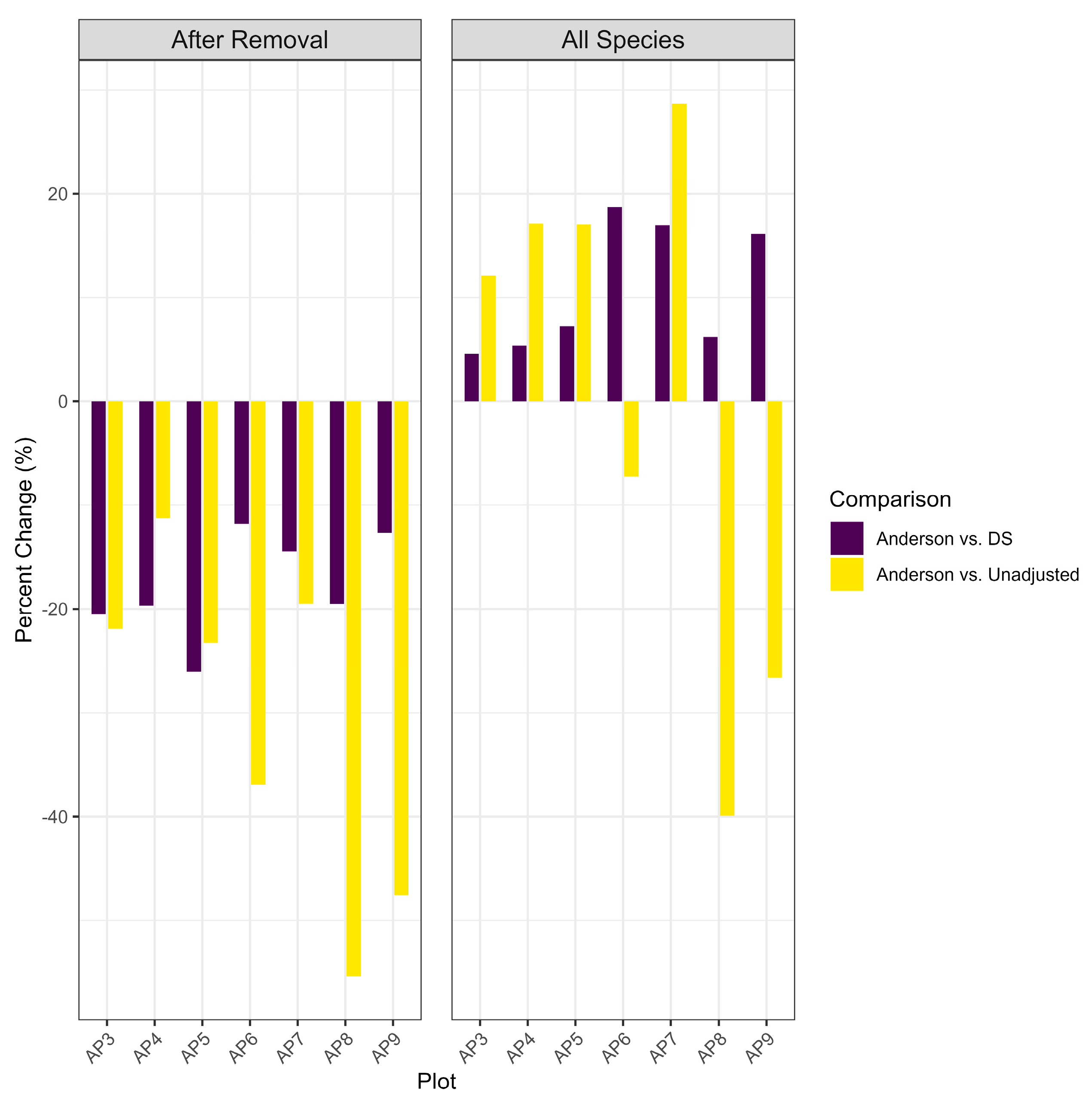
3.3. Habitat Change at Plot and Landscape Levels
3.4. Correlations Between Changes in Habitat and Bird Density
4. Discussion
4.1. Local and Regional Change in Avian Communities
4.2. Avian Response to Agricultural Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BBS | USGS Breeding Bird Survey |
| MDRF | McDonald-Dunn Research Forest |
| WCW | Woods Creek Watershed |
References
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, Models, and Climate Change: Assessing the Assumptions and Uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef]
- Temple, S.; Wiens, J. Bird Populations and Environmental Changes: Can Birds Be Bio-Indicators? Am. Birds 1989, 43, 260–270. [Google Scholar]
- Crick, H.Q.P. The Impact of Climate Change on Birds. Ibis 2004, 146, 48–56. [Google Scholar] [CrossRef]
- Phalan, B.T.; Northrup, J.M.; Yang, Z.; Deal, R.L.; Rousseau, J.S.; Spies, T.A.; Betts, M.G. Impacts of the Northwest Forest Plan on Forest Composition and Bird Populations. Proc. Natl. Acad. Sci. USA 2019, 116, 3322–3327. [Google Scholar] [CrossRef]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American Avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Swanson, F.J.; Franklin, J.F. New Forestry Principles from Ecosystem Analysis of Pacific Northwest Forests. Ecol. Appl. 1992, 2, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.S.H.; Spies, T.A. Forest Cover Changes in the Oregon Coast Range from 1939 to 1993. For. Ecol. Manag. 2004, 200, 129–147. [Google Scholar] [CrossRef]
- McGarigal, K.; McComb, W.C. Relationships Between Landscape Structure and Breeding Birds in the Oregon Coast Range. Ecol. Monogr. 1995, 65, 235–260. [Google Scholar] [CrossRef]
- Betts, M.G.; Hagar, J.C.; Rivers, J.W.; Alexander, J.D.; McGarigal, K.; McComb, B.C. Thresholds in Forest Bird Occurrence as a Function of the Amount of Early-Seral Broadleaf Forest at Landscape Scales. Ecol. Appl. 2010, 20, 2116–2130. [Google Scholar] [CrossRef]
- Harris, S.H.; Betts, M.G. Bird Abundance Is Highly Dynamic across Succession in Early Seral Tree Plantations. For. Ecol. Manag. 2021, 483, 118902. [Google Scholar] [CrossRef]
- Ellis, T.M.; Betts, M.G. Bird Abundance and Diversity across a Hardwood Gradient within Early Seral Plantation Forest. For. Ecol. Manag. 2011, 261, 1372–1381. [Google Scholar] [CrossRef]
- Cahall, R.E.; Hayes, J.P.; Betts, M.G. Will They Come? Long-Term Response by Forest Birds to Experimental Thinning Supports the “Field of Dreams” Hypothesis. For. Ecol. Manag. 2013, 304, 137–149. [Google Scholar] [CrossRef]
- Rivers, J.W.; Verschuyl, J.; Schwarz, C.J.; Kroll, A.J.; Betts, M.G. No Evidence of a Demographic Response to Experimental Herbicide Treatments by the White-Crowned Sparrow, an Early Successional Forest Songbird. Condor 2019, 121, duz004. [Google Scholar] [CrossRef]
- Sallabanks, R.; Arnett, E.B.; Marzluff, J.M. An Evaluation of Research on the Effects of Timber Harvest on Bird Populations. Wildl. Soc. Bull. 2000, 28, 1144–1155. [Google Scholar]
- Vanderwel, M.C.; Malcolm, J.R.; Mills, S.C. A Meta-Analysis of Bird Responses to Uniform Partial Harvesting across North America. Conserv. Biol. 2007, 21, 1230–1240. [Google Scholar] [CrossRef]
- Link, W.A.; Sauer, J.R. Estimating Population Change from Count Data: Application to the North American Breeding Bird Survey. Ecol. Appl. 1998, 8, 258–268. [Google Scholar] [CrossRef]
- Magurran, A.E.; Baillie, S.R.; Buckland, S.T.; Dick, J.M.; Elston, D.A.; Scott, E.M.; Smith, R.I.; Somerfield, P.J.; Watt, A.D. Long-Term Datasets in Biodiversity Research and Monitoring: Assessing Change in Ecological Communities through Time. Trends Ecol. Evol. 2010, 25, 574–582. [Google Scholar] [CrossRef]
- Igl, L.D.; Johnson, D.H. A Retrospective Perspective: Evaluating Population Changes by Repeating Historic Bird Surveys. USDA For. Serv. Gen. Tech. Rep. 2005, PSW-GTR-191, 817–830. [Google Scholar]
- Tingley, M.W. Turning Oranges into Apples: Using Detectability Correction and Bias Heuristics to Compare Imperfectly Repeated Observations. In Stepping in the Same River Twice: Replication in Biological Research; Yale University Press: London, UK, 2017; pp. 215–233. [Google Scholar]
- Robinson, W.D. Long-Term Changes in the Avifauna of Barro Colorado Island, Panama, a Tropical Forest Isolate. Conserv. Biol. 1999, 13, 85–97. [Google Scholar] [CrossRef]
- Iknayan, K.J.; Beissinger, S.R. Collapse of a Desert Bird Community over the Past Century Driven by Climate Change. Proc. Natl. Acad. Sci. USA 2018, 115, 8597–8602. [Google Scholar] [CrossRef] [PubMed]
- Clements, N.M.; Robinson, W.D. A Comparison of Two Snapshot Studies Half a Century Apart Suggests Stability in a Pacific Northwest Winter Forest Bird Community. Front. Bird Sci. 2024, 3, 1304026. [Google Scholar] [CrossRef]
- Tingley, M.W.; Beissinger, S.R. Detecting Range Shifts from Historical Species Occurrences: New Perspectives on Old Data. Trends Ecol. Evol. 2009, 24, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Tingley, M.W.; Monahan, W.B.; Beissinger, S.R.; Moritz, C. Birds Track Their Grinnellian Niche through a Century of Climate Change. Proc. Natl. Acad. Sci. USA 2009, 106, 19637–19643. [Google Scholar] [CrossRef] [PubMed]
- Tingley, M.W.; Beissinger, S.R. Cryptic Loss of Montane Avian Richness and High Community Turnover over 100 Years. Ecology 2013, 94, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.R.; Robinson, W.D. Sixty Years of Change in Avian Communities of the Pacific Northwest. PeerJ 2015, 3, e1152. [Google Scholar] [CrossRef]
- Curtis, J.R.; Robinson, W.D.; McCune, B. Time Trumps Habitat in the Dynamics of an Avian Community. Ecosphere 2016, 7, e01575. [Google Scholar] [CrossRef]
- Ellis, M.S.; Kennedy, P.L.; Edge, W.D.; Sanders, T.A. Twenty-Year Changes in Riparian Bird Communities of East-Central Oregon. Wilson J. Ornithol. 2019, 131, 43–61. [Google Scholar] [CrossRef]
- Anderson, S.H. Ecological Relationships of Birds in Forest of Western Oregon; Oregon State University: Corvallis, OR, USA, 1970. [Google Scholar]
- Anderson, S.H. Seasonal Variations in Forest Birds of Western Oregon. Northwest Sci. 1972, 46, 194–206. [Google Scholar]
- Cottam, G.; Curtis, J.T. The Use of Distance Measures in Phytosociological Sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Clements, N.; Robinson, W. A Re-Survey in 2019-2021 of Winter Bird Communities in the Oregon Coast Range, USA, Initially Surveyed in 1968–1970. Biodivers. Data J. 2022, 10, e91511. [Google Scholar] [CrossRef] [PubMed]
- James, F.C.; Shugart, H.H. A Quantitative Method of Habitat Description. Audubon Field Notes 1970, 24, 727–736. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sample: Estimating Abundance of Biological Populations; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Robinson, W.D.; Hallman, T.A.; Curtis, J.R. Benchmarking the Avian Diversity of Oregon in an Era of Rapid Change. Northwestern Nat. 2020, 101, 180–193. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Thomas, L.; Buckland, S.T.; Rexstad, E.; Laake, J.L.; Strindberg, S.; Hedley, S.L.; Bishop, J.R.B.; Marques, T.A. Distance Software: Design and Analysis of Distance Sampling Surveys for Estimating Population Size. J. Appl. Ecol. 2010, 47, 5–14. [Google Scholar] [CrossRef]
- Sauer, J.R.; Link, W.A.; Hines, J.E. The North American Breeding Bird Survey, Analysis Results 1966–2019; Eastern Ecological Science Center at the Leetown Research Laboratory: Kearneysville, WV, USA, 2020. [Google Scholar]
- Chesser, R.T.; Billerman, S.M.; Burns, K.J.; Cicero, C.; Dunn, J.L.; Hernández-Baños, B.E.; Jiménez, R.A.; Johnson, O.; Kratter, A.W.; Mason, N.A.; et al. AOU Checklist of North and Middle American Birds. Available online: https://checklist.americanornithology.org/taxa/ (accessed on 11 September 2024).
- Shen, F.-Y.; Ding, T.-S.; Tsai, J.-S. Comparing Avian Species Richness Estimates from Structured and Semi-Structured Citizen Science Data. Sci. Rep. 2023, 13, 1214. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, G.L.; Pollock, K.H.; Nichols, J.D.; Simons, T.R.; Hines, J.E.; Sauer, J.R. A Removal Model for Estimating Detection Probabilities from Point-Count Surveys. Auk 2002, 119, 414–425. [Google Scholar] [CrossRef]
- Edwards, B.P.M.; Smith, A.C.; Docherty, T.D.S.; Gahbauer, M.A.; Gillespie, C.R.; Grinde, A.R.; Harmer, T.; Iles, D.T.; Matsuoka, S.M.; Michel, N.L.; et al. Point Count Offsets for Estimating Population Sizes of North American Landbirds. Ibis 2023, 165, 482–503. [Google Scholar] [CrossRef]
- Stanton, R.L.; Morrissey, C.A.; Clark, R.G. Analysis of Trends and Agricultural Drivers of Farmland Bird Declines in North America: A Review. Agric. Ecosyst. Environ. 2018, 254, 244–254. [Google Scholar] [CrossRef]
- Lees, A.C.; Haskell, L.; Allinson, T.; Bezeng, S.B.; Burfield, I.J.; Renjifo, L.M.; Rosenberg, K.V.; Viswanathan, A.; Butchart, S.H.M. State of the World’s Birds. Annu. Rev. Environ. Resour. 2022, 47, 231–260. [Google Scholar] [CrossRef]
- Ammon, E.M.; Gilbert, W.M. Wilson’s Warbler (Cardellina Pusilla), Version 1.0. In Birds World; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Dunn, J.L.; Garrett, K.L. A Field Guide to the Warblers of North America; Houghton Mifflin Company: Boston, MA, USA, 1997. [Google Scholar]
- Fuller, R.J. Birds and Habitat: Relationships in Changing Landscapes; Cambridge University Press: New York, NY, USA, 2012. [Google Scholar]
- Schmidt, K.A. Information Thresholds, Habitat Loss and Population Persistence in Breeding Birds. Oikos 2017, 126, 651–659. [Google Scholar] [CrossRef]
- Stralberg, D.; Bayne, E.M.; Cumming, S.G.; Sólymos, P.; Song, S.J.; Schmiegelow, F.K.A. Conservation of Future Boreal Forest Bird Communities Considering Lags in Vegetation Response to Climate Change: A Modified Refugia Approach. Divers. Distrib. 2015, 21, 1112–1128. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape Modification and Habitat Fragmentation: A Synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Cline, S.P.; Berg, A.B.; Wight, H.M. Snag Characteristics and Dynamics in Douglas-Fir Forests, Western Oregon. J. Wildl. Manag. 1980, 44, 773–786. [Google Scholar] [CrossRef]
- Chambers, C.L.; Carrigan, T.; Sabin, T.E.; Tappeiner, J.; McComb, W.C. Use of Artificially Created Douglas-Fir Snags by Cavity-Nesting Birds. West. J. Appl. For. 1997, 12, 93–97. [Google Scholar] [CrossRef]
- Keller, J.K.; Richmond, M.E.; Smith, C.R. An Explanation of Patterns of Breeding Bird Species Richness and Density Following Clearcutting in Northeastern USA Forests. For. Ecol. Manag. 2003, 174, 541–564. [Google Scholar] [CrossRef]
- McWethy, D.B.; Hansen, A.J.; Verschuyl, J.P. Bird Response to Disturbance Varies with Forest Productivity in the Northwestern United States. Landsc. Ecol. 2010, 25, 533–549. [Google Scholar] [CrossRef]
- Robinson, W.D.; Curtis, J.R. Creating Benchmark Measurements of Tropical Forest Bird Communities in Large Plots. Condor 2020, 122, 1–15. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Allen, C.D.; Betancourt, J.L. Applied Historical Ecology: Using the Past to Manage for the Future. Ecol. Appl. 1999, 9, 1189–1206. [Google Scholar] [CrossRef]
- Szabó, P. Historical Ecology: Past, Present and Future. Biol. Rev. 2015, 90, 997–1014. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Anderson | Unadjusted | DS | Anderson | Unadjusted | DS | Anderson | Unadjusted | DS | Anderson | Unadjusted | DS | Anderson | Unadjusted | DS | Anderson | Unadjusted | DS | Anderson | Unadjusted | DS |
| Turkey Vulture (Cathartes aura) | 11 | 0 | 0 | 11 | 0 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Red-tailed Hawk (Buteo jamaicensis) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Ruffed Grouse (Bonasa umbellus) | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 | 0 | 0 | 0 |
| Mountain Quail (Oreortyx pictus) | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 | 0 | 0 | 0 |
| Mourning Dove (Zenaida macroura) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Band-tailed Pigeon (Patagioenas fasciata) | 0 | 24 | 2 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 32 | 2 | 0 | 9 | 1 | 0 | 8 | 1 | 0 | 0 | 2 |
| Rufous Hummingbird (Selasphorus rufus) | 22 | 0 | 0 | 11 | 9 | 42 | 22 | 17 | 116 | 0 | 0 | 0 | 0 | 26 | 119 | 0 | 0 | 37 | 0 | 0 | 0 |
| Vaux’s Swift (Chaetura vauxi) | 0 | 0 | 0 | 0 | 0 | 0 | 110 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Red-breasted Sapsucker (Sphyrapicus ruber) | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 |
| Downy Woodpecker (Dryobates pubescens) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 0 | 22 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 | 22 | 0 | 0 |
| Hairy Woodpecker (Dryobates villosus) | 22 | 0 | 1 | 0 | 0 | 1 | 22 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 1 | 22 | 0 | 1 |
| Northern Flicker (Colaptes auratus) | 0 | 0 | 0 | 11 | 0 | 1 | 0 | 0 | 0 | 0 | 16 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pileated Woodpecker (Dryocopus pileatus) | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 |
| Olive-sided Flycatcher (Contopus cooperi) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Western Wood-Pewee (Contopus sordidulus) | 44 | 0 | 0 | 22 | 0 | 0 | 22 | 0 | 0 | 22 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 | 22 | 0 | 0 |
| Willow Flycatcher (Empidonax traillii) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hammond’s Flycatcher (Empidonax hammondii) | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 17 | 0 | 0 | 5 | 0 | 0 | 0 | 22 | 0 | 7 |
| Dusky Flycatcher (Empidonax oberholseri) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 22 | 0 | 0 |
| Western Flycatcher (Empidonax difficilis) | 0 | 41 | 43 | 22 | 9 | 21 | 22 | 76 | 35 | 66 | 16 | 46 | 22 | 26 | 48 | 22 | 41 | 33 | 22 | 80 | 58 |
| Canada Jay (Perisoreus canadensis) | 0 | 8 | 2 | 0 | 19 | 2 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 5 | 0 | 0 | 3 | 22 | 18 | 6 |
| Steller’s Jay (Cyanocitta stelleri) | 44 | 24 | 7 | 0 | 9 | 8 | 22 | 8 | 6 | 0 | 0 | 5 | 22 | 0 | 3 | 22 | 0 | 4 | 0 | 0 | 2 |
| Common Raven (Corvus corax) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Purple Martin (Progne subis) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Chestnut-backed Chickadee (Poecile rufescens) | 66 | 41 | 16 | 88 | 0 | 4 | 88 | 25 | 13 | 22 | 0 | 8 | 66 | 17 | 10 | 110 | 8 | 12 | 110 | 0 | 8 |
| Red-breasted Nuthatch (Sitta canadensis) | 44 | 41 | 47 | 44 | 28 | 29 | 66 | 25 | 34 | 22 | 16 | 61 | 66 | 17 | 37 | 33 | 41 | 34 | 22 | 0 | 16 |
| Brown Creeper (Certhia americana) | 55 | 0 | 9 | 44 | 19 | 8 | 22 | 8 | 3 | 44 | 0 | 4 | 44 | 26 | 12 | 55 | 0 | 15 | 22 | 0 | 9 |
| House Wren (Troglodytes aedon) | 44 | 0 | 2 | 0 | 9 | 4 | 22 | 17 | 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Pacific Wren (Troglodytes pacificus) | 0 | 57 | 39 | 0 | 19 | 39 | 22 | 42 | 34 | 44 | 48 | 76 | 22 | 104 | 72 | 0 | 24 | 62 | 44 | 80 | 90 |
| Golden-crowned Kinglet (Regulus satrapa) | 22 | 0 | 0 | 44 | 0 | 2 | 22 | 0 | 1 | 22 | 0 | 4 | 0 | 0 | 3 | 0 | 0 | 0 | 22 | 0 | 5 |
| American Robin (Turdus migratorius) | 0 | 24 | 7 | 0 | 9 | 8 | 0 | 8 | 8 | 0 | 0 | 1 | 0 | 0 | 6 | 0 | 0 | 8 | 0 | 0 | 1 |
| Varied Thrush (Ixoreus naevius) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16 | 5 | 0 | 43 | 7 | 0 | 0 | 0 | 0 | 9 | 13 |
| Hermit Thrush (Catharus guttatus) | 0 | 8 | 6 | 0 | 9 | 11 | 11 | 0 | 1 | 77 | 0 | 0 | 22 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| Swainson’s Thrush (Catharus ustulatus) | 0 | 81 | 55 | 0 | 19 | 28 | 0 | 135 | 66 | 0 | 80 | 55 | 0 | 147 | 92 | 0 | 24 | 39 | 0 | 88 | 101 |
| Hutton’s Vireo (Vireo huttoni) | 22 | 0 | 2 | 0 | 0 | 4 | 0 | 0 | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | 22 | 0 | 0 |
| Warbling Vireo (Vireo gilvus) | 0 | 0 | 0 | 0 | 28 | 25 | 0 | 25 | 12 | 0 | 0 | 4 | 0 | 17 | 7 | 0 | 0 | 1 | 0 | 0 | 13 |
| Purple Finch (Haemorhous purpureus) | 0 | 0 | 20 | 0 | 0 | 4 | 0 | 8 | 42 | 0 | 0 | 20 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 |
| Pine Siskin (Spinus pinus) | 0 | 0 | 11 | 0 | 9 | 5 | 0 | 0 | 4 | 0 | 16 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 9 | 4 |
| Red Crossbill (Loxia curvirostra) | 0 | 8 | 97 | 0 | 0 | 18 | 0 | 0 | 85 | 0 | 0 | 32 | 0 | 9 | 86 | 0 | 8 | 49 | 0 | 9 | 159 |
| Evening Grosbeak (Coccothraustes vespertinus) | 0 | 8 | 1 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 48 | 14 | 44 | 9 | 7 | 0 | 0 | 1 | 0 | 9 | 3 |
| American Goldfinch (Spinus tristis) | 0 | 0 | 0 | 0 | 9 | 13 | 0 | 0 | 6 | 0 | 0 | 11 | 0 | 9 | 6 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spotted Towhee (Pipilo maculatus) | 0 | 32 | 40 | 0 | 0 | 10 | 22 | 17 | 29 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16 | 20 | 0 | 0 | 0 |
| Dark-eyed Junco (Junco hyemalis) | 44 | 57 | 51 | 22 | 28 | 34 | 44 | 68 | 53 | 44 | 16 | 7 | 44 | 9 | 18 | 55 | 32 | 52 | 44 | 0 | 5 |
| Song Sparrow (Melospiza melodia) | 0 | 0 | 2 | 0 | 0 | 4 | 0 | 8 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 |
| White-crowned Sparrow (Zonotrichia leucophrys) | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 8 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Orange-crowned Warbler (Leiothlypis celata) | 0 | 49 | 45 | 0 | 46 | 35 | 0 | 59 | 99 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 24 | 19 | 0 | 0 | 0 |
| Yellow Warbler (Setophaga petechia) | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Black-throated Gray Warbler (Setophaga nigrescens) | 0 | 16 | 8 | 0 | 0 | 0 | 0 | 17 | 14 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 4 | 0 | 0 | 0 |
| Hermit Warbler (Setophaga occidentalis) | 44 | 41 | 18 | 0 | 56 | 15 | 0 | 8 | 6 | 0 | 48 | 32 | 44 | 35 | 28 | 0 | 8 | 6 | 0 | 35 | 22 |
| MacGillivray’s Warbler (Geothlypis tolmiei) | 22 | 0 | 3 | 0 | 19 | 5 | 0 | 34 | 22 | 0 | 0 | 0 | 22 | 0 | 3 | 0 | 0 | 0 | 22 | 0 | 0 |
| Wilson’s Warbler (Cardellina pusilla) | 22 | 89 | 66 | 22 | 65 | 51 | 22 | 85 | 68 | 0 | 48 | 71 | 22 | 104 | 79 | 0 | 16 | 50 | 0 | 18 | 45 |
| Western Tanager (Piranga ludoviciana) | 44 | 16 | 5 | 22 | 19 | 6 | 44 | 17 | 6 | 0 | 0 | 7 | 44 | 9 | 7 | 0 | 0 | 6 | 44 | 18 | 5 |
| Black-headed Grosbeak (Pheucticus melanocephalus) | 22 | 0 | 2 | 0 | 9 | 4 | 0 | 17 | 2 | 0 | 0 | 2 | 0 | 9 | 4 | 0 | 0 | 2 | 0 | 0 | 2 |
| Plot | Anderson | Unadjusted | DS |
|---|---|---|---|
| 3 | 17 | 19 | 36 |
| 4 | 15 | 22 | 41 |
| 5 | 20 | 25 | 41 |
| 6 | 12 | 12 | 30 |
| 7 | 13 | 18 | 32 |
| 8 | 13 | 12 | 36 |
| 9 | 16 | 11 | 28 |
| Total | 34 | 35 | 41 |
| Plot | Δ Shrubs/ha | Δ Trees/ha | Δ Percent Forest Cover | Δ Percent Clearcut Cover | Percent Harvested |
|---|---|---|---|---|---|
| AP3 | −264 | 236 | 7.0 | −0.5 | 28.9 |
| AP4 | −8030 | −101 | 78.8 | −80.4 | 6.5 |
| AP5 | −10860 | 71 | 6.9 | 1.5 | 71.5 |
| AP6 | −2099 | −107 | 6.1 | −6.1 | 0 |
| AP7 | −2337 | 213 | 8.3 | −3.8 | 21.02 |
| AP8 | −16052 | 201 | 12.5 | −8.3 | 11.5 |
| AP9 | −9367 | −49 | 0.8 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clements, N.M.; Shen, F.-Y.; Robinson, W.D. A 50-Year Perspective on Changes in a Pacific Northwest Breeding Forest Bird Community Reveals General Stability of Abundances. Diversity 2025, 17, 123. https://doi.org/10.3390/d17020123
Clements NM, Shen F-Y, Robinson WD. A 50-Year Perspective on Changes in a Pacific Northwest Breeding Forest Bird Community Reveals General Stability of Abundances. Diversity. 2025; 17(2):123. https://doi.org/10.3390/d17020123
Chicago/Turabian StyleClements, Nolan M., Fang-Yu Shen, and W. Douglas Robinson. 2025. "A 50-Year Perspective on Changes in a Pacific Northwest Breeding Forest Bird Community Reveals General Stability of Abundances" Diversity 17, no. 2: 123. https://doi.org/10.3390/d17020123
APA StyleClements, N. M., Shen, F.-Y., & Robinson, W. D. (2025). A 50-Year Perspective on Changes in a Pacific Northwest Breeding Forest Bird Community Reveals General Stability of Abundances. Diversity, 17(2), 123. https://doi.org/10.3390/d17020123