


Genetic Diversity of Calligonum L. Species Analyzed Using Newly Developed Genomic Simple Sequence Repeats and Their Application to Conservation

 ,
,
Abstract
1. Introduction
2. Results
2.1. An Analysis of the Distribution of SSRs in the Genome of Calligonum
2.2. Development of Genome SSR Markers for Calligonum
2.3. Genetic Diversity of Calligonum L.
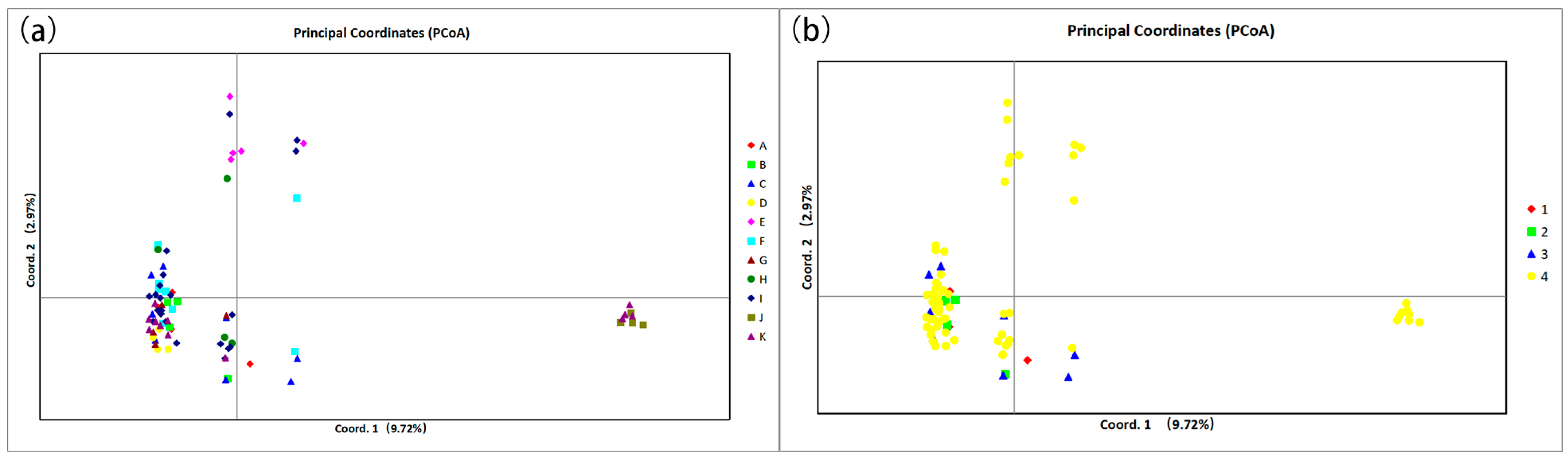
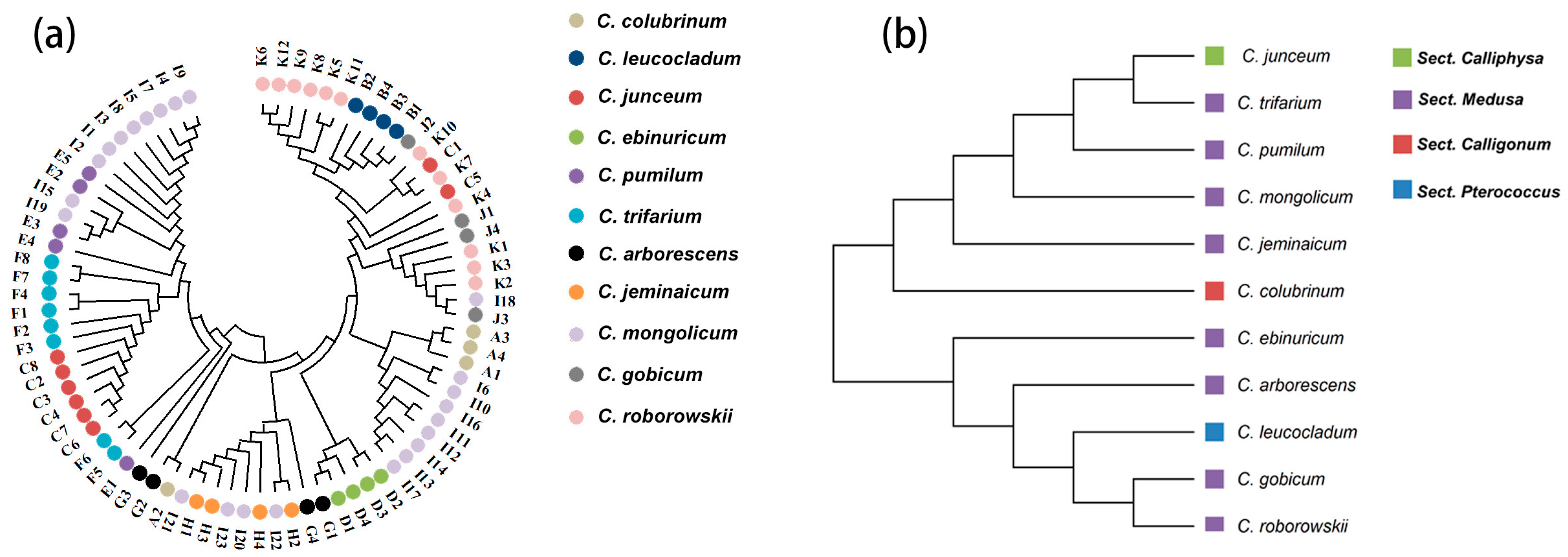
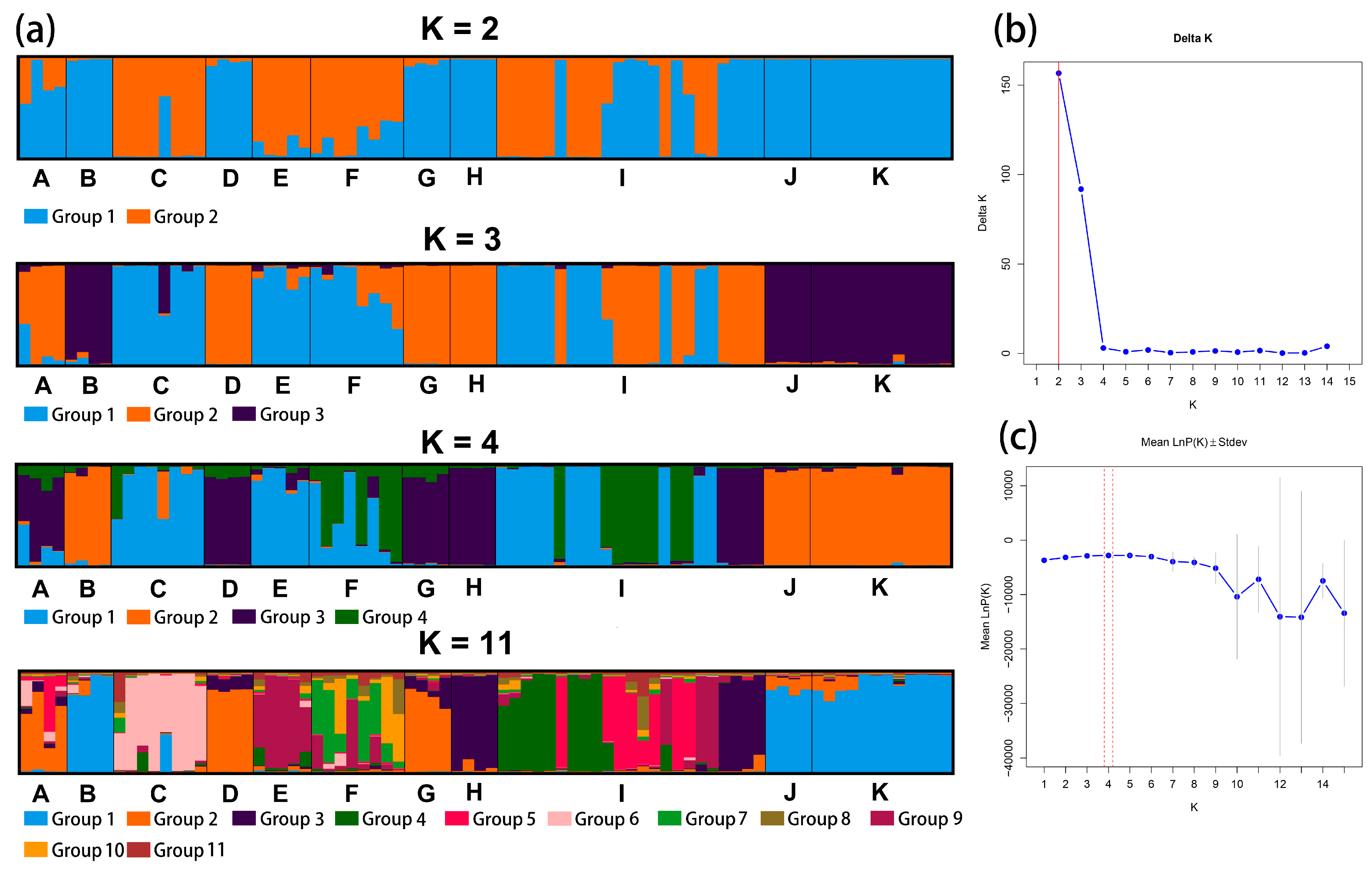
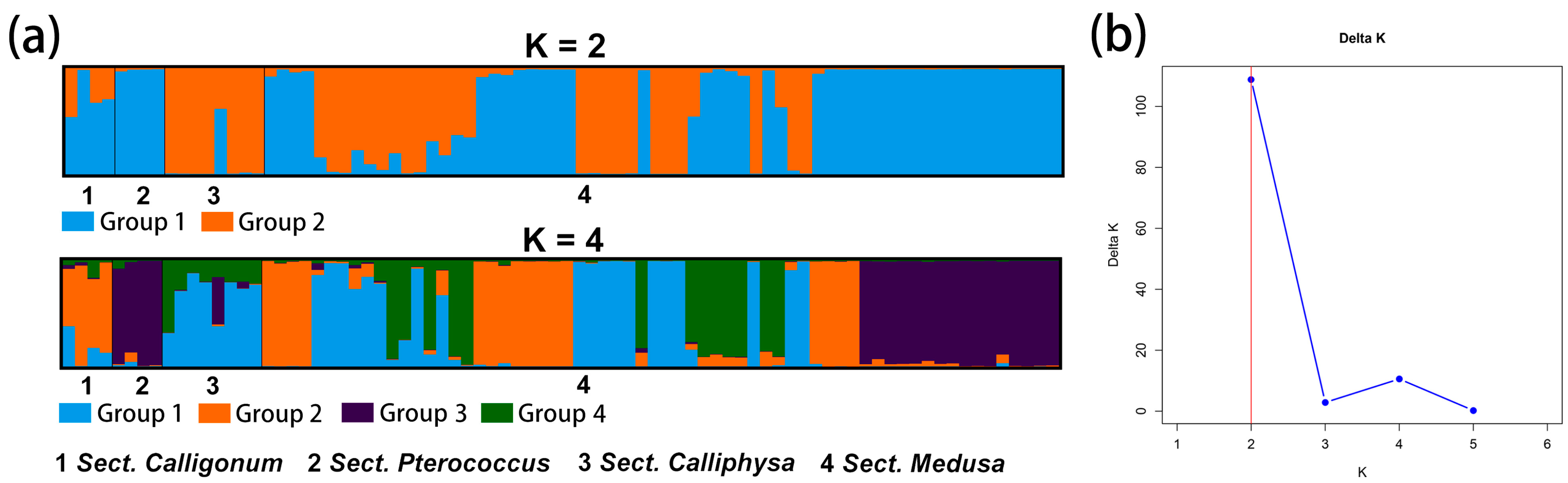
2.4. Genetic Relationship, Structure, and Differentiation Among 11 Species of Calligonum L.
3. Discussion
3.1. Genetic Diversity of Calligonum L.
3.2. Breakdown in Genetic Structure and Taxonomic Identification in Calligonum L.
3.3. Conservation and Utilization of Calligonum L. Genetic Resources
4. Materials and Methods
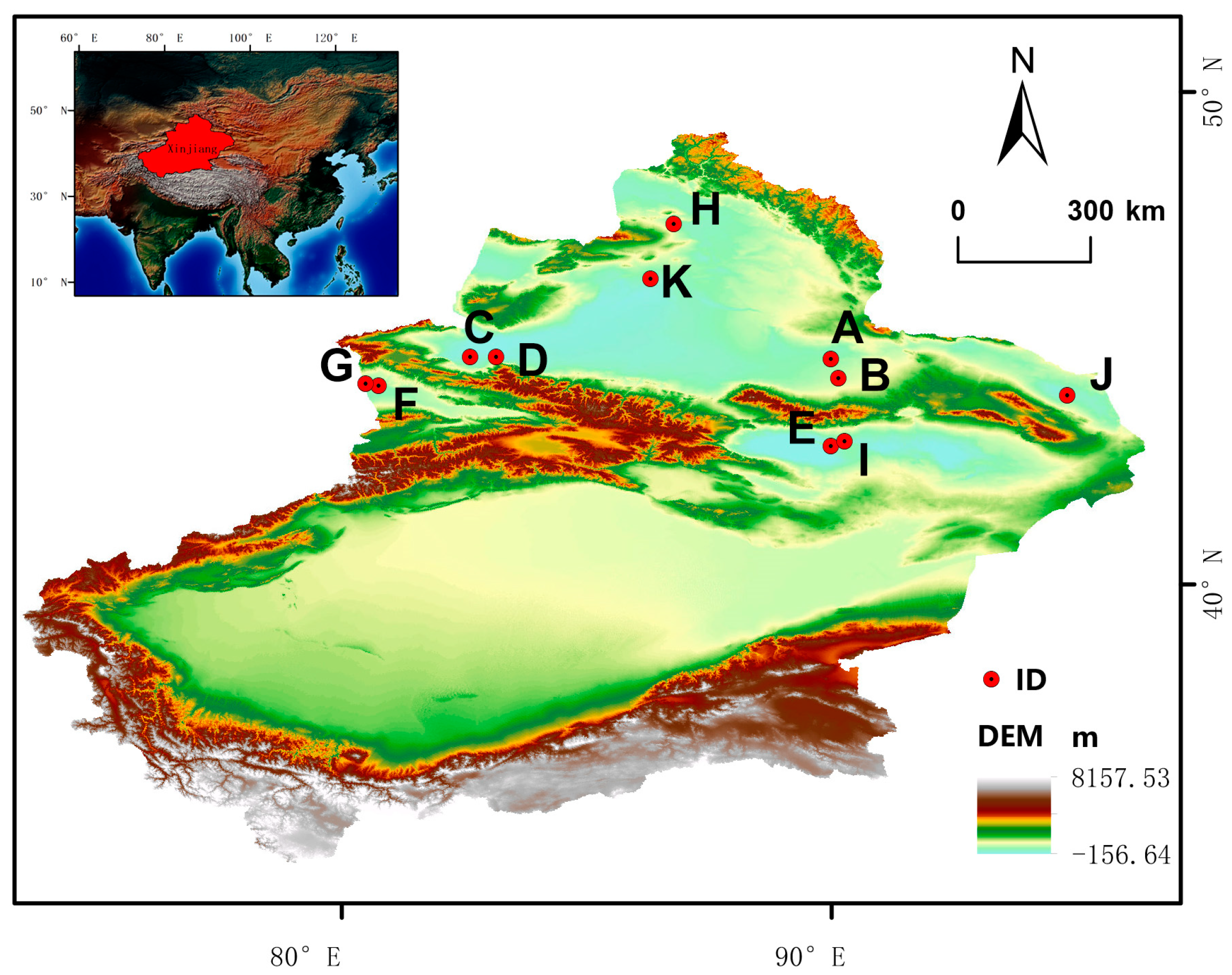
4.1. Experimental Materials
4.2. SSR Primer Development
4.3. DNA Extraction
4.4. Primer Screening
4.5. PCR Amplification
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, Y.; Pan, B.; Yan, C. Biodiversity and Distribution Pattern of Calligonum L. in Xinjiang. J. Arid. Land Resour. Environ. 2008, 22, 139–144. [Google Scholar]
- Li, A.; Gao, Z.; Mao, Z.; Editorial Committee of the Flora of China of Chinese Academy of Science. Flora of China; Science Publishing House: Beijing, China, 1998. [Google Scholar]
- Song, F.; Li, T.; Burgess, K.S.; Feng, Y.; Ge, X.-J. Complete plastome sequencing resolves taxonomic relationships among species of Calligonum L. (Polygonaceae) in China. BMC Plant Biol. 2020, 20, 261. [Google Scholar] [CrossRef]
- Purohit, C.S.; Kumar, R. A review on genus Calligonum L. (Polygonaceae) from India and report Calligonum crinitum an addition for Flora of India. J. Asia-Pac. Biodivers. 2020, 13, 319–324. [Google Scholar] [CrossRef]
- Brummitt, N.; Cui, Q.; Bai, P.; Liu, X.; Scott, B.; De Palma, A.; Araujo, A.C. Reducing the number of Data Deficient plant species. Syst. Biodivers. 2024, 22, 2302180. [Google Scholar] [CrossRef]
- Feng, Y.; Pan, B.; Shen, G. Seed Morphology of Calligonum Sect. EucaUigonum (Polygonaceae) in Xinjiang and Its Taxonomic Significance. Acta Bot. Yunnanica 2008, 30, 47–50. [Google Scholar]
- Liu, P.; Shi, W.; Wen, J.; Fayzullaevich, S.K.; Pan, B. A phylogeny of Calligonum L. (Polygonaceae) yields challenges to current taxonomic classifications. Acta Bot. Bras. 2021, 35, 310–322. [Google Scholar] [CrossRef]
- Lui, Y. Desert Flora of China; Science Publishing House: Beijing, China, 1985. [Google Scholar]
- Dong, Z.B.; Chen, G.T.; He, X.D.; Han, Z.W.; Wang, X.M. Controlling blown sand along the highway crossing the Taklimakan Desert. J. Arid Environ. 2004, 57, 329–344. [Google Scholar] [CrossRef]
- Lu, Q.; Li, X.; Xiao, H. Desert Ecosystem Observation Methods; China Environmental Science Publishing House: Beijing, China, 2004. [Google Scholar]
- Ci, J.; Zheng, L. Xinjiang Desert and Sand Fixation Afforestation; Xinjiang People’s Publishing House: Urumqi, China, 1963. [Google Scholar]
- Wang, J.; Zhang, W.; Xu, D. Influence of Calcium on the Plant Height, Photosynthesis, Fluorescence and Chlorophyll Content of Calligonum arborescensunder NaCl Stress. J. Desert Res. 2015, 35, 167–174. [Google Scholar]
- Tan, Y.; Pan, B.; Shiming, D.; Xu, G. Alpha diverstiy of four Calligonum L. Communities and soil interpretation in the Junggar Basin. J. Arid. Land Resour. Environ. 2009, 23, 136–142. [Google Scholar]
- An, Q.; Feng, Y.; Yang, Z.; Hu, L.; Wu, D.; Gong, G. Differences in Albizia odoratissima genetic diversity between Hainan Island and mainland populations in China. Front. Plant Sci. 2024, 15, 1369409. [Google Scholar] [CrossRef]
- Hedrick, P.W.; Miller, P.S. Conservation genetics: Techniques and fundamentals. Ecol. Appl. 1992, 2, 30–46. [Google Scholar] [CrossRef]
- Ahmad, R.; Malik, W.; Anjum, M.A. Genetic Diversity and Selection of Suitable Molecular Markers for Characterization of Indigenous Zizyphus Germplasm. Erwerbs-Obstbau 2019, 61, 345–353. [Google Scholar] [CrossRef]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Choudhary, M.; Singh, A.; Das, M.M.; Kumar, P.; Naliath, R.; Singh, V.; Kumar, B.; Rakshit, S. Morpho-physiological traits and SSR markers-based analysis of relationships and genetic diversity among fodder maize landraces in India. Mol. Biol. Rep. 2023, 50, 6829–6841. [Google Scholar] [CrossRef] [PubMed]
- Boccacci, P.; Aramini, M.; Ordidge, M.; van Hintum, T.J.L.; Marinoni, D.T.; Valentini, N.; Sarraquigne, J.-P.; Solar, A.; Rovira, M.; Bacchetta, L.; et al. Comparison of selection methods for the establishment of a core collection using SSR markers for hazelnut (Corylus avellana L.) accessions from European germplasm repositories. Tree Genet. Genom. 2021, 17, 48. [Google Scholar] [CrossRef]
- Qian, F.; Gao, Z.; Hu, L.; Wang, H. Characteristics of Crambe abyssinica Chloroplast Genome and Its Phylogenetic Relationship in Brassicaceae. Biotechnol. Bull. 2022, 38, 174–186. [Google Scholar]
- Wencai, Z. Application of SSR Molecular Marker in Soybean. J. Anhui Agric. Sci. 2007, 35, 3156–3158. [Google Scholar]
- Li, L.; Yan, C.; Xing, Y.; Wang, X.; Zhang, X. Utilization of SSR Marker in Genetic Diversity Analysis and Heterotic Grouping of Maize. Maize Sci. 2005, 13, 12–15. [Google Scholar]
- Mezzomo, P.; Mielniczki-Pereira, A.A.; Sausen, T.L.; Marinho, J.R.; Cansian, R.L. Molecular inferences about the genus Hypostomus Lacepede, 1803 (Siluriformes: Loricariidae): A review. Mol. Biol. Rep. 2020, 47, 6179–6192. [Google Scholar] [CrossRef]
- Algarawi, A.M.; Abd-Elgawad, M.E. Genetic diversity of closely related Calligounum species collected from Saudi habitats by analyzing the matK and rpoC1 genes, and SCoT and IRAP markers. Plant Biotechnol. Rep. 2025, 19, 55–66. [Google Scholar] [CrossRef]
- Gasmi, A.; Triki, T.; Benabderrahim, M.A.; Nagaz, K.; Guasmi, F. Assessing phenolic and molecular diversity of arta (Calligonum comosum L.), a wild Tunisian desert plant. S. Afr. J. Bot. 2022, 151, 166–174. [Google Scholar] [CrossRef]
- Shi, W.; Wen, J.; Zhao, Y.; Johnson, G.; Pan, B. Reproductive biology and variation of nuclear ribosomal ITS and ETS sequences in the Calligonum mongolicum complex (Polygonaceae). PhytoKeys 2017, 76, 71–88. [Google Scholar] [CrossRef]
- Hildebrand, C.E.; Torney, D.C.; Wagner, R.P. Informativeness of Polymorphic DNA Markers. Los Alimos Sricnce 1992, 2, 100–102. [Google Scholar]
- Duan, H.; Zhang, Q.; Tian, F.; Hu, Y.; Wang, C.; Lu, Y.; Yuan, H.; Yang, H.; Cui, G. Complete chloroplast genome of Calligonum mongolicum Turcz. and comparative analysis with other Calligonum species. J. Appl. Res. Med. Aromat. Plants 2022, 27, 100370. [Google Scholar] [CrossRef]
- Yang, B.; Li, L.; Liu, J.; Zhang, L. Plastome and phylogenetic relationship of the woody buckwheat Fagopyrum tibeticum in the Qinghai-Tibet Plateau. Plant Divers. 2021, 43, 198–205. [Google Scholar] [CrossRef]
- Zuo, X.; Song, Y.; Ma, X.; Yang, Y.; Wang, Y.; Guo, Z.; Zhu, X.; Liu, Y. Mining SSR Loci and Analysis the Genetic Diversity of Tartary Buckwheat Based on the Whole Genome Sequence. J. Agric. Sci. Technol. 2022, 24, 38–51. [Google Scholar]
- Booy, G.; Hendriks, R.J.J.; Smulders, M.J.M.; Van Groenendael, J.M.; Vosman, B. Genetic diversity and the survival of populations. Plant Biol. 2000, 2, 379–395. [Google Scholar] [CrossRef]
- Gaudeul, M.; Taberlet, P.; Till-Bottraud, I. Genetic diversity in an endangered alpine plant, Eryngium alpinum L. (Apiaceae), inferred from amplified fragment length polymorphism markers. Mol. Ecol. 2000, 9, 1625–1637. [Google Scholar] [CrossRef]
- Li, J.; Jin, Z.; Lou, W.; Li, J. Genetic diversity of Lithocarpus harlandii populations in three forest communities with different succession stage. Chin. J. Ecol. 2007, 26, 509–514. [Google Scholar] [CrossRef]
- Wei, J.; Li, X.; Xu, H.; Wang, Y.; Zang, C.; Xu, J.; Pei, X.; Zhao, X. Evaluation of the genetic diversity of Pinus koraiensis by EST-SSR and its management, utilization and protection. For. Ecol. Manag. 2022, 505, 119882. [Google Scholar] [CrossRef]
- Song, Y.; Cheng, Z.; Dong, Y.; Liu, D.; Bai, K.; Jarvis, D.; Feng, J.; Long, C. Diversity of Tartary Buckwheat (Fagopyrum tataricum) Landraces from Liangshan, Southwest China: Evidence from Morphology and SSR Markers. Agronomy 2022, 12, 1022. [Google Scholar] [CrossRef]
- Ekincialp, A.; Erdinc, C.; Turan, S.; Cakmakci, O.; Nadeem, M.A.; Baloch, F.S.; Sensoy, S. Genetic Characterization of Rheum ribes (Wild Rhubarb) Genotypes in Lake Van Basin of Turkey through ISSR and SSR Markers. Int. J. Agric. Biol. 2019, 21, 795–802. [Google Scholar]
- Dow, B.D.; Ashley, M.V. Microsatellite analysis of seed dispersal and parentage of saplings in bur oak, Quercus macrocarpa. Mol. Ecol. 1996, 5, 615–627. [Google Scholar] [CrossRef]
- Chabane, K.; Ablett, G.A.; Cordeiro, G.M.; Valkoun, J.; Henry, R.J. EST versus genomic derived microsatellite markers for genotyping wild and cultivated barley. Genet. Resour. Crop Evol. 2005, 52, 903–909. [Google Scholar] [CrossRef]
- Sabirhazi, G.; Borong, P. Variability of Fruit Characters of Calligonum roborovskii A. Los. Acta Bot. Boreali-Occident. Sin. 2008, 28, 370–374. [Google Scholar]
- Shi, W.; Pan, B.; Duan, S.; Kang, X. Difference of Fruit Morphological Characters of Calligonum mongolicum and Related Species. J. Desert Res. 2011, 31, 121–128. [Google Scholar]
- Feng, Y.; Wen, Z.; Li, Y.; Li, W. Climatic Factors Influencing Geographical Replacement in the Desert Genus Calligonum Sect. Medusa (Polygonaceae) in Xinjiang, China. Pak. J. Bot. 2021, 53, 1339–1342. [Google Scholar]
- Pan, H.; Feng, Y.; Wang, X.; Li, Y. Examination and comparison of the physiological characteristics of ten Calligonum species in a desert environment. Acta Prataculturae Sin. 2017, 26, 68–75. [Google Scholar]
- Zhang, Y.Z.; Zhang, Q.; Kang, X.S.; Pan, B.R.; Duan, S.M. Soil Characteristics of the Different Habitats of the Chinese Endemic Species Calligonum ebinuricum. Bull. Bot. Res. 2011, 31, 347–353. [Google Scholar]
- Rivers, M.C.; Brummitt, N.A.; Lughadha, E.N.; Meagher, T.R. Do species conservation assessments capture genetic diversity? Glob. Ecol. Conserv. 2014, 2, 81–87. [Google Scholar] [CrossRef]
- Sabirhazi, G.; Pan, B.-R. Chromosome numbers of three Calligonum species (Polygonaceae). Nord. J. Bot. 2009, 27, 284–286. [Google Scholar] [CrossRef]
- Gui, C.; Guan, S. Chromosomal geographic distribution of the genus Calligonum species (Polygonaceae) in Xinjiang. Arid. Zone Res. 1986, 02, 28–31. [Google Scholar]
- Beier, S.; Thiel, T.; Muench, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Adeyemo, O.A.; Ayodele, O.O.; Ajisafe, M.O.; Okinedo, U.E.; Adeoye, D.O.; Afanou, A.B.; Akinsemoyin, F.A.; Ogunjobi, O.O.; Kasali, O.J.; Chukwudiri, E.E. Evaluation of dark jute SSR markers and morphological traits in genetic diversity assessment of jute mallow (Corchorus olitorius L.) cultivars. S. Afr. J. Bot. 2021, 137, 290–297. [Google Scholar] [CrossRef]
- Vit, P.; Krak, K.; Douda, J.; Mandak, B. Microsatellite markers for Anthericum ramosum: Development, characterization, and cross-species amplification. Appl. Plant Sci. 2020, 8, e11323. [Google Scholar] [CrossRef]
- Glaubitz, J.C. CONVERT: A user-friendly program to reformat diploid genotypic data for commonly used population genetic software packages. Mol. Ecol. Notes 2004, 4, 309–310. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Carpio, A.J.; Apollonio, M.; Acevedo, P. Wild ungulate overabundance in Europe: Contexts, causes, monitoring and management recommendations Palabras clave. Mamm. Rev. 2021, 51, 95–108. [Google Scholar] [CrossRef]
- Muellner, D. fastcluster: Fast Hierarchical, Agglomerative Clustering Routines for R and Python. J. Stat. Softw. 2013, 53, 1–18. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Sonstebo, J.H.; Tollefsrud, M.M.; Myking, T.; Steffenrem, A.; Nilsen, A.E.; Edvardsen, O.M.; Johnskas, O.R.; El-Kassaby, Y.A. Genetic diversity of Norway spruce (Picea abies (L.) Karst.) seed orchard crops: Effects of number of parents, seed year, and pollen contamination. For. Ecol. Manag. 2018, 411, 132–141. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Francis, R.M. POPHELPER: An R package and web app to analyse and visualize population structure. Mol. Ecol. Resour. 2017, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Number |
|---|---|
| Total size of genome (Mb) | 1357.67 |
| Total number of identified SSRs | 380,328 |
| Total length of SSRs (bp) | 6,967,136 |
| Frequency (SSRs/Mb) | 280.13 |
| Density (bp/Mb) | 5131.69 |
| Repeat Type | Predominant Type | Number | Proportion (%) | Frequency (SSRs/Mb) | Total Length (bp) | Average Length (bp) |
|---|---|---|---|---|---|---|
| Mono | A/T | 204,094 | 65.28 | 150.33 | 2,514,452 | 12.32 |
| Di | AT/TA | 99,173 | 31.72 | 73.05 | 1,422,190 | 14.34 |
| Tri | AAT/ATT | 6527 | 2.09 | 4.81 | 43,106 | 6.60 |
| Tetra | ACAT/ATGT | 1129 | 0.36 | 0.83 | 9221 | 8.17 |
| Penta | - | 769 | 0.25 | 0.57 | 4216 | 5.48 |
| Hexa | - | 939 | 0.30 | 0.69 | 9462 | 10.08 |
| Locus | Na | Ne | I | Ho | He | PIC |
|---|---|---|---|---|---|---|
| Sgz001 | 3.00 | 2.02 | 0.77 | 0.24 | 0.44 | 0.68 |
| Sgz002 | 1.90 | 1.51 | 0.43 | 0.35 | 0.26 | 0.60 |
| Sgz004 | 3.09 | 2.08 | 0.85 | 0.66 | 0.49 | 0.68 |
| Sgz005 | 3.27 | 2.30 | 0.88 | 0.28 | 0.49 | 0.70 |
| Sgz006 | 5.09 | 3.41 | 1.23 | 0.61 | 0.61 | 0.86 |
| Sgz007 | 2.45 | 1.82 | 0.66 | 0.30 | 0.39 | 0.64 |
| Sgz010 | 2.00 | 1.47 | 0.41 | 0.02 | 0.23 | 0.48 |
| Sgz011 | 2.63 | 2.16 | 0.81 | 0.74 | 0.48 | 0.55 |
| Sgz012 | 3.54 | 2.74 | 1.02 | 0.60 | 0.55 | 0.82 |
| Sgz014 | 2.27 | 1.65 | 0.49 | 0.11 | 0.29 | 0.49 |
| Sgz016 | 1.81 | 1.45 | 0.38 | 0.01 | 0.24 | 0.51 |
| Sgz017 | 4.36 | 3.27 | 1.26 | 0.97 | 0.67 | 0.75 |
| Sgz019 | 1.72 | 1.45 | 0.34 | 0.00 | 0.21 | 0.51 |
| Sgz020 | 2.18 | 1.70 | 0.40 | 0.02 | 0.22 | 0.62 |
| Sgz021 | 1.81 | 1.51 | 0.38 | 0.00 | 0.23 | 0.56 |
| Sgz022 | 2.27 | 1.49 | 0.43 | 0.05 | 0.25 | 0.48 |
| Sgz023 | 2.18 | 1.70 | 0.40 | 0.00 | 0.21 | 0.61 |
| Mean | 2.68 | 1.99 | 0.66 | 0.29 | 0.37 | 0.62 |
| Species | Na | Ne | I | Ho | He | uHe | Private Alleles |
|---|---|---|---|---|---|---|---|
| C. colubrinum | 2.00 | 1.76 | 0.52 | 0.25 | 0.33 | 0.37 | 0.00 |
| C. leucocladum | 2.23 | 1.88 | 0.56 | 0.27 | 0.32 | 0.37 | 0.11 |
| C. junceum | 2.82 | 1.98 | 0.74 | 0.29 | 0.43 | 0.46 | 0.47 |
| C. ebinuricum | 1.94 | 1.71 | 0.46 | 0.30 | 0.28 | 0.32 | 0.05 |
| C. pumilum | 2.52 | 1.96 | 0.66 | 0.27 | 0.38 | 0.42 | 0.11 |
| C. trifarium | 3.94 | 2.68 | 0.98 | 0.40 | 0.51 | 0.54 | 1.17 |
| C. arborescens | 2.05 | 1.86 | 0.55 | 0.29 | 0.34 | 0.39 | 0.00 |
| C. jeminaicum | 2.29 | 1.87 | 0.60 | 0.32 | 0.36 | 0.41 | 0.23 |
| C. mongolicum | 6.11 | 3.33 | 1.30 | 0.36 | 0.64 | 0.66 | 2.05 |
| C. gobicum | 0.76 | 0.66 | 0.12 | 0.07 | 0.07 | 0.08 | 0.05 |
| C. roborowskii | 2.82 | 2.15 | 0.72 | 0.36 | 0.40 | 0.43 | 0.41 |
| Mean | 2.68 | 1.99 | 0.66 | 0.29 | 0.37 | 0.41 | 0.42 |
| Source | df | SS | MS | Est. Var. | % | P | Nm |
|---|---|---|---|---|---|---|---|
| Among Species | 10 | 370.38 | 37.04 | 1.45 | 5% | 0.05 | 0.38 |
| Within Species | 69 | 1869.32 | 27.10 | 27.09 | 95% |
| A | B | C | D | E | F | G | H | I | J | K | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.00 | A | ||||||||||
| 0.63 | 0.00 | B | |||||||||
| 0.33 | 0.78 | 0.00 | C | ||||||||
| 0.28 | 0.52 | 0.54 | 0.00 | D | |||||||
| 0.36 | 1.00 | 0.26 | 0.44 | 0.00 | E | ||||||
| 0.42 | 0.85 | 0.22 | 0.46 | 0.20 | 0.00 | F | |||||
| 0.31 | 0.56 | 0.56 | 0.13 | 0.47 | 0.39 | 0.00 | G | ||||
| 0.20 | 0.86 | 0.57 | 0.36 | 0.45 | 0.42 | 0.33 | 0.00 | H | |||
| 0.21 | 0.73 | 0.26 | 0.26 | 0.22 | 0.24 | 0.24 | 0.22 | 0.00 | I | ||
| 1.09 | 0.85 | 1.92 | 1.01 | 1.83 | 1.63 | 0.88 | 1.15 | 1.33 | 0.00 | J | |
| 0.50 | 0.19 | 0.83 | 0.72 | 0.90 | 0.85 | 0.63 | 0.51 | 0.65 | 0.76 | 0.00 | K |
| ID | Species | Number | Latitude (N) | Longitude (E) | Elevation (m) |
|---|---|---|---|---|---|
| A | Calligonum colubrinum | 4 | 44.58 | 89.98 | 520.51 |
| B | Calligonum leucocladum | 4 | 44.19 | 90.14 | 755.91 |
| C | Calligonum junceum | 8 | 44.62 | 82.63 | 277.57 |
| D | Calligonum ebinuricum | 4 | 44.62 | 83.16 | 370.29 |
| E | Calligonum pumilum | 5 | 42.81 | 89.98 | 129.20 |
| F | Calligonum trifarium | 8 | 44.03 | 80.75 | 564.00 |
| G | Calligonum arborescens | 4 | 44.08 | 80.49 | 639.00 |
| H | Calligonum jeminaicum | 4 | 47.32 | 86.77 | 574.20 |
| I | Calligonum mongolicum | 23 | 42.89 | 90.25 | 387.00 |
| J | Calligonum gobicum | 4 | 43.84 | 94.81 | 361.87 |
| K | Calligonum roborowskii | 12 | 46.21 | 86.31 | 438.23 |
| Locus Name | Primers (5′ to 3′) | Allele Size (bp) | Ta (°C) |
|---|---|---|---|
| Sgz001 | F: ACCTCTTCAGTAGTTGACCTCA | 195 | 59 |
| R: TCCCTAAGGCTAGTTGGAGGA | |||
| Sgz002 | F: CAACTGCCGACACACAGATG | 175 | 59 |
| R: AGTTTGCTTGGTTGGGTCTGA | |||
| Sgz004 | F: AGCATACAAGTGTCGGCCTC | 178 | 59 |
| R: TGGTCTTGAGTGCGGATGAT | |||
| Sgz005 | F: TCGAGTTAGTCCCGGATCGT | 131 | 59 |
| R: ACTTCACCATCTCAATCACCTCT | |||
| Sgz006 | F: TCCATGTCTCCAAACGGCTG | 140 | 60 |
| R: TCCCTCCACATCTCACGAATC | |||
| Sgz007 | F: GAAGGGTAAGCAGAAACTGGGA | 190 | 60 |
| R: TCGCAGAACCACAACGATCC | |||
| Sgz010 | F: CCTACGTCTGAGGCGTTCTC | 189 | 60 |
| R: AGCAAAGAGGGCTTAGTTCTTCA | |||
| Sgz011 | F: ACGTCGTGCTTCTAAAGGGT | 122 | 57 |
| R: ACTGACAATTATATGGAGGCACA | |||
| Sgz012 | F: GGTTACAGCGCCAGGAATCT | 144 | 59 |
| R: TGCTACACTTGTACTTATACCCTGA | |||
| Sgz014 | F: ATACGTGGCACAGAAGTCCG | 165 | 60 |
| R: TGGGTGGAAAGGTGAATCGAG | |||
| Sgz016 | F: AGGCGTTAACACCCGAAACT | 177 | 58 |
| R: AGCAAGGAGGGCTTAGTTCT | |||
| Sgz019 | F: CCGCCCGAAACTCTGTGTT | 169 | 59 |
| R: AGCAAAGAGGGCTTAGTTCTTCA | |||
| Sgz020 | F: CCGCCCGAAACTCTGTGTT | 169 | 59 |
| R: AGCAAAGAGGGCTTAGTTCTTCA | |||
| Sgz021 | F: CCGCCCGAAACTCTGTGTT | 169 | 59 |
| R: AGCAAAGAGGGCTTAGTTCTTCA | |||
| Sgz022 | F: TTCCCGCCCGAAACTTTGT | 167 | 58 |
| R: AGGGCTTTGTTCTTCATAGAGCA | |||
| Sgz023 | F: CCGCCCGAAACTCTGTGTT | 169 | 59 |
| R: AGCAAAGAGGGCTTAGTTCTTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.; Li, X.; Liu, S.; Wang, X.; Zhang, D.; Pan, B.; Wei, M.; Wang, J.; Shi, W. Genetic Diversity of Calligonum L. Species Analyzed Using Newly Developed Genomic Simple Sequence Repeats and Their Application to Conservation. Diversity 2025, 17, 196. https://doi.org/10.3390/d17030196
Su J, Li X, Liu S, Wang X, Zhang D, Pan B, Wei M, Wang J, Shi W. Genetic Diversity of Calligonum L. Species Analyzed Using Newly Developed Genomic Simple Sequence Repeats and Their Application to Conservation. Diversity. 2025; 17(3):196. https://doi.org/10.3390/d17030196
Chicago/Turabian StyleSu, Jiayi, Xuerong Li, Shiqing Liu, Xiyong Wang, Daoyuan Zhang, Borong Pan, Mengmeng Wei, Jancheng Wang, and Wei Shi. 2025. "Genetic Diversity of Calligonum L. Species Analyzed Using Newly Developed Genomic Simple Sequence Repeats and Their Application to Conservation" Diversity 17, no. 3: 196. https://doi.org/10.3390/d17030196
APA StyleSu, J., Li, X., Liu, S., Wang, X., Zhang, D., Pan, B., Wei, M., Wang, J., & Shi, W. (2025). Genetic Diversity of Calligonum L. Species Analyzed Using Newly Developed Genomic Simple Sequence Repeats and Their Application to Conservation. Diversity, 17(3), 196. https://doi.org/10.3390/d17030196