2. The Data
There are eight species of the genus Podarcis distributed in Greece, of which five are endemic to islands and Peloponnesos. According to their phylogenetic relationships, they can be divided into three groups: The P. erhardii group (P. erhardii, P. peloponnesiaca, P. cretensis, P. levendis), the P. taurica group (P. taurica, P. milensis, P. gaigeae) and a last group with a single representative P. muralis.
Phylogenetic relationships among the major lineages of Podarcis and consequently among the groups are not clear.
The study of the
P. erhardii’s group [
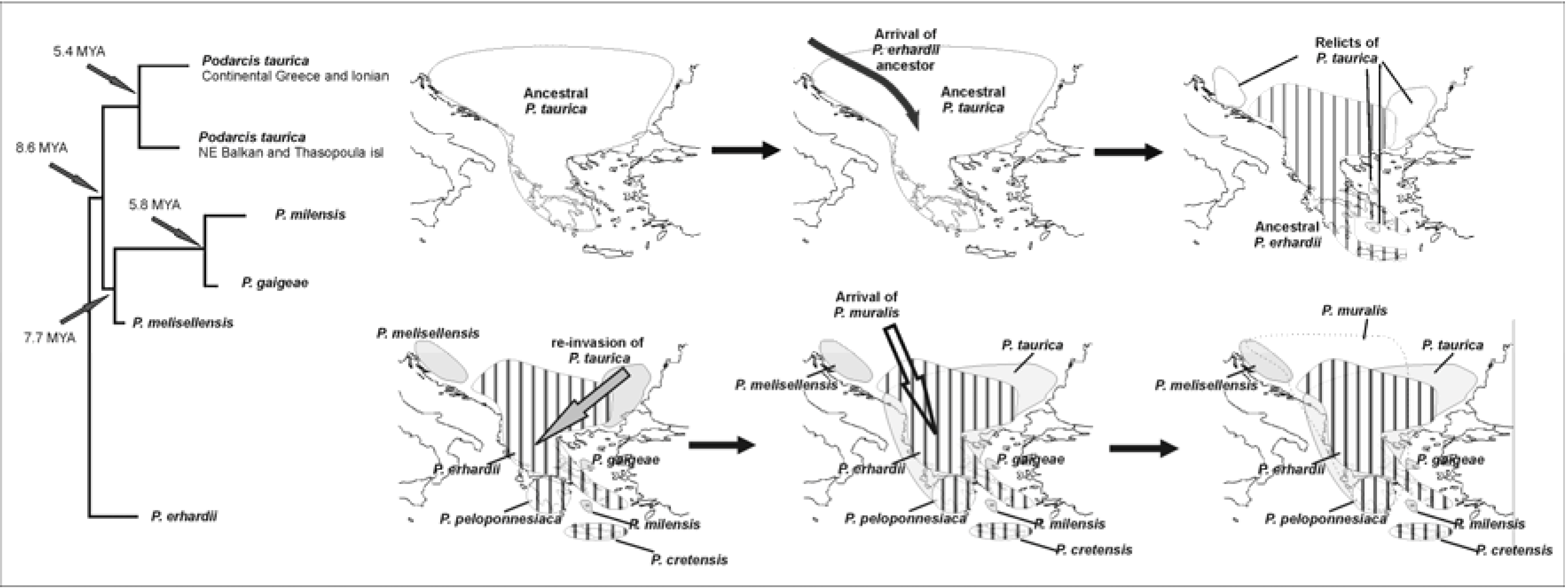
15,
16] revealed hidden diversity and concluded with a plausible biogeographic scenario. In brief, the group’s ancestral form appears to have reached Greece from the north and initially split in two lineages. Consequent vicariant events formed the distinct taxa we now recognize (
Figure 4).
The second group (
P. taurica), largely overlapping with the previous one in time and space, has a similar history in the area [
4]. The interaction of the two species groups appears as another factor that influenced present day distributions (
Figure 5).
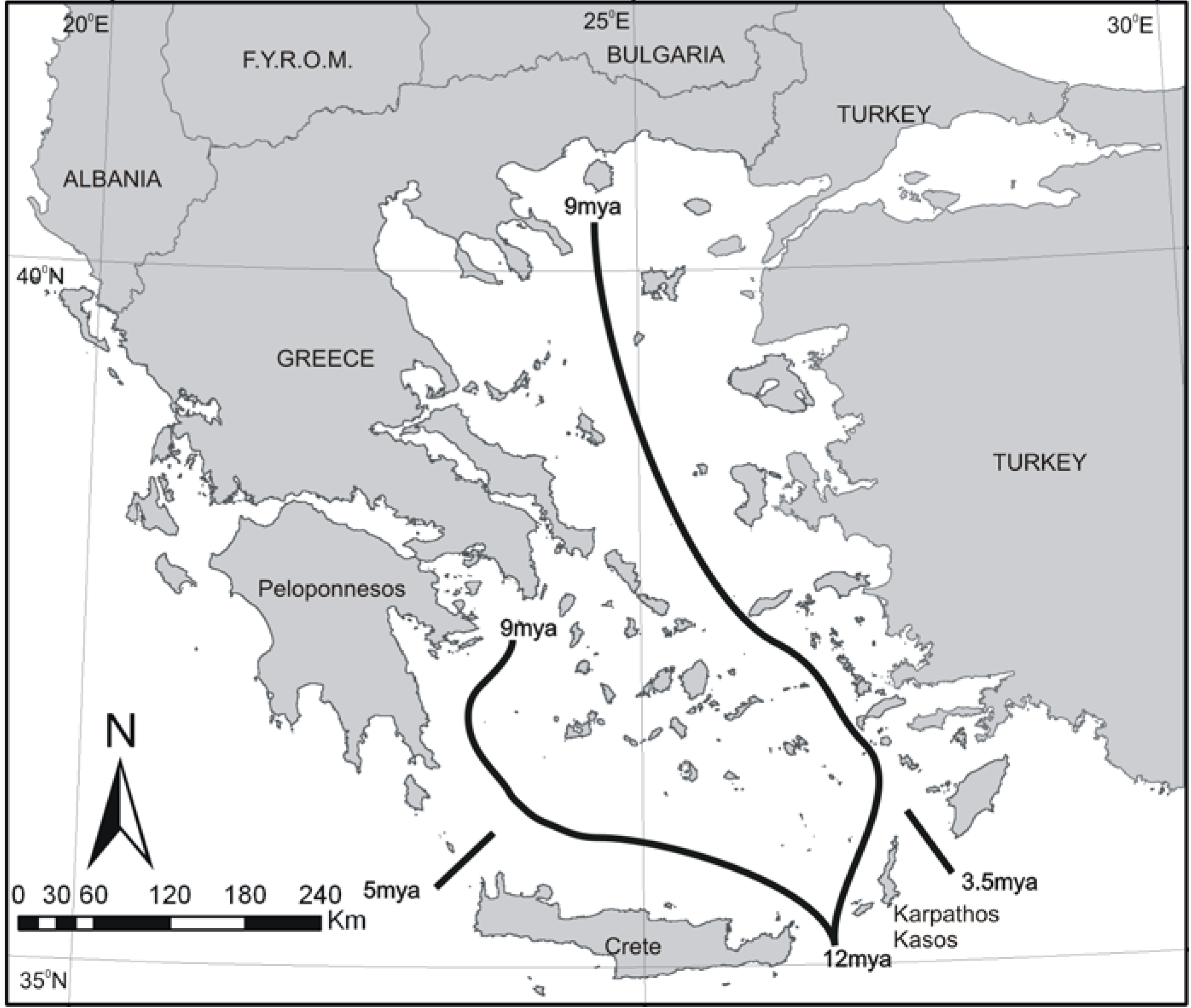
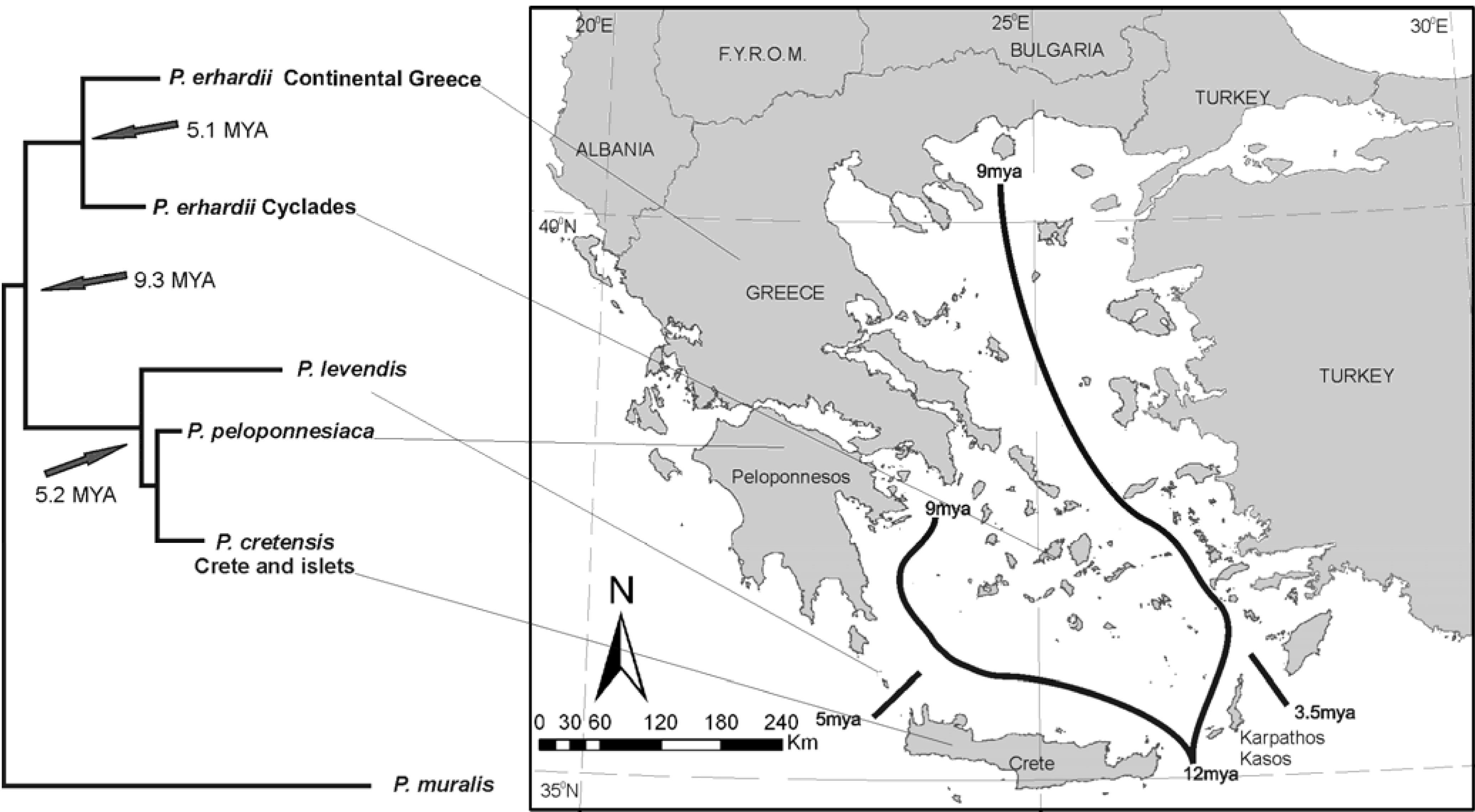
Figure 4.
Phylogenetic relationships of P. erhardii group. Note that the taxa respect the 12-9 MYA boundaries and that contemporary species differentiate around 5 MYA, i.e., near the Messinian crisis.
Figure 4.
Phylogenetic relationships of P. erhardii group. Note that the taxa respect the 12-9 MYA boundaries and that contemporary species differentiate around 5 MYA, i.e., near the Messinian crisis.
Figure 5.
Phylogenetic relationships of
P. taurica group and the proposed interaction with the
P. erhardii group after [
17]. Taxa respect the 12-9 MYA boundaries and contemporary species, such as
P. erhardii, differentiate near the Messinian crisis. Dotted line depicts proposed dispersal route. Grey arrow suggests a second invasion from the north-east.
Figure 5.
Phylogenetic relationships of
P. taurica group and the proposed interaction with the
P. erhardii group after [
17]. Taxa respect the 12-9 MYA boundaries and contemporary species, such as
P. erhardii, differentiate near the Messinian crisis. Dotted line depicts proposed dispersal route. Grey arrow suggests a second invasion from the north-east.
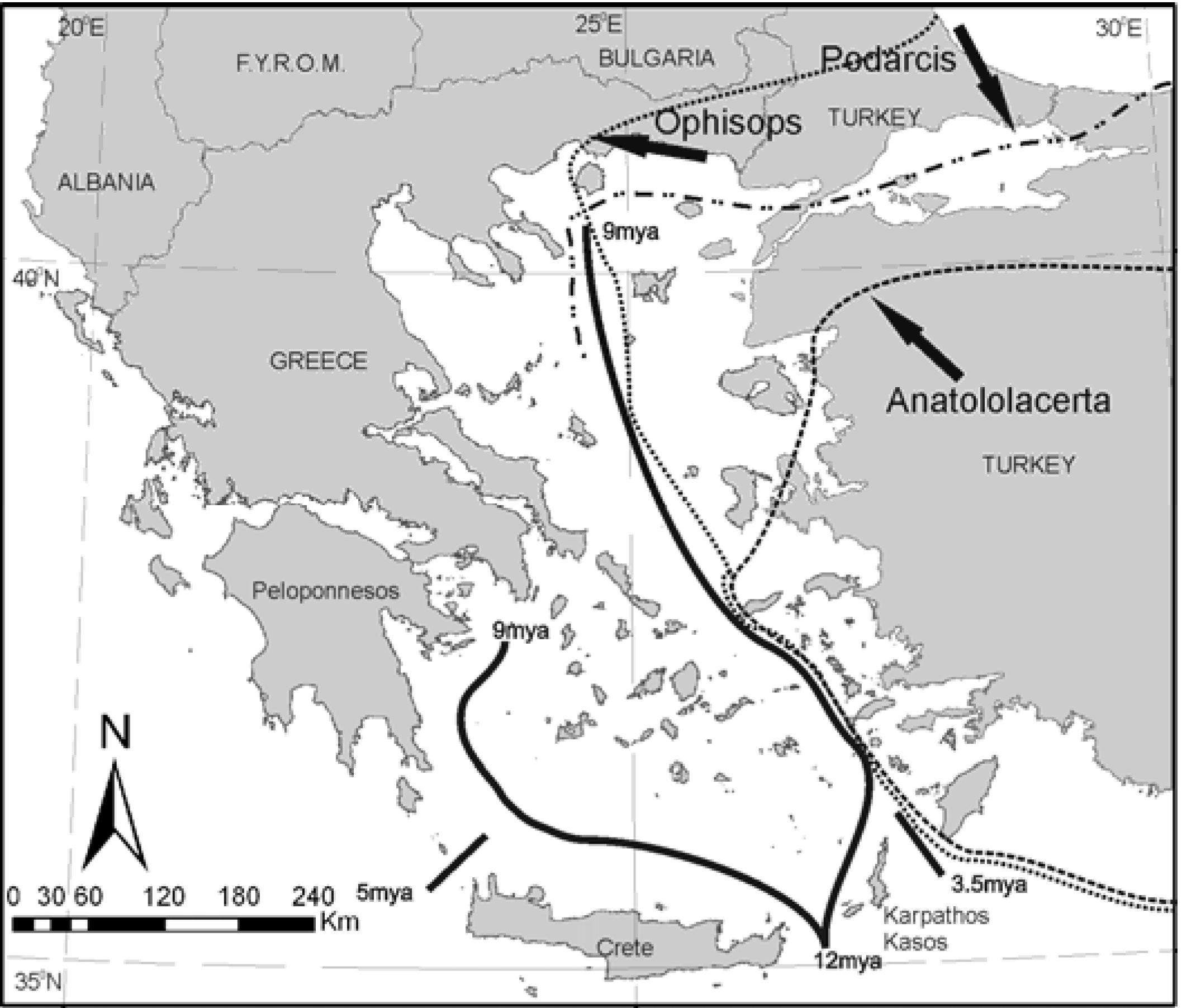
The genus
Podarcis is considered to have evolved in south-west Europe and expanded eastwards [
18]. However, it never made it into Asia, blocked at least in its southern range by the forming of the MAT. A different lacertid,
Ophisops elegans, which arrived in the area from Asia Minor, hardly made it to Europe in Thrace, NE Greece [
19]. It is possible that
O. elegans arrived in the area later than 3.5 MYA since it is “unexpectedly absent from Kasos and Karpathos” as observed in [
14].
A different pattern is shown by another lacertid genus,
Anatololacerta. The genus is distributed in west and south Turkey, but only on some south eastern Aegean islands and not the ones in the north (
Figure 6). A possible explanation is a very recent northwards dispersal of
Anatololacerta, after the splitting of e.g., Lesvos and Khios, which split from Anatolia after the last ice age
circa 12 KYA [
20]. The relative distribution of the three genera is shown in
Figure 6.
Figure 6.
Relative position of the distribution limits of three lacertid genera in the Aegean.
Figure 6.
Relative position of the distribution limits of three lacertid genera in the Aegean.
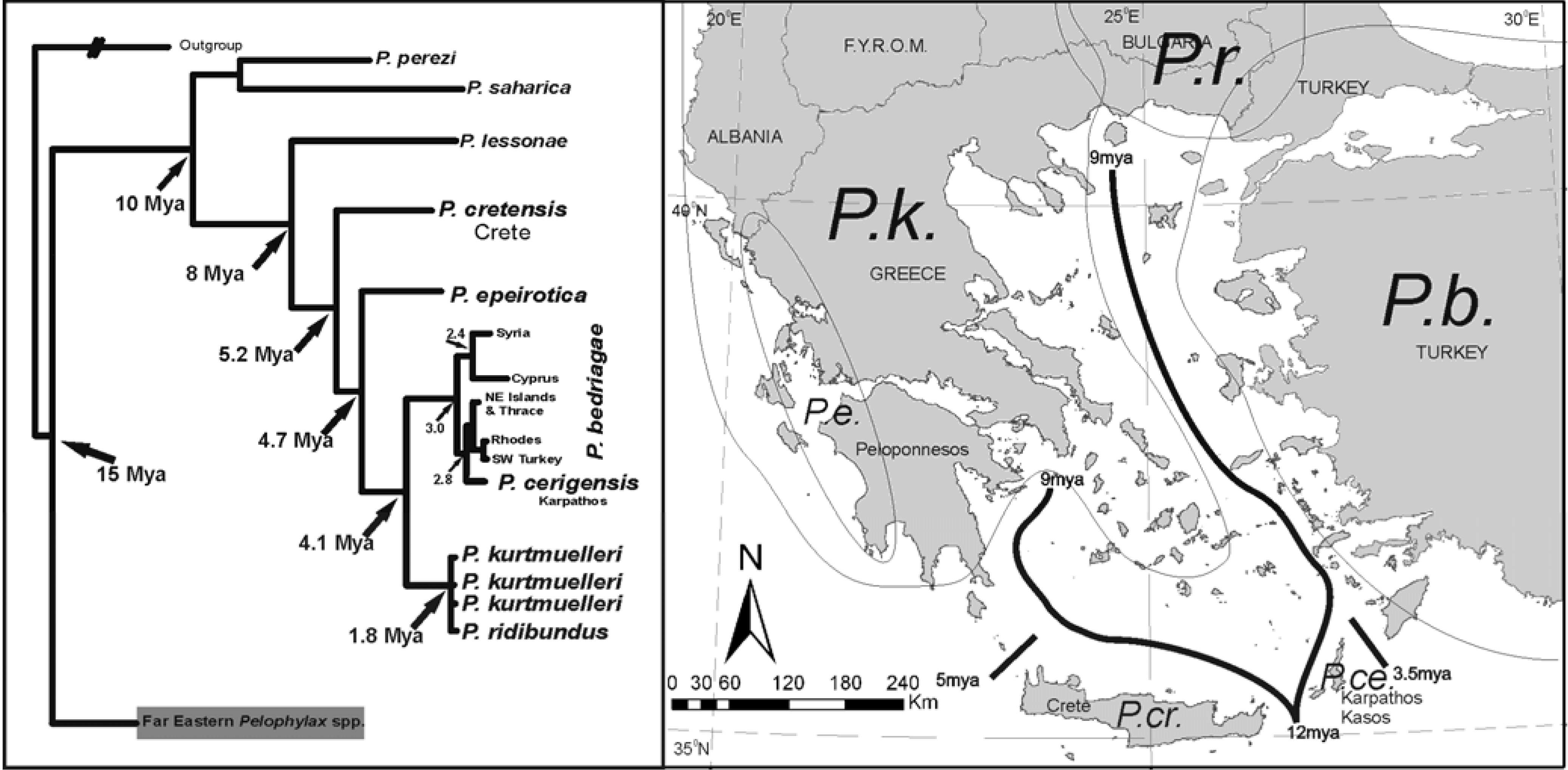
The case of
Pelophylax spp. in the area is more complex, involving several refugia, recolonizations and in some cases possible overseas dispersal. The taxonomy of the group remains uncertain despite the plethora of morphological, bioacoustic, protein and DNA data analyzed in several works. A tentative explanation of the observed pattern is proposed in
Figure 7.
The splitting of southern European taxa of the
Bufo viridis group occurred between Lower and Middle Pliocene,
i.e., after the Messinian crisis [
24], a pattern similar to the one described for
Pelophylax (
Figure 7). Later retreats and recolonizations have occurred in a different manner in
Bufo than the one proposed for
Pelophylax, in which a northern refuge is proposed to have played an important role. The broader biogeographic pattern, which describes the evolution of both groups however is similar to the one showed for many Palaearctic animal and plant species: extinction and recolonization in higher latitudes and altitudinal shifts and complex refugia nearer the tropics [
25]. A third group of anuran amphibians, belonging to the genus
Hyla, was until now considered uniform in the area, as in the case of
Bufo toads.
Hyla spp. have been found to have a similar pattern with the two previous groups,
i.e., different taxa on the two sides of the MAT, without however having the split between the two taxa dated yet [
26].
Figure 7.
Phylogenetic relationships and distribution of
Pelophylax spp. in the area after [
21]. Despite the complex relationships, taxa still respect the 12-9 MYA boundaries. The differentiation of contemporary species once again begins near the Messinian crisis, but several later speciation incidents follow. Vicariant events followed by recolonization episodes appear as the prevailing scenario for the pattern observed. The case of
Pelophylax supports the hypothesis of refugia within refugia [
22,
23] (and references therein).
Figure 7.
Phylogenetic relationships and distribution of
Pelophylax spp. in the area after [
21]. Despite the complex relationships, taxa still respect the 12-9 MYA boundaries. The differentiation of contemporary species once again begins near the Messinian crisis, but several later speciation incidents follow. Vicariant events followed by recolonization episodes appear as the prevailing scenario for the pattern observed. The case of
Pelophylax supports the hypothesis of refugia within refugia [
22,
23] (and references therein).
Another amphibian group, Salamanders, of the genera
Salamandra,
Mertensiella and
Lyciasalamandra, exhibit a pattern predominantly formed by vicariant events. Weisrock [
27] suggested an initial vicariant event split
Salamandra from
Mertensiella (the latter at the time of the publication contained the genus
Lyciasalamandra) at
circa 12MYA, a date which corresponds to the opening of the MAT. A little earlier,
i.e., 13MYA, is the time European
Salamandra spp. split with
S. infraimmaculata [
28].
The differentiation within
Lyciasalamandra (
Mertensiella luschani group at the time) is estimated to have begun at 5.8 MYA and the authors contribute it to the intense folding of the Anatolian plateau, which in turn caused rapid isolations among populations. The latter appears as a polytomy in phylogenetic trees [
29]. However, a simple vicariant event may not explain this situation when regarded in further detail [
30]: The species from Kasos and Karpathos in the Aegean appears to have sister species relationships (in regards to mt DNA) with the easternmost species in the Beydaglari Mountains some 300 km to the east, and crossing the distributions of all the remaining congeneric species. Veith and coworkers [
29] propose dispersal events (namely rafting) to explain the discordance between mt DNA relationships and geography.
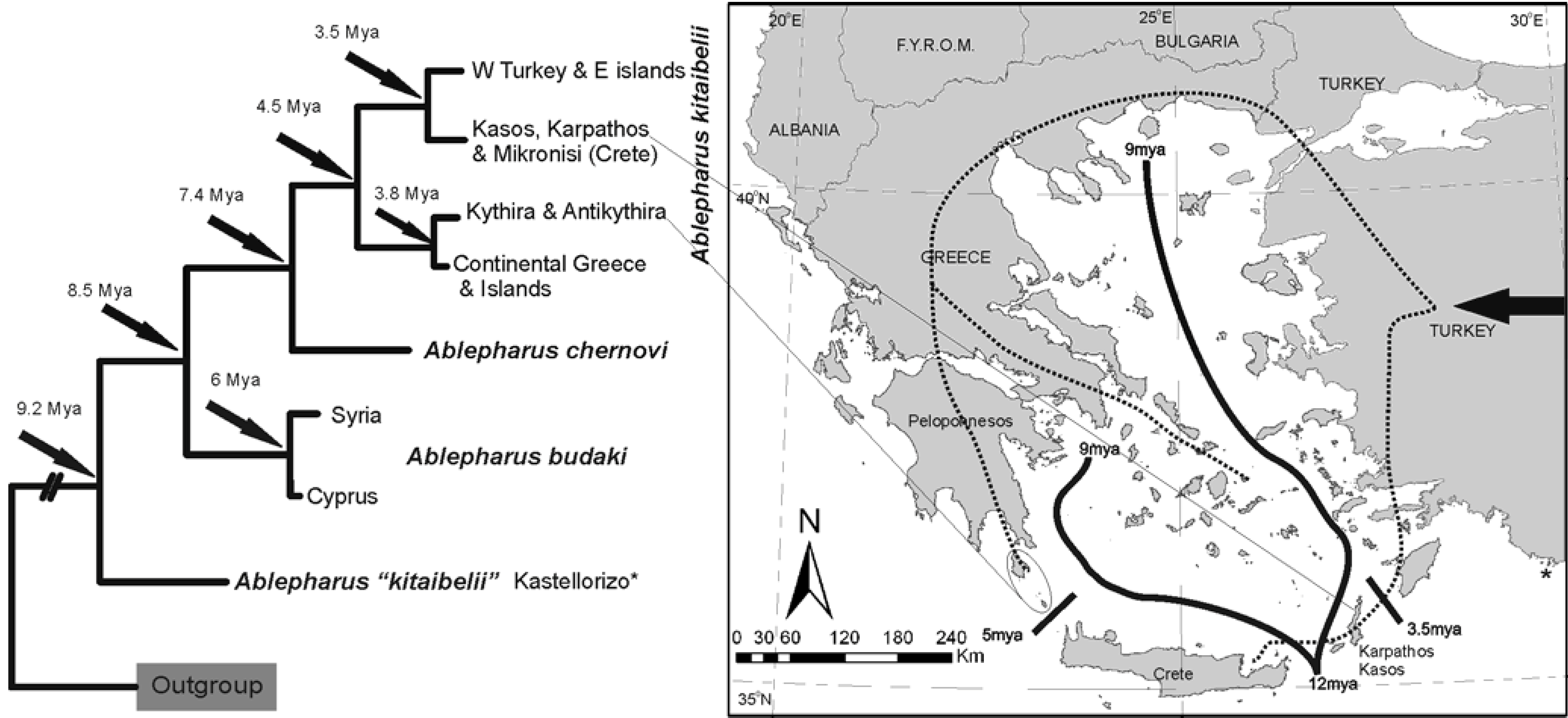
The snake-eyed skink,
Ablepharus kitaibelii, is considered to have arrived in the area from the east and is distributed throughout Greece with the exception of Crete, reaching as far north as the northern Balkans. The uniformity of the nominal subspecies distribution over a vast area was challenged [
31]. Phylogeographic relationships and the biogeographical scenario for its distribution in the Aegean are shown in
Figure 8.
Figure 8.
Phylogenetic relationships and distribution of Ablepharus taxa in the area. The results suggest a dispersal (dotted lines) versus a vicariant pattern of distribution. Taxa respect the 12-9 MYA boundaries.
Figure 8.
Phylogenetic relationships and distribution of Ablepharus taxa in the area. The results suggest a dispersal (dotted lines) versus a vicariant pattern of distribution. Taxa respect the 12-9 MYA boundaries.
The differentiation of Aegean populations of Ablepharus once again begins soon after the Messinian crisis and two isolation episodes follow during the Pliocene. The population of Kastellorizo (*), presumably related to populations of south-west Turkey, render A. kitaibelli paraphyletic. Detailed sampling in that area may reveal further hidden diversity.
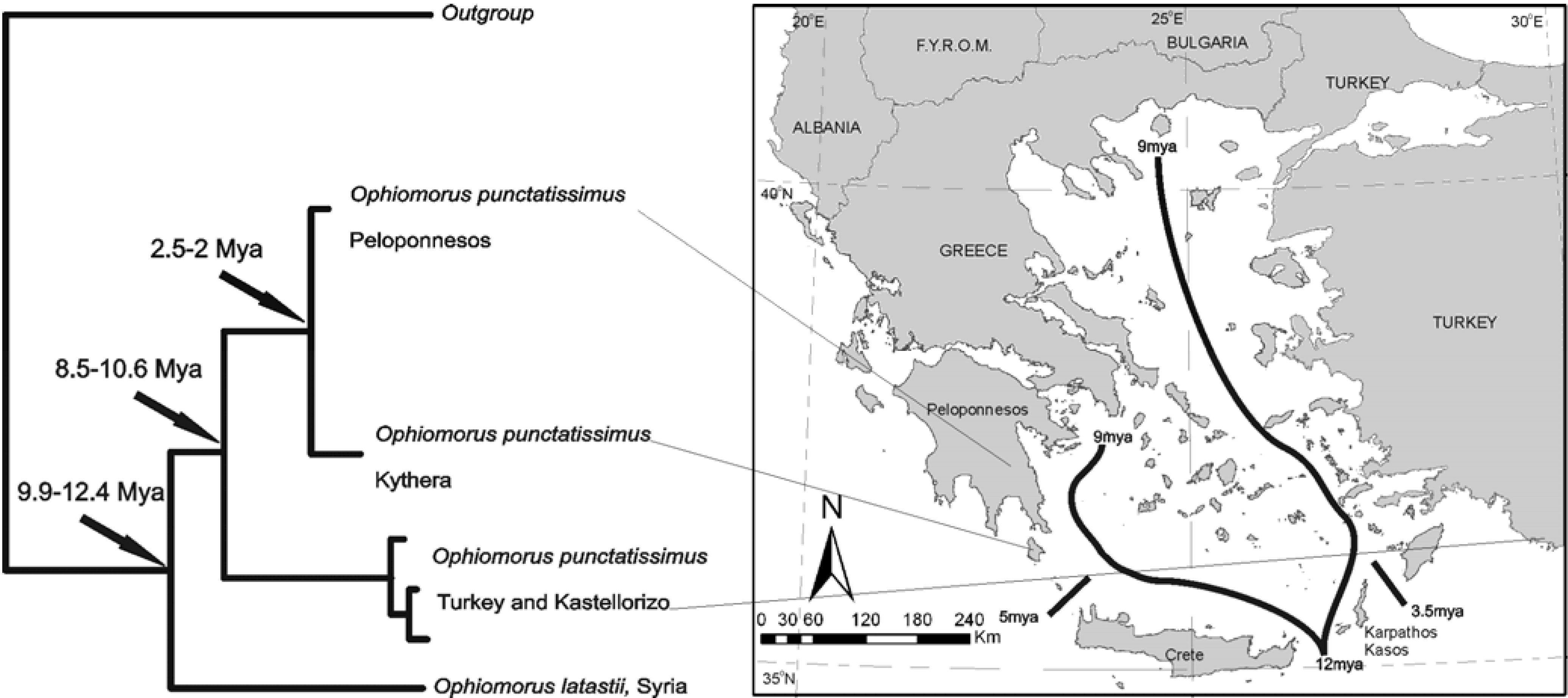
A species with a disjunct distribution,
Ophiomorus punctatissimus, was examined [
32] to explore its phylogeographic relationships. Populations are restricted to Peloponnesos, the island of Kythera, a single spot in the south-east of continental Greece, a small area in southwest Turkey and the nearby Greek island of Kastellorizo (
Figure 9).
Another species,
Cyrtopodion kotschyi, appears to have an equally long history in the area [
33]. Unlike
Ophiomorus, which remained restricted locally,
C. kotschyi is widespread throughout the area. It has dense populations on small islands and islets and is restricted on continental areas and big islands (e.g., Crete). The authors find a high intraspecific variation in CO1 (ranging from 1.2–21.6%) and no geographic coherence of the major clades. This result possibly indicates the need for more samples and more molecular markers to better understand the animal’s history in the area, as in the case of yet another pre–Messinian occupant of the area, the dice snake (
Natrix tessellata). An elaborate explanation of the dice snake’s history, involving Asian origin, several refugia and instances of recession and recolonization has been proposed [
34].
Two cases of human introductions have been described recently. The first regards the Moorish gecko,
Tarentola mauritanica [
35]. The authors found mt DNA fragments of all European populations to be identical to the animals from Tunisia. The species originates from Northern Africa where mt DNA is differentiated in a geographically structured pattern.
Figure 9.
Phylogenetic relationships and distribution of Ophiomorus taxa in the area. The most plausible scenario is that the species existed in the area that was separated into two with the 12-9 MYA splitting of the Aegean. If this hypothesis is correct, fossils of the species are expected to be found in Cyclades—possibly even Crete.
Figure 9.
Phylogenetic relationships and distribution of Ophiomorus taxa in the area. The most plausible scenario is that the species existed in the area that was separated into two with the 12-9 MYA splitting of the Aegean. If this hypothesis is correct, fossils of the species are expected to be found in Cyclades—possibly even Crete.
The Mediterranean distribution of another gecko,
Hemidactylus turcicus, which originates from the Near East (Jordan), has been shown to be the result of human introductions [
36].
Apart from their biological interest, the two latter cases bring history in to play: which trade and at what time in each case, caused such an extensive, human induced, dispersal? Corti and coworkers [
37] have presented a preliminary approach to this issue.
The Occelated skink,
Chalcides occelatus, is a somewhat different case from the two geckos as both dispersal and human introductions appear to be responsible for its present distribution [
38]. The species originated in Morocco from where during Early Pliocene it expanded eastwards. However mt DNA relationships appear having a geographical structure up to Cyrenaica in Libya. From there and eastwards, population relationships do not seem to follow any geographic pattern indicating possible involuntary human introductions.
A much more peculiar case of possible human introduction regards
Hierophis viridiflavus, a species grossly distributed from the Pyrenees through France and Italy to the Northern Dalmatian coast. The long discussed
Coluber gyarosensis, a snake found exclusively on the non inhabited island of Gyaros in the Cyclades, cannot be distinguished , in regards to CO1 and 12s rRNA genes as well as morphological characters, from
H. viridiflavus [
39]. Our hypothesis is that there is little chance that the species was involuntarily introduced to Gyaros as it is possible for the two geckos and the skink species. Snake rituals were common during the antiquity; even today there are snake rituals connected to Greek orthodox ceremonies in the island of Kephallonia involving
Telescopus fallax, hence the case of
H. viridiflavus represents yet another riddle for archaeologists and historians to solve.
Eurasian Vipers of the genera
Montivipera and
Macrovipera from the one side and
Vipera and
Daboia from the other appear to have split earlier than 20 MYA [
40,
41,
42]. In what concerns
Vipera ammodytes, a post Messinian differentiation in four distinct groups within the Balkans is proposed [
41], further supporting the idea of “refugia within refugia”. It is interesting to compare this with the proposed phylogeny of the second clade (
Montivipera/Macrovipera).
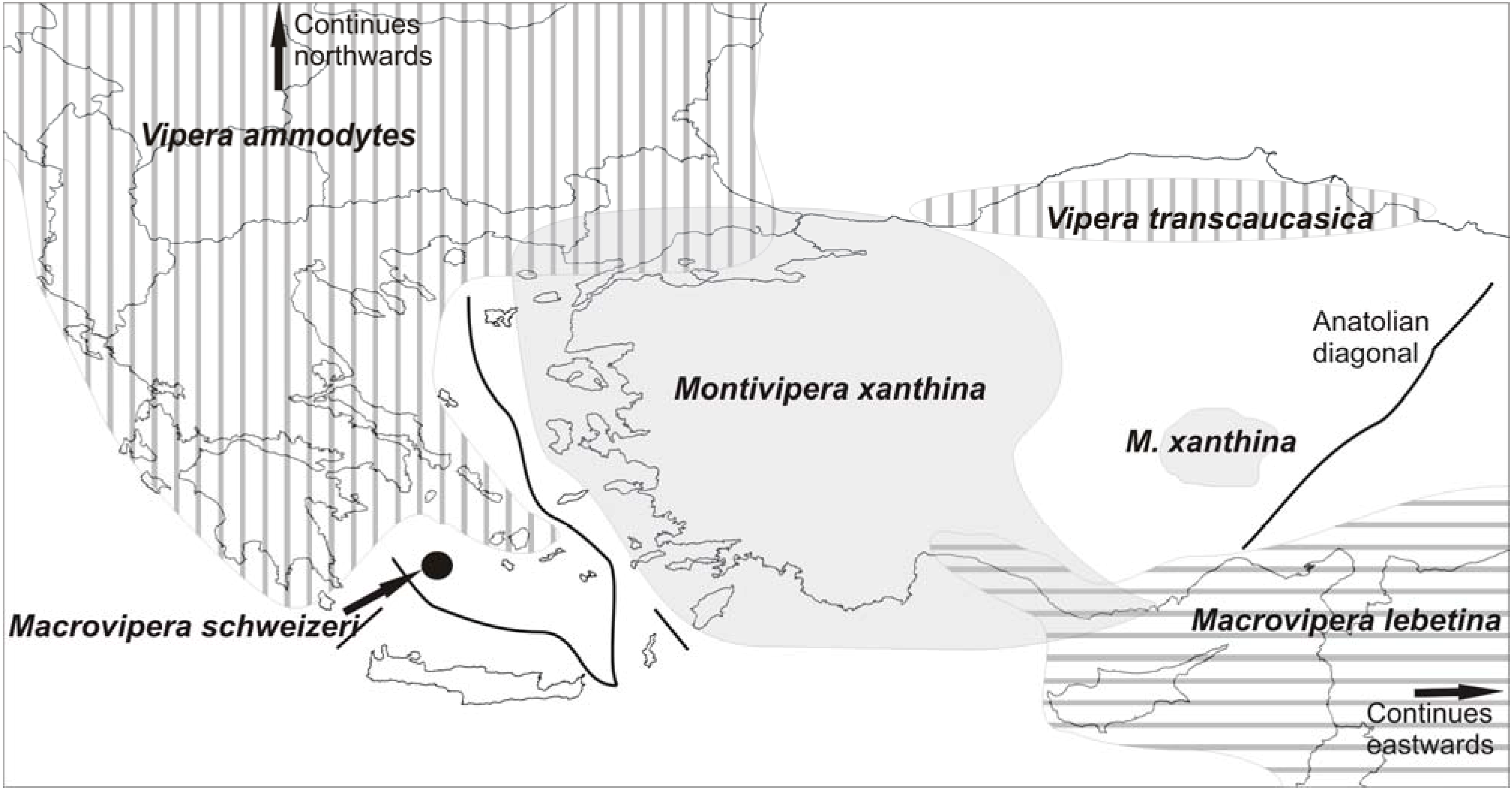
Macrovipera is currently considered to contain two species:
M. schweizeri and
M. lebetina. These have split during the Messinian crisis ca 5MYA [
40,
43], leaving today a restricted population on the Aegean island group of Milos (
M. schweizeri) and
M. lebetina with a wide distribution to the east of the Anatolian Diagonal [
44] in Turkey, with a wide gap void of the genus in Greek islands and western Turkey (
Figure 10). This void may have left space for (or may have in part been caused by) the invasion and further differentiation of
V. ammodytes in the area. Moreover, a third species is distributed in western Turkey and in particular west of the Anatolian Diagonal reaching Eastern Greece:
Montivipera xanthina [
44]. The authors suggest a vicariant differentiation of
Montivipera beginning in the early Plioecene to the widespread
M. xanthina and four more species with restricted distributions along the mountains forming the Anatolian diagonal. Once more we may see this event coupled with the earlier splitting of
Macrovipera and to its extinction from east of Milos until the Anatolian diagonal.
Figure 10.
Approximate distributions of certain vipers in the Aegean area. Not mentioned in the text but shown here is V. transcaucasica, until recently considered a subspecies of V. ammodytes. Another isolated population of the species (not shown) occurs east of the Anatolian diagonal. A vicariant event and/or competition with M. xanthina may have been responsible for the gap in V. ammodytes distribution. Note that despite their long history in the area, vipers respect the MAT as well as the Anatolian Diagonal and they appear to have never reached the southern Aegean.
Figure 10.
Approximate distributions of certain vipers in the Aegean area. Not mentioned in the text but shown here is V. transcaucasica, until recently considered a subspecies of V. ammodytes. Another isolated population of the species (not shown) occurs east of the Anatolian diagonal. A vicariant event and/or competition with M. xanthina may have been responsible for the gap in V. ammodytes distribution. Note that despite their long history in the area, vipers respect the MAT as well as the Anatolian Diagonal and they appear to have never reached the southern Aegean.
A case of another snake,
Malpolon insignitus [
45] is one of impressive long distance dispersal. Ancestral populations of the species in the Maghreb (NW Africa) dispersed both westwards and eastwards during late Miocene. The east–moving clade traveled through northern Africa and from there continued to the east and north reaching the Caspian Sea to the north-east and the Dalmatian coast to the north-west. The later clade invaded Greece, but did not make it to the Cyclades or Crete.
A unique pattern is observed in a freshwater terrapin,
Mauremys rivulata. Mantziou and coworkers [
46] compared partial mt DNA sequences of animals from Greece, Cyprus, and Jordan finding as little as 0.7% mean genetic difference with no geographic structure. The terrapin appears to have dispersed erratically. This pattern may be explained since the species abounds in fresh and brackish waters and at least for short periods is tolerant of salt water.
3. Discussion
3.1. Sources of Diversity
The sheer amount of diversity studied renders the separation of the effect of each of the aforementioned phenomena difficult to account for. On the other hand “given the plethora of species it is easy to find distributional patterns which may fit the craziest hypothesis” [
47]. The same author however notes that one should not close the eyes before patterns which have not been expected and are repeatedly found. Before approaching the herpetofaunal diversity addressed here we consider that we should recapitulate the diversity of factors which influence it.
Abiotic diversity
Geological events in the area account for the diversity in geomorphological features as well as for various substrates (sedimentary, volcanic, metamorphic or combinations of the three). Both factors, but mainly geomorphology, influence geographic diversity by hindering or bringing in contact faunal elements from different origins. At the same time geological events have impact on the climate. The above change in unpredictable combinations with time.
Biological diversity A: non human induced
Biological processes generate diversity, which is indispensable for providing material for natural selection to process. The means of producing this diversity are interactions among the faunal elements as well as between the later and the environment both submitting to its changes and causing it to change [
48]. Diversity increases in a positive feedback loop [
49,
50]. In the words of Benton and Emerson [
49], “Ironically, high species diversity may lead to more speciation, not, as had been assumed, less: more species create more opportunities and selective pressures for other species to respond to, rather than capping diversity at a fixed equilibrium level.”
A more specific point regarding the area in question lays in its role as a refugium in several instances during the quaternary, where the genetic information already present is redistributed, increasing diversity [
22,
51].
The meeting of faunal elements of different origins may initially have a negative impact on overall diversity by mutual exclusion of species with the same or similar niches. However, in the long term they will follow the flow of the positive feedback described, increasing diversity.
Biological diversity B: human induced
Humans act upon biological diversity in practically all conceivable manners: voluntary and involuntary introductions as well as extinctions of species (e.g., hunting, poisoning, polluting, clearing forests for cultivation or drying up wetlands), which may occur in various scales (local to global), and overall, lead to homogenization processes (see also [
52] and references therein). Even though there are indications showing that human practices in the area have covered the entire spectrum described, it is difficult, if at all possible, to assess in details their impact on the herpetofauna.
The interaction of the described sources of diversity has resulted in the high biodiversity we record today.
3.2. Three Basic Patterns, an Outlier and Exceptions
The “Ancients”
The first regards animals present in the area before the opening of the Mid Aegean Trench (MAT). Four of the described cases (
Samalamandra/Mertensiella-Lyciasalamandra),
Ophiomorus,
C. kotschyi,
N. tessellata) may be included in this group. It is highly probable that two more groups,
Pelophylax and
Bufo may be as well included in this category, but this hypothesis may not be supported until more data become available. Another uncertain case is the one of
Macrovipera, which has been assumed to have split during the Messinian crisis [
43]. However,
M. schweizeri shares the Milos island group with
Podarcis milensis which has been suggested to have split from mainland populations
circa 8 MYA [
4], a time compatible with the opening of the MAT. The opening of the MAT resulted in a vicariant splitting of populations, which may or may not be evident today. It is still obvious in the distance between
Ophiomorus populations from Peloponnesos and SW Turkey and the couple
Salamandra/Lyciasalamandra. In the case of the dice snake, events which occurred in later instances dominate present day distributions possibly blurring older effects.
All cases presented regard Palaearctic taxa. African animals have not yet come in to play.
Post MAT
The second basic pattern regards animals which arrived in the area after the formation of the MAT. Podarcis, Ablepharus, Ophisops, Malpolon as well as possibly Bufo and Pelophylax.
Beginning by the “ancients”, Ophiomorus remains restricted on the two sides of the MAT. Lyciasalamandra does not cross the MAT westwards even though it remained on Kasos/Karpathos islands. Asian and European species of Salamandra remained in their respective continents.
The anurans Pelophylax spp., Bufo spp. and Hyla spp., which evolved West and East of the MAT never crossed it, even though dispersal events occurred and in some cases brought distinct lineages into contact but only if the MAT was circumvented from the north.
Similarly, all taxa which arrived after the forming of the MAT appear bound by it. Ophisops, Podarcis, Ablepharus, Anatololacerta, Malpolon, Vipera and Montivipera are taxa of various origins (European, Asian, African) which fall in this case (but see paragraph on exceptions below). However, bounded by the MAT does not mean uniformity in pattern. Each taxon, restricted by its niche’s requirements and time, reacts differently to the same challenge. This is obvious comparing the cases of the three Asian lizards (Ablepharus, Anatololacerta, Ophisops).
The newcomers
The third category regards recent arrivals via human induced (voluntary or not) introductions. These animals (
Tarentola,
Hemidactylus,
Chalcides,
H. viridiflavus,
Chamaeleo africanus) originate from all three continents and their distributions in the Aegean are random. Only as an observation, not yet deserving the “status” of a pattern, we notice that these species are found only in the southern part of the Aegean except for the anthropophilic
H. turcicus, which marginally occupies coastal areas of the N Aegean. There may be an explanation for this observation, namely the turbulent geological history of the area, which resulted in severe fragmentation of landmasses and/or long periods of isolation (e.g., the island of Crete) which possibly decimated extant faunas. Crete in particular hosts all three lizard species of this category and has a recorded fossil herpetofauna, which includes animals presently not found on the island such as
Discoglossus sp.,
Testudo sp., and
Agama sp. [
53]. Extant herpetofauna on Crete is considered as poor (14 species compared to more than 20 on the much smaller island of Rhodes) rendering it susceptible to host newcomers. Actually three alien taxa have arrived within the past decades (
Testudo spp.,
Trachemys scripta,
Lithobates catesbeianus), the two latter having established viable populations. Finally, the randomness mentioned, reflects an amalgam of ecological requirements with specific commercial routes and destinations. Answers to this problem may be sought in a historical/archaeological rather than a biological quest.
The outlier
The last category includes a single animal,
Mauremys rivulata, for which the MAT is practically non-existent. This is an indirect indication of the efficiency of sea as a barrier for land taxa, whereas a reptile not strictly bounded by sea water barriers apparently ignores it. It is worth noting the difference of the pattern exhibited by
M. rivulata with the one of another terrapin,
Emys orbicularis. The phylogenetic relationships of the latter appear geographically structured with higher diversity in south areas of its distribution [
54]. Unlike
Mauremys, which is commonly found in marginal and often temporary wetlands,
Emys (although tolerant of brackish waters) prefers large permanent water bodies. This point is supported by the difference in the two species’ distributions:
Mauremys is “restricted” to coastal east Mediterranean areas whereas
Emys is widely distributed throughout Europe, excluding small islands and even some big ones as Crete. In other words, being a terrapin is not sufficient to overlook the sea as a barrier. The pattern of
Emys is more similar to the case of
Pelophylax than to its more closely related
Mauremys.
Exceptions
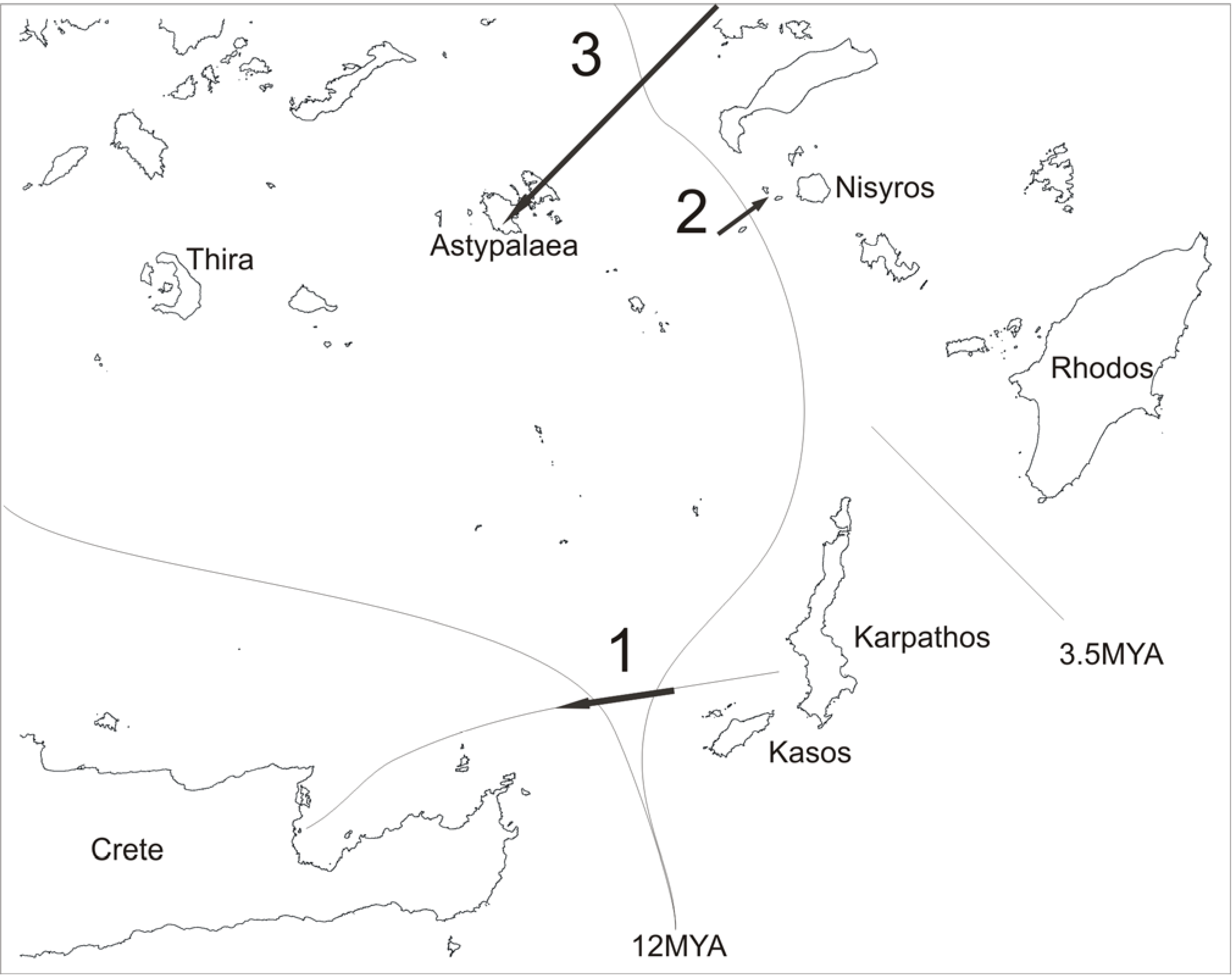
There are three well documented cases of species that have crossed the MAT by dispersal, most probably not human induced (
Figure 11).
Figure 11.
Three cases of documented MAT crossings. 1.
Ablepharus crossed from the Kasos−Karpathos complex to a tiny (4.4 ha) islet in the bay of Ag. Nikolaos, Crete. Although an explanation involving rafting appears to be most probable, it is strange how the rafts “missed” Crete or other larger islets. It is equally difficult to explain how another islet in the same bay with an area of 35 ha, less than 50m from the first, , has not been invaded. 2.
Podarcis erhardii crossed the MAT towards a couple of volcanic islets west of Nisyros, most probably from an equally small nearby islet west of the trench. 3. One of the few remaining small wetlands in the Cyclades is located on the south part of Astypalaea. It is inhabited by a population of
Pelophylax bedriagae with phylogenetic affiliation to populations found in the northern Aegean and northern Turkey [
21] and not as one intuitively would have thought from nearby central-western Turkey or Rhodos.
Figure 11.
Three cases of documented MAT crossings. 1.
Ablepharus crossed from the Kasos−Karpathos complex to a tiny (4.4 ha) islet in the bay of Ag. Nikolaos, Crete. Although an explanation involving rafting appears to be most probable, it is strange how the rafts “missed” Crete or other larger islets. It is equally difficult to explain how another islet in the same bay with an area of 35 ha, less than 50m from the first, , has not been invaded. 2.
Podarcis erhardii crossed the MAT towards a couple of volcanic islets west of Nisyros, most probably from an equally small nearby islet west of the trench. 3. One of the few remaining small wetlands in the Cyclades is located on the south part of Astypalaea. It is inhabited by a population of
Pelophylax bedriagae with phylogenetic affiliation to populations found in the northern Aegean and northern Turkey [
21] and not as one intuitively would have thought from nearby central-western Turkey or Rhodos.
All three successful crossings of the MAT have been realized in the southern Aegean. In cases 1 and 2 the host-islands had a single lizard (C. kotschyi) prior to the newcomers. Astypalaea’s only Anuran inhabitant is P. bedriagae. The islands, being void from potential competitors, renders them susceptible to successful invasions. This point is most obvious in the second case described. The two potential invaders do not coexist on any island: Podarcis erhardii is largely distributed west of the MAT and Ophisops elegans on practically all eastern Aegean islands. It was only a matter of luck that Podarcis made it first to the two small islets. In any case, we consider these cases as true exceptions regarding single crossings of the MAT as opposed to the erratic dispersal of M. rivulata an animal less hindered by sea.
One more point worth noting from the examples of exceptions is the effect that random events may have a significant impact, especially for smaller islands and islets. Islands and islets may have several species or their entire faunas wiped out by extreme conditions (e.g., storms or tsunamis); the forming of the new ecosystem is often defined by who makes it there first (see also [
55]).
3.3. What about Other Organisms?
In all cases studied with one exception (see below), animals and plants appear to follow the patterns described above. Specifically, land snails of the genus
Mastus [
56] scorpions of the genus
Mesobuthus [
57] terrestrial isopoda of the genera
Ligidium [
58] and
Orthometopon [
59] cave crickets of the genus
Dolichopoda [
60] as well as
Nigella arvensis of the plant family Ranunculaceae [
61] are shown to have populations geographically structured and differentiated in concordance with the barriers discussed, falling into one of the three basic patterns described (see also [
62]).
The exception mentioned is described in the work of Papadopoulou and coworkers [
63]. One of the points made regard the darkling beetle
Eutagenia smyrnensis. Phylogenetic analyses of Aegean populations of this species produce two distinct mt DNA lineages. The latter suggests need for taxonomic revision, hence the authors refer to the species as the “species complex
E. smyrnensis sensu lato”. What is interesting is that the two lineages correspond to animals with clearly distinct ecological preferences: one group includes animals found on stable soil and the other to animals found on sandy beaches. Going one step further into the two phylogenetic clades, the stable surface group appears geographically structured, and similarly to the other cases discussed, restricted by the barriers shown in
Figure 3. The second (sand dwelling) clade, however, has a feeble geographical structure where populations east and west of the MAT appear together in some subclades. The authors suggest that dispersal rates and hence gene flow among populations are lower in the more stable compared to the less stable habitats. Therefore, sand-inhabiting darkling beetles are subject to selection for a higher migration rate.
From the patterns proposed, the case sand of dwelling Eutagenia is similar to the ones regarding animals which cross the MAT, namely M. rivulata (random crossings) and the “exceptions” (single crossings). Despite being a terrestrial animal, sand dwelling Eutagenia’s pattern resembles the one of M. rivulata i.e., not bounded by sea.
3.4. Vicariance vs. Dispersal
In the cases discussed, one may find typical examples of vicariance (e.g., Ophiomorus) or dispersal (e.g., Malpolon) as the causal factor for the distribution observed. However, in most cases both factors come in play. We consider that the extent that the one or the other will dominate is not genetically or otherwise inherent but largely dependent on the physical and biological background of the time it occurs.
3.5. Pre vs. Post Quaternary Differentiation and Quaternary Refugia
In reality this is not a true dilemma. Organisms “carry” their history, which in some cases may be reconstructed. In several cases discussed we may discern pre-Quaternary events, most prominent of which is the Messinian crisis, as the time of differentiation of several taxa recognized today at the species level. However we do not treat this simplistically as a case of “biogeographical provensialism” (see discussion in [
64]). We usually regard the Messinian crisis as a phenomenon which desiccated the Mediterranean and associate it to tectonics. The events during this period were much more complex involving successive flooding and desiccation as well as climatic changes [
2,
65]. The triggering of speciation via joining of landmasses or vicariant events may have been enhanced by the coupling with these phenomena.
However, contemporary distributions have largely been shaped during the Quaternary, and more specifically, may be attributed to the climatic oscillations that occurred throughout its duration [
66,
67]. The changes in distributions due to climate changes have also caused, or at least triggered, speciation events, as well through repeated allopatry [
25] after periods of redistribution of the genetic information already present among populations in refugia [
22,
51]. These speciation events are at various phases
i.e., from clear-cut, to incipient species and finally to marginal populations which have initiated divergence (morphological and/or molecular) from their “maternal” one. A similar situation is found in central European taxa [
22].
Many authors e.g., [
25,
51] as well as the cases presented, warn against a simplistic consideration of refugia. Their locations are determined by complex historical and environmental factors, the cumulative effects of which need to be considered because they have occurred since the Tertiary, rather than solely during the last glacial period [
68]. Moreover, we should keep an open mind about the role of refugia apart from on the Mediterranean peninsulas or near the sea as discussed in e.g., [
25,
69] and suggested in other works [
4,
21]. Additionally, proposed refugia should be critically and case-by-case assessed as what is a refuge for one species is not necessarily for another. Finally, however scarce [
37], the fossil evidence is an irreplaceable failsafe for controlling proposed phylogenetic histories.
3.6. General Trends
The main distribution patterns in the Aegean are related to geological time. The “ancients” group consists only of Palaearctic animals, whereas in the other cases African animals are involved as well. Nevertheless, the general patterns are general insofar as we observe them at a broad scale. Each group studied has its peculiarities in what regards the routes followed, refugia (and microrefugia) they originated from, mechanisms involved (
i.e., vicariant, dispersal) and degree of differentiation. This view appears consistent with the perspective given by Benton [
70]
i.e., predominance of abiotic factors at large temporal and geographical scales whereas at smaller scales biotic interactions may be more important.
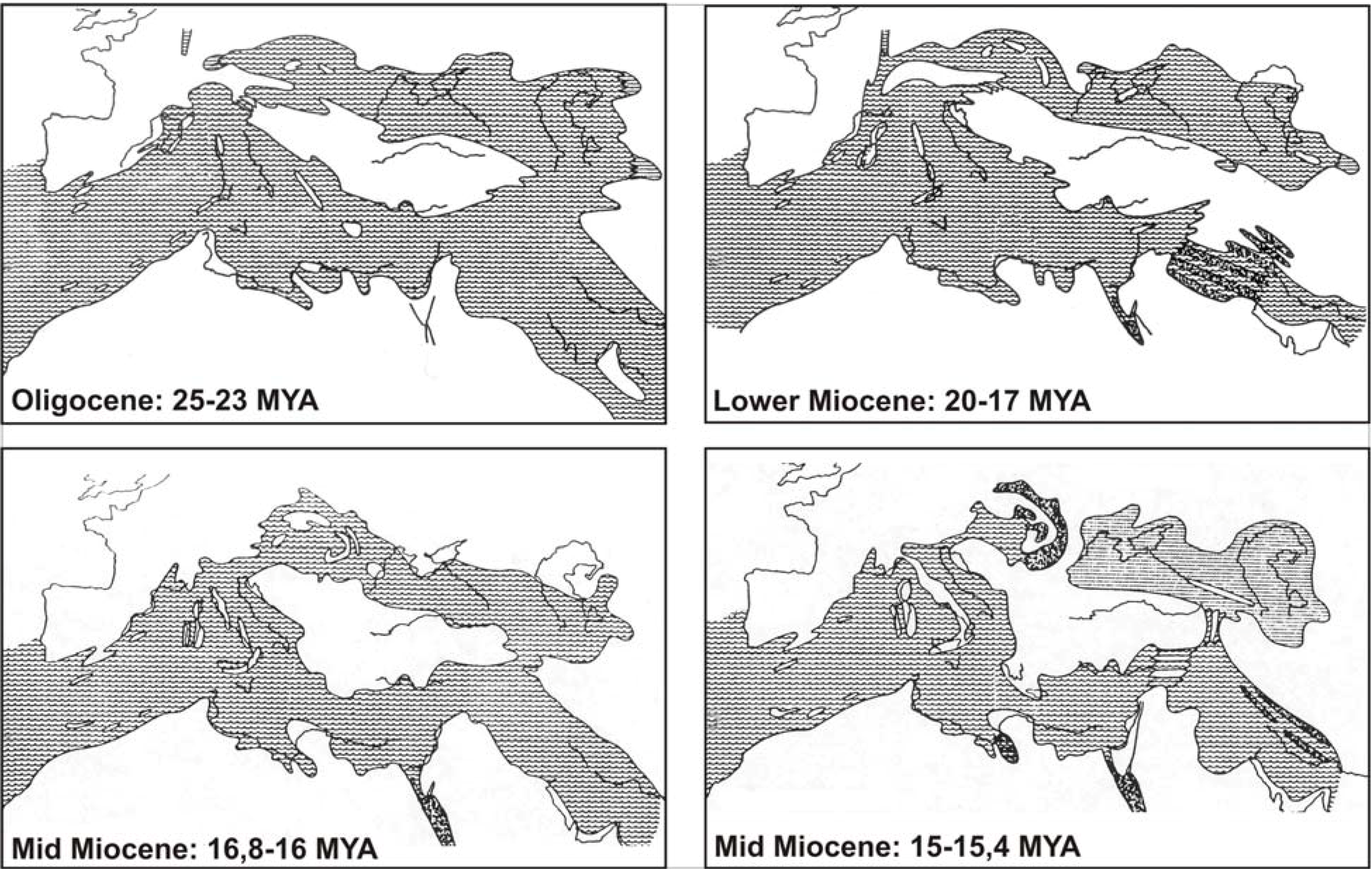
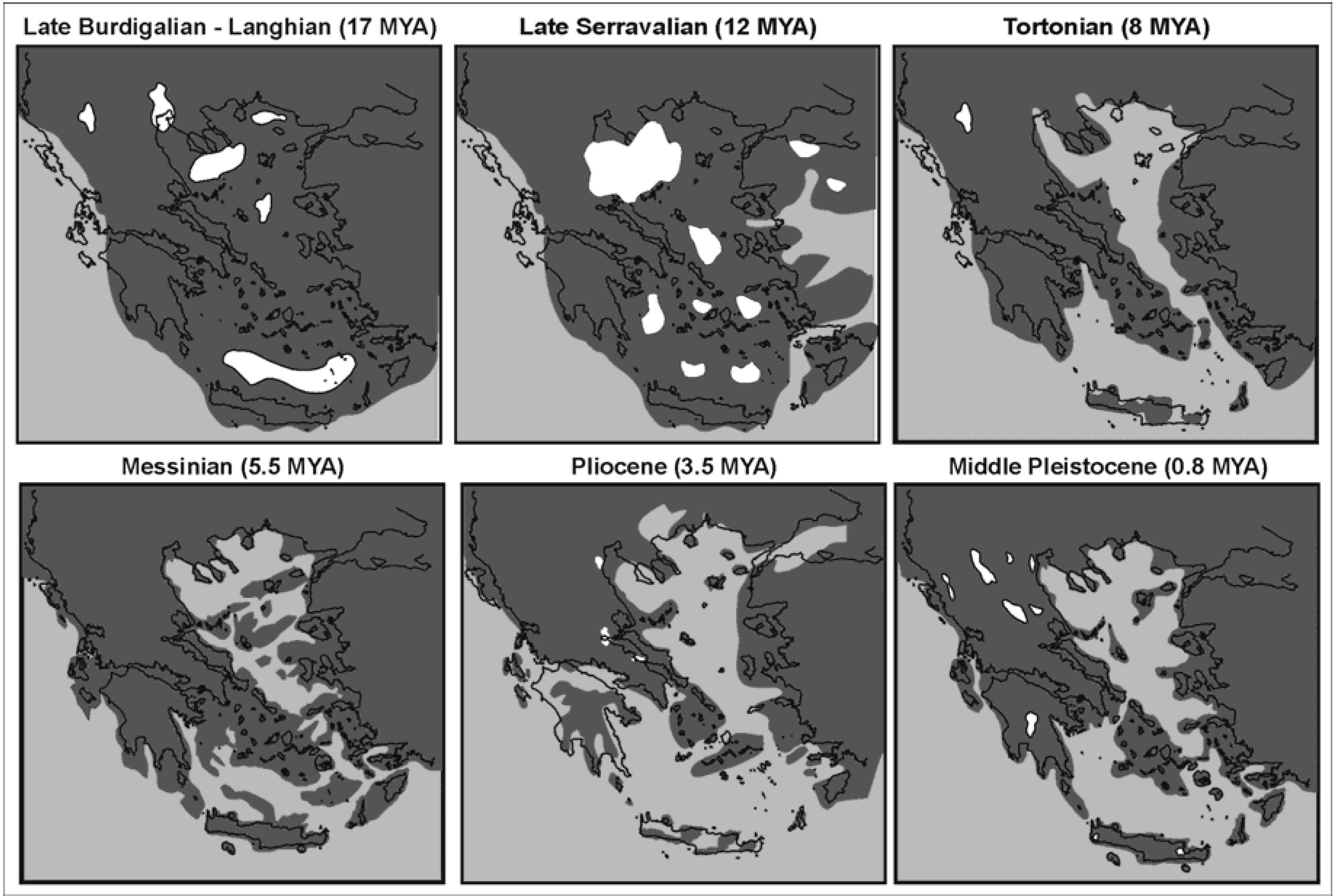
Key dates appear to be the initiation of the MAT split (12 MYA), as well as its ending (9 MYA), the Messinian crisis (5 MYA), climate oscillations during the Pleistocene (where however the last glacial maximum has been disproportionally better investigated) and finally the Holocene when modern humans came into the picture, causing massive ecosystem alterations, voluntary and involuntary animal introductions.
Pre-Quaternary geomorphological rearrangements at the respective times (MAT, separation of Crete) efficiently blocked dispersal in most cases of terrestrial herpetofauna. During the Quaternary, climate alterations caused consecutive distributional rearrangements, but species still remained within the boundaries created earlier by tectonics. The efficiency of the barriers may have been enhanced by the differentiation and organization of local faunal communities in the “host” areas. Some species, however, when carried by humans much later did not appear to have problems with the crossing of local or much larger barriers (e.g., from Africa to Europe). Last but not least, stochastic events may have a significant impact especially on smaller islands and islets. A general view on Aegean’s biodiversity, largely consistent with the major points made here is supported in [
62].
3.7. Diversity
The backbone of the present work is nothing else than a list of taxa. This first level of diversity (taxonomic) may be unfolded and expanded in several dimensions revealing sources of diversity and interactions among them: Diversity in geological processes, geomorphology, faunal origins, climate, biological assemblages including their interactions and a special case of the latter, human activities.
We approach all the above parameters and many more in the search of causal factors that may explain the taxonomic diversity. Our quest focuses on which evolutionary mechanisms are affected by each of the factors mentioned above. When discussing here for example vicariance versus dispersal, or pre versus post quaternary differentiation, we invoke different evolutionary mechanisms to account for the diversity observed. But we have not directly discussed the relation of evolution and diversity.
Even though rarely stated explicitly, the prevailing view on the relation between evolution and diversity is that “Evolution begets Diversity”. But could it be the other way around? After all, there is no evolution without diversity. Is it legitimate to say that diversity begets evolution? Or could it be that both statements are true, simultaneously bonding Evolution and Diversity in a dialectical relationship? And if this is true, how would it alter our approaches in biology? For example, how would a conservationist define biodiversity? Or on a completely different line, how would it affect the study of artificial intelligence or other complex (=diverse?) systems such as economy? Moreover, what would be the impact on broader biological manifestations, e.g., science, art or politics? This is not a paper on philosophy, and in any case is not here to deal with such questions. However, intrigued by the biodiversity described, we wondered about Diversity as a broader quality, in line with the very need that led to a dedicated publication [
52]. We include this last paragraph to highlight the lack of theoretical exploration of this notion, which we believe is important to address.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










