A Molecular Survey of the Diversity of Microbial Communities in Different Amazonian Agricultural Model Systems


Abstract
:1. Introduction
2. Material and Methods
2.1. Model I: The “Slash-and-Burn” System in Western Amazonia
2.1.1. Study sites and soil sampling
2.1.2. DNA extraction and ARISA fingerprinting
2.1.3. ARISA Data Analysis
2.2. Model II: The Amazonian Dark Earth and Sustainable Agriculture in the Central Amazon
2.2.1. Soil sample collection
2.2.2. DNA Extraction and PCR amplification to T-RFLP analysis
2.2.3. Pyrosequencing of the 16S rRNA gene isolated from ADE and black carbon
3. Results and Discussion
3.1. Model I of Amazonian Agricultural System: The “Slash-and-Burn” System in Western Amazonia
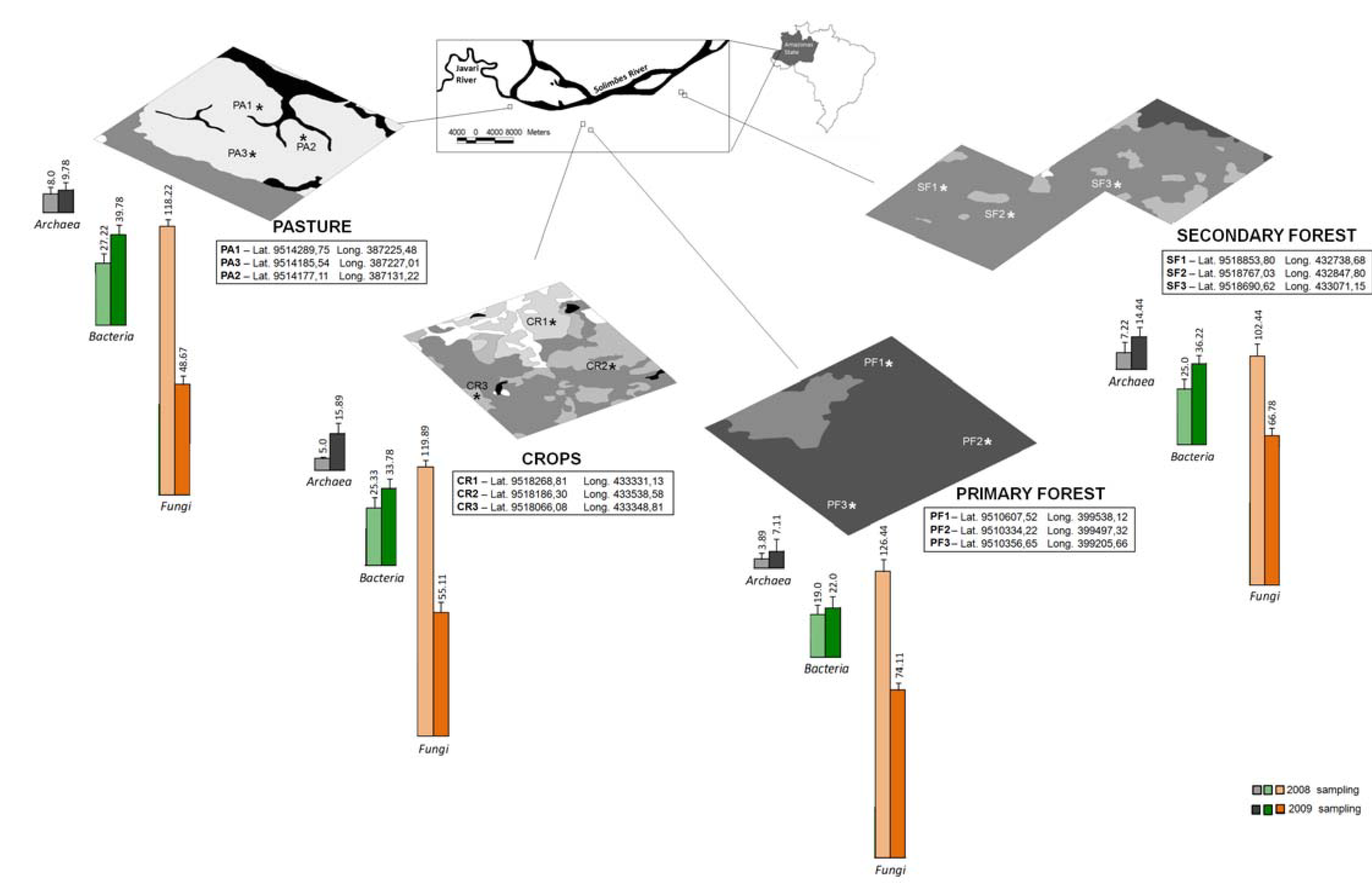
 ), secondary forest (
), secondary forest (  ), crops (
), crops (  ), pasture (
), pasture (  ), water (
), water (  ) and exposed soil (
) and exposed soil (  ). Soil samples were collected at different points (*), at a depth of 0–20 cm from sites representing primary tropical rain forest, traditional crops systems of the indigenous people, pasture and secondary forest over a two-year sampling (March 2008 and January 2009). The richness values were calculated based on triplicate PCR products from the soil samples.
), secondary forest ( ), crops ( ), pasture ( ), water ( ) and exposed soil ( ). Soil samples were collected at different points (*), at a depth of 0–20 cm from sites representing primary tropical rain forest, traditional crops systems of the indigenous people, pasture and secondary forest over a two-year sampling (March 2008 and January 2009). The richness values were calculated based on triplicate PCR products from the soil samples.
). Soil samples were collected at different points (*), at a depth of 0–20 cm from sites representing primary tropical rain forest, traditional crops systems of the indigenous people, pasture and secondary forest over a two-year sampling (March 2008 and January 2009). The richness values were calculated based on triplicate PCR products from the soil samples.
), secondary forest ( ), crops ( ), pasture ( ), water ( ) and exposed soil ( ). Soil samples were collected at different points (*), at a depth of 0–20 cm from sites representing primary tropical rain forest, traditional crops systems of the indigenous people, pasture and secondary forest over a two-year sampling (March 2008 and January 2009). The richness values were calculated based on triplicate PCR products from the soil samples.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attributes | Land Use | |||||||
|---|---|---|---|---|---|---|---|---|
| Primary forest* | Secondary forest* | Crops* | Pasture* | |||||
| 2008 | 2009 | 2008 | 2009 | 2008 | 2009 | 2008 | 2009 | |
| Chemical pH (CaCl2) | 4.0 ± 0.4 | 4.1 ± 0.2 | 3.9 ± 0.05 | 4.0 ± 0.3 | 4.8 ± 0.8 | 4.4 ± 0.6 | 4.5 ± 0.6 | 3.88 ± 0.1 |
| OM (g.dm-3) | 29.0 ± 14.7 | 32.3 ± 5.2 | 34.7 ± 20.3 | 26.3 ± 3.9 | 39.3 ± 15.5 | 25.7 ± 2.9 | 27.3 ± 9.2 | 31.3 ± 5.5 |
| Fe (mg.dm-3) | 159.7 ± 17.1 | 93 ± 8.2 | 109.3 ± 30.1 | 69.7 ± 18.6 | 55.0 ± 10.8 | 74.7 ± 13.7 | 66.3 ± 13.0 | 147.3 ± 21.2 |
| P (mg.dm-3) | 9.7 ± 1.1 | 11.0 ± 2.0 | 8.0 ± 1.4 | 6.0 ± 1.0 | 1.9 ± 0.8 | 9.3 ± 1.1 | 6.0 ± 0.8 | 6.7 ± 0.7 |
| K (mmolc.dm-3) | 1.4 ± 0.5 | 1.3 ± 0.1 | 1.7 ± 0.5 | 1.2 ± 0.5 | 1.9 ± 0.9 | 2.1 ± 0.8 | 2.1 ± 0.3 | 1.6 ± 0.9 |
| Physical Clay (%) | 31.0 ± 7.6 | 35.7 ± 6.1 | 35.5 ± 12.7 | 36.0 ± 12.3 | 25.8 ± 2.6 | 22.3 ± 7.5 | 25.4 ± 6.3 | 27.7 ± 9.4 |
| Silt (%) | 51.7 ± 1.9 | 47.7 ± 3.4 | 45.8 ± 6.5 | 43.7 ± 3.2 | 53.6 ± 7.3 | 41.3 ± 2.5 | 36.6 ± 11.0 | 37.7 ± 1.8 |
| Sand (%) | 17.3 ± 5.7 | 16.7 ± 4.9 | 18.7 ± 13.4 | 20.3 ± 9.5 | 20.6 ± 4.7 | 36.3 ± 3.1 | 38.0 ± 16.6 | 34.7 ± 2.2 |
 ); Secondary Forest (SF,
); Secondary Forest (SF,  ); Crops (CR,
); Crops (CR,  ); Pasture (PA,
); Pasture (PA,  ). Each point in the ordination is identified by the abbreviation of the correspondent land use system followed by the point sampling (1, 2 and 3) and replicate PCR product analyzed (A, B and C).
); Secondary Forest (SF, ); Crops (CR, ); Pasture (PA, ). Each point in the ordination is identified by the abbreviation of the correspondent land use system followed by the point sampling (1, 2 and 3) and replicate PCR product analyzed (A, B and C).
). Each point in the ordination is identified by the abbreviation of the correspondent land use system followed by the point sampling (1, 2 and 3) and replicate PCR product analyzed (A, B and C).
); Secondary Forest (SF, ); Crops (CR, ); Pasture (PA, ). Each point in the ordination is identified by the abbreviation of the correspondent land use system followed by the point sampling (1, 2 and 3) and replicate PCR product analyzed (A, B and C).
| NMDS | ||||||||||
| Stressa | Correlationb(%) | Variabilityc(%) | Stressa | Correlationb(%) | Variabilityc(%) | Interactionsd | ANOSIM | |||
| 2008 | 2009 | 2008 | 2009 | |||||||
| CR,PF | 0.275 | 0.881 | ||||||||
| Archaea | 0.021 | 62.8 | 43.0 | 0.031 | 61.1 | 63.5 | PA,PF | 0.725 | 0.616 | |
| SF,PF | 0.300 | 0.152 | ||||||||
| CR,PF | 0.448 | 0.642 | ||||||||
| Bacteria | 0.028 | 83.7 | 75.5 | 0.022 | 58.5 | 58.5 | PA,PF | 0.985 | 0.701 | |
| SF,PF | 0.270 | 0.601 | ||||||||
| CR,PF | 0.221 | 0.762 | ||||||||
| Fungi | 0.039 | 63.9 | 75.5 | 0.019 | 54.7 | 83.7 | PA,PF | 0.057 | 0.983 | |
| SF,PF | 0.218 | 0.579 | ||||||||
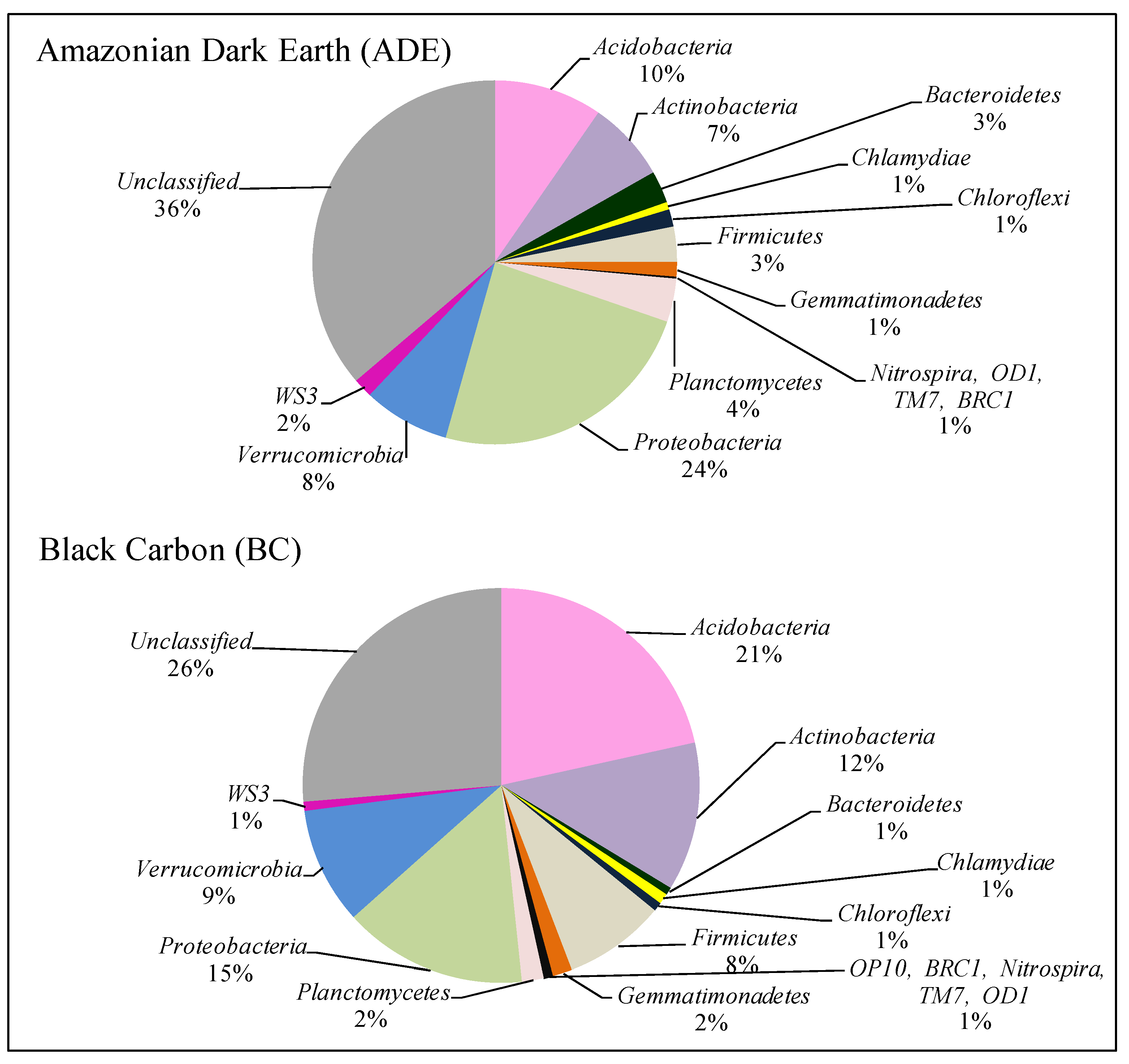
3.2. Model II of an Amazonian Agricultural System: Bacterial Community Structure in Amazonian Dark Earth (ADE) and Black Carbon (BC)
| Attributes | Hatahara | |
| ADE | ADJ | |
| pH (CaCl2) | 5.2 ± 0.2a | 3.7 ± 0.11b |
| OM (g.dm-3) | 53 ± 2.64a | 33.3 ± 1.15b |
| P (mg.dm-3) | 508.7 ± 45.5a | 7.3 ± 1.15b |
| S (mg.dm-3) | 4.3 ± 0.57a | 3.6 ± 0.57b |
| K (mmolc.dm-3) | 1.1 ± 0.25a | 0.6 ± 0.05b |
| Ca (mmolc.dm-3) | 138.3 ± 7.63a | 6.3 ± 1.15b |
| Mg (mmolc.dm-3) | 13.7 ± 3.05a | 1 ± 0b |
| Al (mmolc.dm-3) | 0 ± 0a | 14 ± 1.73b |
| H+Al (mmolc.dm-3) | 48.7 ± 5.77a | 95.7 ± 14.6b |
| SB (mmolc.dm-3) | 153.7 ± 5.39a | 8 ± 1.12b |
| CEC (mmolc.dm-3) | 201.7 ± 2.25a | 103 ± 615.7b |
| BS (%) | 76 ± 2.64a | 7.7 ± 0.57b |
 ) and ADJ (
) and ADJ (  ) soil and BC (
) soil and BC (  ) based on the structure of soil bacterial community as determined by terminal restriction fragment length polymorphism (T-RFLP) and soil attributes. The ordinations were performed using data obtained with the enzymes MspI and HhaI.
) and ADJ ( ) soil and BC ( ) based on the structure of soil bacterial community as determined by terminal restriction fragment length polymorphism (T-RFLP) and soil attributes. The ordinations were performed using data obtained with the enzymes MspI and HhaI.
) based on the structure of soil bacterial community as determined by terminal restriction fragment length polymorphism (T-RFLP) and soil attributes. The ordinations were performed using data obtained with the enzymes MspI and HhaI.
) and ADJ ( ) soil and BC ( ) based on the structure of soil bacterial community as determined by terminal restriction fragment length polymorphism (T-RFLP) and soil attributes. The ordinations were performed using data obtained with the enzymes MspI and HhaI.
| Samples | T-RFs Richness | Evenness | H´(loge) | |||
| MspI | HhaI | MspI | HhaI | MspI | HhaI | |
| ADE | 100 ± 15.7 | 10.3 ± 5 | 0.90 ± 0.01 | 0.80 ± 0.09 | 4.15 ± 0.09 | 1.84 ± 0.64 |
| ADJ | 42.6 ± 20.5 | 11.6 ± 3 | 0.84 ± 0.02 | 0.85 ± 0.06 | 3.10 ± 0.52 | 2.09 ± 0.31 |
| BC | 56.3 ± 9.8 | 20.3 ± 24 | 0.82 ± 0.08 | 0.81 ± 0.09 | 3.33 ± 0.39 | 1.99 ± 0.68 |
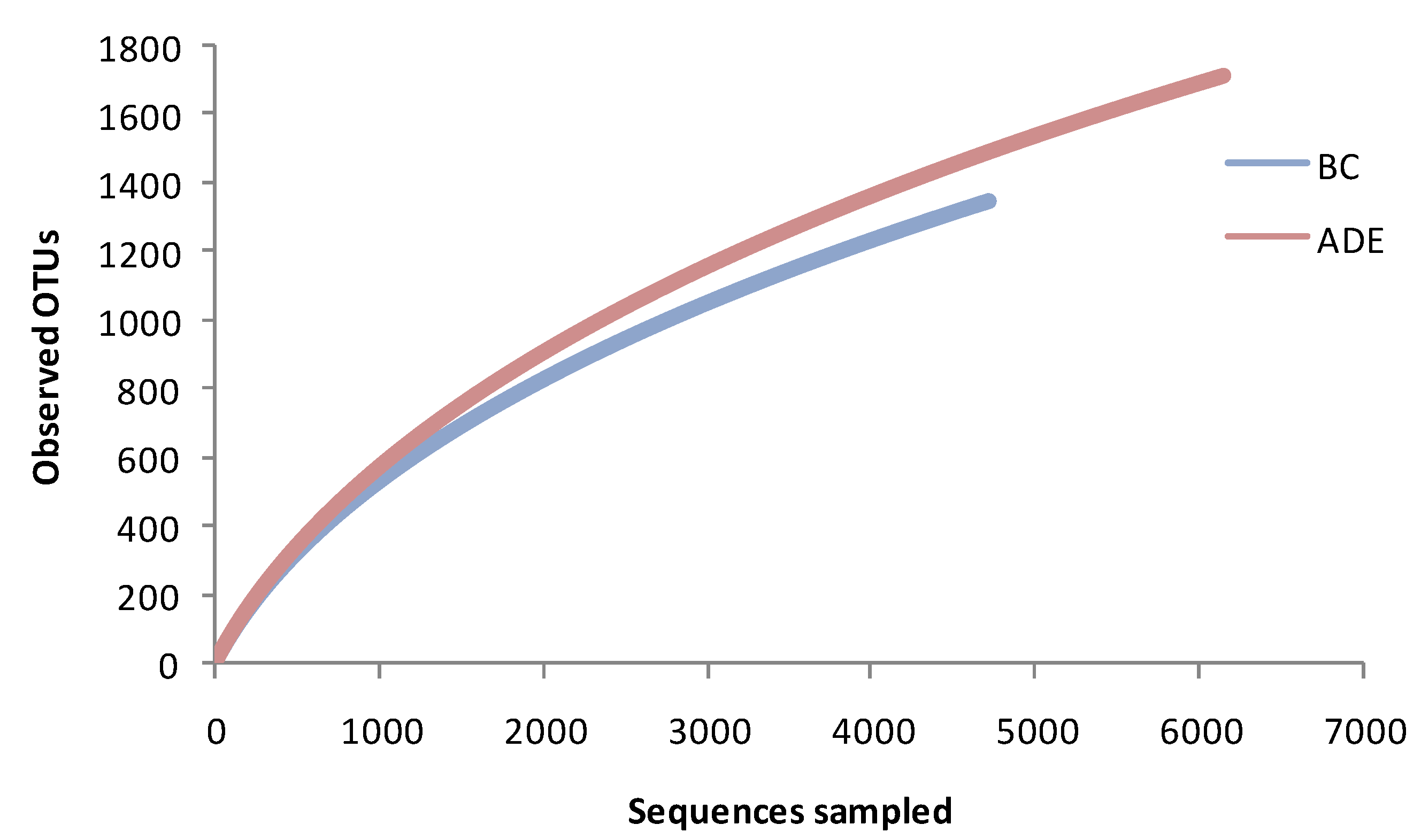
3.2.1. The bacterial OTU richness of ADE and BC



4. Conclusions
Acknowledgements
References
- Fearnside, P.M. Uso da terra na Amazônia e as mudanças climáticas globais. Braz. J. Ecol. 2006, 7, 83–100. [Google Scholar]
- Nye, P.H.; Greenland, D.J. The soils under shifting cultivation. Commonwealth Bureau Soils. In Tech. Comm.; Harpenden, UK, 1960; No. 51. [Google Scholar]
- Sanchez, P.A; Buol, S.W. Soils in the tropics and world food crisis. Science 1975, 188, 598–603. [Google Scholar]
- Barrow, C.J. Land Degradation; Cambridge University Press: Cambridge/London/New York, 1991. [Google Scholar]
- Juo, A.S.; Manu, R.A. Chemical dynamics in slash-and-burn agriculture. Agricult. Ecosys. Environ. 1996, 58, 49–60. [Google Scholar] [CrossRef]
- Fidalgo, E.C.C.; Coelho, M.R.; Araujo, F.O.; Moreira, F.M.S.; Santos, H.G.; Brefin, M.L.M.S.; Huising, J. Levantamento do uso e cobertura da terra de seis áreas amostrais relacionadas ao projeto BiosBrasil (Conservation and Sustainable Management of Below-Groud Biodiversity: Phase I). In Município de Benjamin Constant (AM).; Rio de Janeiro: Embrapa Solos, 2005; (Boletim De Pesquisa E Desenvolvimento/Embrapa Solos, ISSN 1678-0892; 71). [Google Scholar]
- Lehmann, J.; Kern, D.C.; German, L.; McCann, J.; Martins, G.C.; Moreira, A. Soil fertilityand production potential. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W.I., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 105–124. [Google Scholar]
- Marris, E. Putting the carbon back: Black is the new green. Nature 2006, 442, 624–626. [Google Scholar] [CrossRef]
- Woods, W.I. Development of anthrosol research. In Amazonian Dark Earths: Origin, Properties, and Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W.I., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 3–14. [Google Scholar]
- Coelho, M.R.; Fidalgo, E.C.C.; Araújo, F.O.; dos Santos, H.G.; Mendonça, S.M.L.; Pérez, D.V.; Moreira, F.M.S. 2005; Solos das áreas-piloto do Projeto BiosBrasil (Conservation and Sustainable Management of Below Ground Biodiversity: Phase I). In Município de Benjamim Constant; Estado do Amazonas [recurso eletrônico] Rio de Janeiro: Embrapa Solos.; (Boletim de pesquisa e desenvolvimento/Embrapa Solos, ISSN 1678-0892; 67). [Google Scholar]
- Bentivenga, S.P.; Morton, J.B. Stability and heritability of fatty acid methyl ester profiles of glomalean endomycorrhizal fungi. Mycolog. Res. 1994, 98, 1419–1426. [Google Scholar] [CrossRef]
- Garcia-Martínez, J.; Rodriguez-Valera, F. Microdiversity of uncultured marine prokaryotes: the SAR11 cluster and the marine Archaea of group I. Molecular Ecol. 2000, 9, 935–948. [Google Scholar] [CrossRef]
- Casamayor, E.O.; Massana, R.; Benlloch, S.; Øvreas, L.; Díez, B.; Goddard, V.J.; Gasol, J.M.; Joint, G.I.; Rodríguez-Valera, F.; Pedrós-Alió, C. Changes in archaeal, bacterial and eukaryal assemblages along a salinity gradient by comparison of genetic fingerprinting methods in a multipond solar saltern. Env. Microbiol. 2002, 4, 338–348. [Google Scholar] [CrossRef]
- Sequerra, J.; Marmeisse, R.; Valla, G.; Normand, P.; Capellano, A.; Moiroud, A. Taxonomic position and intraspecific variability of the nodule forming Penicillium nodositatum inferred from RFLP analysis of the ribosomal intergenic spacer and random amplified polymorphic DNA. Mycolog. Res. 1997, 101, 465–472. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blocker, H.; Emde, M.; Bottger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef]
- Woese, C.R.; Kandler, O.; Wheelis, L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria and Eucarya. Proc. Nat. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Kindt, R.; Coe, R.; Tree Diversity Analysis. A Manual and Software for Common Statistical Methods and Biodiversity Studies. World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005. [Google Scholar]
- R Development Core Team. R: A language and environment for statistical computing. ISBN 3-900051-07-0; R Foundation for Statistical Computing: Vienna, Austria, 2007. Available online: http://www.R-project.org (accessed May 2010).
- Wang, Q. Artificial neural networks as cost engineering methods in a collaborative manufacturing environment. Int. J. Prod. Econ. 2007, 109, 53–64. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M,; Tiedje, J.M. The Ribosomal Database Project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, 141–145. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; Sahl, J.W.; Stres, B.; Thallinger, G.G.; Van Horn, D.J.; Weber, C.F. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Borneman, J.; Triplett, E.W. Molecular microbial diversity in soils from Eastern Amazonia: evidence for unusual microorganisms and microbial population shifts associated with deforestation. Appl. Environ. Microbiol. 1997, 63, 2647–2653. [Google Scholar]
- Donovan, S.E.; Purdy, K.J.; Kane, M.D.; Eggleton, P. Comparison of Euryarchaea strains in the guts and food-soil of the soil-feeding termite Cubitermes fungifaber across different soil types. Appl. Environ. Microbiol. 2004, 70, 3884–3892. [Google Scholar] [CrossRef]
- Winter, C.; Smit, A.; Herndl, G.J.; Weinbauer, M.G. Impact of virioplankton on archaeal and bacterial community richness as assessed in seawater batch cultures. Appl. Environ. Microbiol. 2004, 70, 804–813. [Google Scholar] [CrossRef]
- Jesus, E.C.; Marsh, T.L.; Tiedje, J.M.; Moreira, F.M.S. Changes in land use alter the structure of bacterial communities in Western Amazon soils. ISME J. 2009, 3, 1004–1011. [Google Scholar] [CrossRef]
- Rambelli, A.; Persiani, A.M.; Maggi, O.; Lunghini, D.; Onofri, S.; Riess, S.; Dowgiallo, G.; Puppi, G. Comparative studies on microfungi in tropical ecosystems. Mycological studies in South Western Ivory Coast forest. MAB, UNESCO 1983, 102. [Google Scholar]
- Rambelli, A.; Persiani, A.M.; Maggi, O.; Onofri, S.; Riess, S.; Dowgiallo, G.; Zucconi, L. Further mycological studies in south western Ivory Coast forest. Report no. 2. Giornale Botanici Italica 1984, 118, 201–243. [Google Scholar] [CrossRef]
- Maggi, O.; Persiani, A.M.; Casado, M.A.; Pineda, F.D. Edaphic mycoflora recovery in tropical forests after shifting cultivation. Acta Oecologica 1990, 11, 337–350. [Google Scholar]
- Persiani, A.M.; Maggi, O.; Casado, M.A.; Pineda, F.D. Diversity and variability in soil fungi from a disturbed tropical rain forest. Mycologia 1998, 90, 206–214. [Google Scholar] [CrossRef]
- Antoniolli, Z.I.; Schachtman, D.P.; Ophel-Keller, K.; Smith, S.E. Variation in rDNA ITS sequences in Glomus mosseae and Gigaspora margarita spores from a permanent pasture. Mycol. Res. 2000, 104, 708–715. [Google Scholar] [CrossRef]
- Piccolo, M.C.; Neill, C.; Cerri, C.C. Natural abundance of 15N in soils along forest-to-pasture chronosequences in the western Brazilian Amazon Basin. Oecologia 1994, 99, 112–117. [Google Scholar] [CrossRef]
- Reiners, W.A.; Bouwman, A.F.; Parsons, W.F.J.; Keller, M. Tropical rain forest conversion to pasture: changes in vegetation and soil properties. Ecol. Appl. 1994, 4, 363–377. [Google Scholar] [CrossRef]
- Moraes, J.F.L.; Neill, C.; Volkoff, B.; Cerri, C.C.; Melillo, J.; Lima, V.C.; Steudler, P.A. Soil carbon and nitrogen stocks following forest conversion to pasture in the Western Brazilian Amazon Basin. Acta Scientiarum 2002, 24, 1369–1376. [Google Scholar]
- Bünemann, E.K.; Marschner, P.; Smernik, R.J.; Conyers, M.; McNeill, A.M. Soil organic phosphorus and microbial community composition as affected by 26 years of different management strategies. Biol. Fertil. Soils 2008, 44, 717–726. [Google Scholar] [CrossRef]
- Makarov, M.I.; Haumaier, L.; Zech, W.; Marfenina, O.E.; Lysak, L.V. Can 31P NMR spectroscopy be used to indicate the origins of soil organic phosphates? Soil Biol. Biochem. 2005, 37, 15–25. [Google Scholar] [CrossRef]
- Buckley, D.H.; Schmidt, T.M. Diversity and dynamics of microbial communities in soils from agro-ecosystems. Environ. Microbiol. 2003, 5, 441–452. [Google Scholar] [CrossRef]
- Bending, G.D.; Turner, M.K.; Jones, J.E. Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biol. Biochem. 2000, 34, 1073–1082. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impacts of carbon and flooding on soil microbial communities: phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Bulluck, L.R.; Ristaino, J.B. Effect of synthetic and organic soil fertility amendments on southern blight soil microbial communities and yield of processing tomatoes. Phytopathology 2002, 92, 181–189. [Google Scholar] [CrossRef]
- de Brito Alvarez, M.A.; Gagne, S.; Antoun, H. Effect of compost on rhizosphere microflora of the tomato and on the incidence of plant-growth promoting rhizobacteria. Appl. Environ. Microbiol. 1995, 61, 194–199. [Google Scholar]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Duncan, L.C. Is the microbial community in a soil with reduced catabolic diversity less resistant to stress or disturbance? Soil Biol. Biochem. 2001, 33, 1143–1153. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Rice, W.A.; Clayton, G.W. Soil microbial diversity and community structure under wheat as influenced by tillage and crop rotation. Soil Biol. Biochem. 1998, 30, 1733–1741. [Google Scholar] [CrossRef]
- Steenwerth, K.L.; Jackson, L.E.; Calderón, F.J.; Stromberg, M.R.; Scow, K.M. Soil microbial community composition and land use history in cultivated and grassland ecosystems of coastal California. Soil Biol. Biochem. 2003, 35, 489–500. [Google Scholar] [CrossRef]
- Fisher, M.M.; Triplett, E. Automated approach for ribosomal intergenic spacer analysis of microbial diversity and its application to freshwater bacterial communities. Appl. Environ. Microbiol. 1999, 65, 4630–4636. [Google Scholar]
- Ranjard, L.; Poly, F.; Lata, J-C.; Mougel, C.; Thioulouse, J.; Nazaret, S. Characterization of bacterial and fungal soil communities by automated ribosomal intergenic spacer analysis fingerprints: biological and methodological variability. Appl. Environ. Microbiol. 2001, 67, 4479–4487. [Google Scholar] [CrossRef]
- Saison, C.; Degrange, V.; Oliver, R.; Millard, P; Commeaux, C.; Montange, D.; Roux, X.L. Alternation and resilience of the soil microbial community following compost amendment: effects of compost level and compost-borne microbial community. Environ. Microbiol. 2006, 8, 247–257. [Google Scholar] [CrossRef]
- Wu, T.; Chellemi, D.O.; Graham, J.H.; Martin, K.J.; Rosskopf, E.N. Comparison of soil bacterial communities under diverse agricultural land management and crop production practices. Microb. Ecol. 2008, 55, 293–310. [Google Scholar]
- Lima, H.N.; Schaefer, C.E.R.; Mello, J.W.V.; Gilkes, R.J.; Ker, J.C. Pedogenesis and pre-Columbian land use of “Terra Preta Anthrosols” (“Indian black earth”) of Western Amazonia. Geoderma 2002, 110, 1–17. [Google Scholar] [CrossRef]
- Falcão, N.P.S.; Borges, L.F. Efeito da fertilidade de terra preta de índio da Amazônia Central no estado nutricional e na produtividade do mamão hawaí (Carica papaya L.). Acta Amazônica 2006, 36, 401–406. [Google Scholar]
- Lehmann, J.; Silva, J.P.; Steiner, C.; Nehls, T.; Zech, W.; Glaser, B. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: Fertilizer, manure and charcoal amendments. Plant Soil 2003, 249, 343–357. [Google Scholar] [CrossRef]
- Liang, B.; Lehmann , J.; Solomon, D.; Kinyangi, J.; Grossman, J.; O´Neill, B.; Skjemstad, O.; Thies, J.; Luizão, F.J.; Petersen, J.; Neves, E.G. Black carbon increases cation exchange capacity in soils. Soil Sci. Soc. Am. J. 2006, 70, 1719–1730. [Google Scholar]
- Glaser, B.; Lehmann, J.; Zech, W. Ameliorating physical and chemical properties of highly weathered soils in the tropics with charcoal—a review. Biol. Fertil. Soils 2002, 35, 219–230. [Google Scholar] [CrossRef]
- Kim, J.S.; Sparovek, G.; Longo, R.M.; De Melo, W.J.; Crowley, D. Bacterial diversity of terra preta and pristine forest soil from the Western Amazon. Soil Bio. Biochem. 2007, 39, 684–690. [Google Scholar] [CrossRef]
- O’Neill, B.; Grossman, J.; Tsai, S.M.; Gomes, J.E.; Lehmann, J.; Peterson, J.; Neves, E.; Thies, J.E. Bacterial community composition in Brazilian anthrosols and adjacent soils characterized using culturing and molecular identification. Microb. Ecol. 2009, 58, 23–35. [Google Scholar]
- Glaser, B. Prehistorically modified soils of central Amazonia: a model for sustainable agriculture in the twenty-first century. Phil. Trans. R. Soc. B 2007, 362, 187–196. [Google Scholar] [CrossRef]
- Pietikainen, J.; Kiikkila, O.; Fritze, H. Charcoal as a habitat for microbes and its effect on the microbial community of the underlying humus. Oikos 2000, 89, 231–242. [Google Scholar]
- Thies, J.E.; Rillig, M.C. Characteristics of biochar: Biological properties. In Biochar for Environmental Management; Lehmann, J., Joseph, S., Eds.; Earthscan, Dunstan House: London, UK, 2008. [Google Scholar]
- Warnock, D.D.; Lehmann, J.; Kuyper, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil—concepts and mechanisms. Plant Soil 2007, 300, 9–20. [Google Scholar] [CrossRef]
- Liang, B.; Lehmann, J.; Solomon, D.; Kinyangi, J.; Grossman, J.; O’Neill, B.; Skejmstad, J.O.; Thies, J.; Luizao, F.J.; Petersen, J.; Neves, E.G. Black Carbon increases cation exchange capacity in soils. Soil Sci. Soc. Am. J. 2006, 70, 1719–1730. [Google Scholar] [CrossRef]
- Nusslein, K.; Tiedje, J.M. Soil bacterial community shift correlated with change from forest to pasture vegetation in a tropical soil. Appl. Environ. Microb. 1999, 65, 3622–3626. [Google Scholar]
- Pessenda, L.C.R.; Gouveia, S.E.M.; Aravena, R. Radiocarbon dating of total soil organic matter and humin fraction and its comparison with C-14 ages of fossil charcoal. Radiocarbon 2001, 43, 595–601. [Google Scholar]
- Taketani, R.G.; Tsai, S.M. The influence of different land uses on the structure of archaeal communities in Amazonian anthrosols based on 16S rRNA and amoA genes. Microb. Ecol. 2010. [Google Scholar] [CrossRef]
- Glaser, B.; Guggenberger, G.; Zech, W. Organic chemistry studies on Amazonian Dark Earths. In Amazonian Dark Earths: Origin, Properties, and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer: Dordrecht, The Netherlands, 2003; pp. 227–241. [Google Scholar]
- Glaser, B.; Guggenberger, G.; Zech, W.; Ruivo, M.L. Soil organic matter stability in Amazonian Dark Earths. In Amazonian Dark Earths: Origin, Properties, and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer: Dordrecht, The Netherlands, 2003; pp. 141–158. [Google Scholar]
- Lehmann, J.; Liang, B.; Solomon, D.; Lerotic, M.; Luizao, F.; Kinyangi, J.; Schafer, T.; Wirick, S.; Jacobsen, C. Near-edge X-ray absorptive fine structure (NEXAFS) spectroscopy for mapping nano-scale distributions of organic carbon forms in soil: application to .black carbon particles. Glob Biogeochem. Cycles 2005, 19, 1–12. [Google Scholar]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef]
- Tsai, S.M.; O’Neill, B.; Cannavan, F.S.; Saito, D.; Falcão, N.P.S.; Kern, D.; Grossman, J.; Thies, J. Microbial World of Terra Preta. In Terra Preta Nova: A Tribute to Wim Sombroek, 1st ed.; Woods, W.I., Ed.; Springer Verlag: New York, NY, USA, 2008; volume 1, pp. 299–308. [Google Scholar]
- Buckley, D.H.; Schmidt, T.M. Environmental factors in£uencing the distribution of rRNA from Verrucomicrobia in soil. FEMS Microbiol. Ecol. 2001, 35, 105–112. [Google Scholar] [CrossRef]
- Neves, E.G.; Petersen, J.B.; Bartone, R.N.; Silva, C.A. Historical and socio-cultural origins of Amazonian Dark Earths. In Amazonian Dark Earths: Origin, Properties, and Management; Lehmann, J., Kern, D., Glaser, B., Woods, W., Eds.; Kluwer: Dordrecht, The Netherlands, 2003; pp. 3–14. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Navarrete, A.A.; Cannavan, F.S.; Taketani, R.G.; Tsai, S.M. A Molecular Survey of the Diversity of Microbial Communities in Different Amazonian Agricultural Model Systems. Diversity 2010, 2, 787-809. https://doi.org/10.3390/d2050787
Navarrete AA, Cannavan FS, Taketani RG, Tsai SM. A Molecular Survey of the Diversity of Microbial Communities in Different Amazonian Agricultural Model Systems. Diversity. 2010; 2(5):787-809. https://doi.org/10.3390/d2050787
Chicago/Turabian StyleNavarrete, Acácio A., Fabiana S. Cannavan, Rodrigo G. Taketani, and Siu M. Tsai. 2010. "A Molecular Survey of the Diversity of Microbial Communities in Different Amazonian Agricultural Model Systems" Diversity 2, no. 5: 787-809. https://doi.org/10.3390/d2050787
APA StyleNavarrete, A. A., Cannavan, F. S., Taketani, R. G., & Tsai, S. M. (2010). A Molecular Survey of the Diversity of Microbial Communities in Different Amazonian Agricultural Model Systems. Diversity, 2(5), 787-809. https://doi.org/10.3390/d2050787