
Marine Biodiversity in Temperate Western Australia: Multi-Taxon Surveys of Minden and Roe Reefs

 ,
,  , , , add
Show full author list
, , , add
Show full author list
Abstract
:
1. Introduction
2. Results
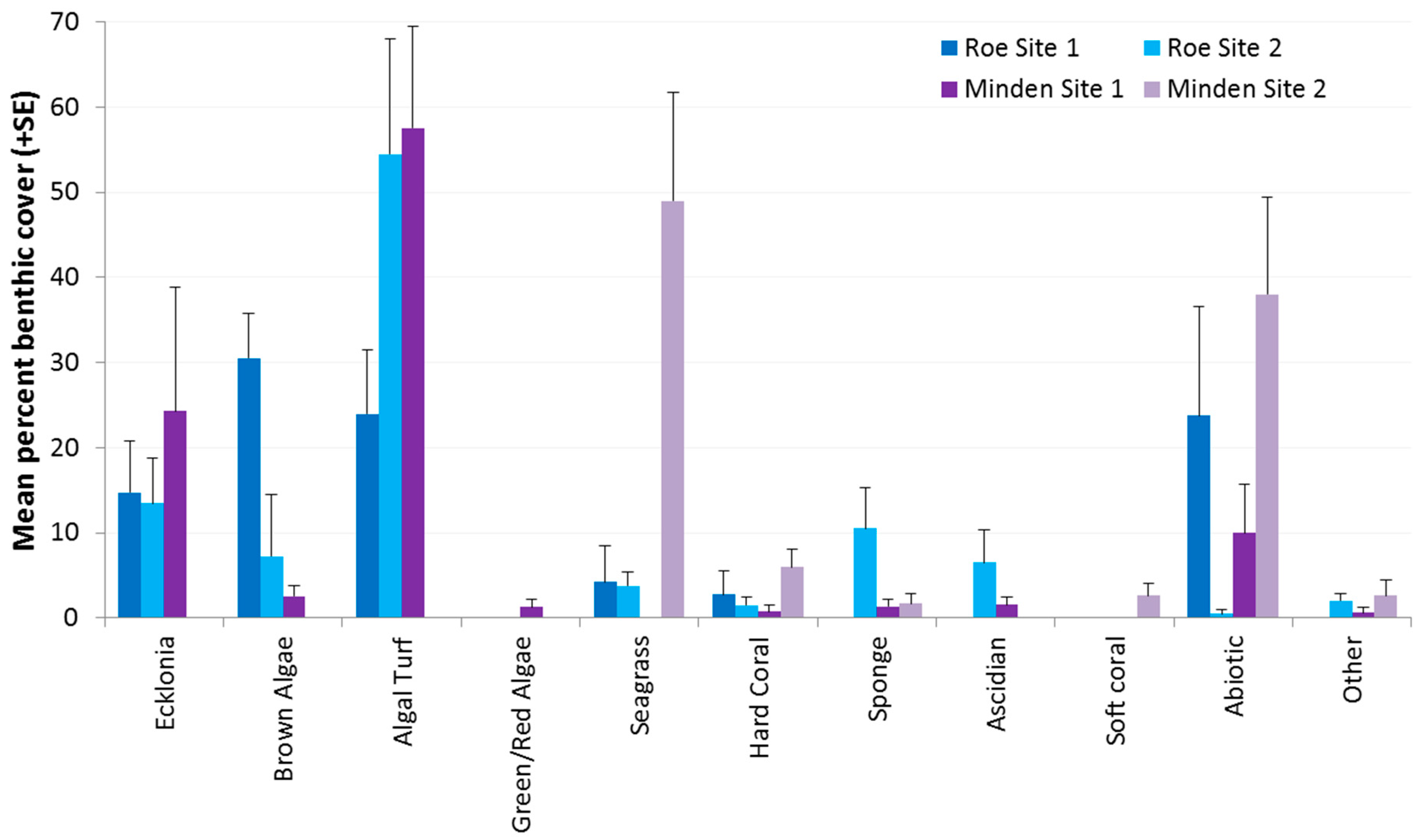
2.1. Benthic Cover
2.2. Hard Corals
2.3. Soft Corals (Octocorals)
2.4. Sponges
2.5. Molluscs
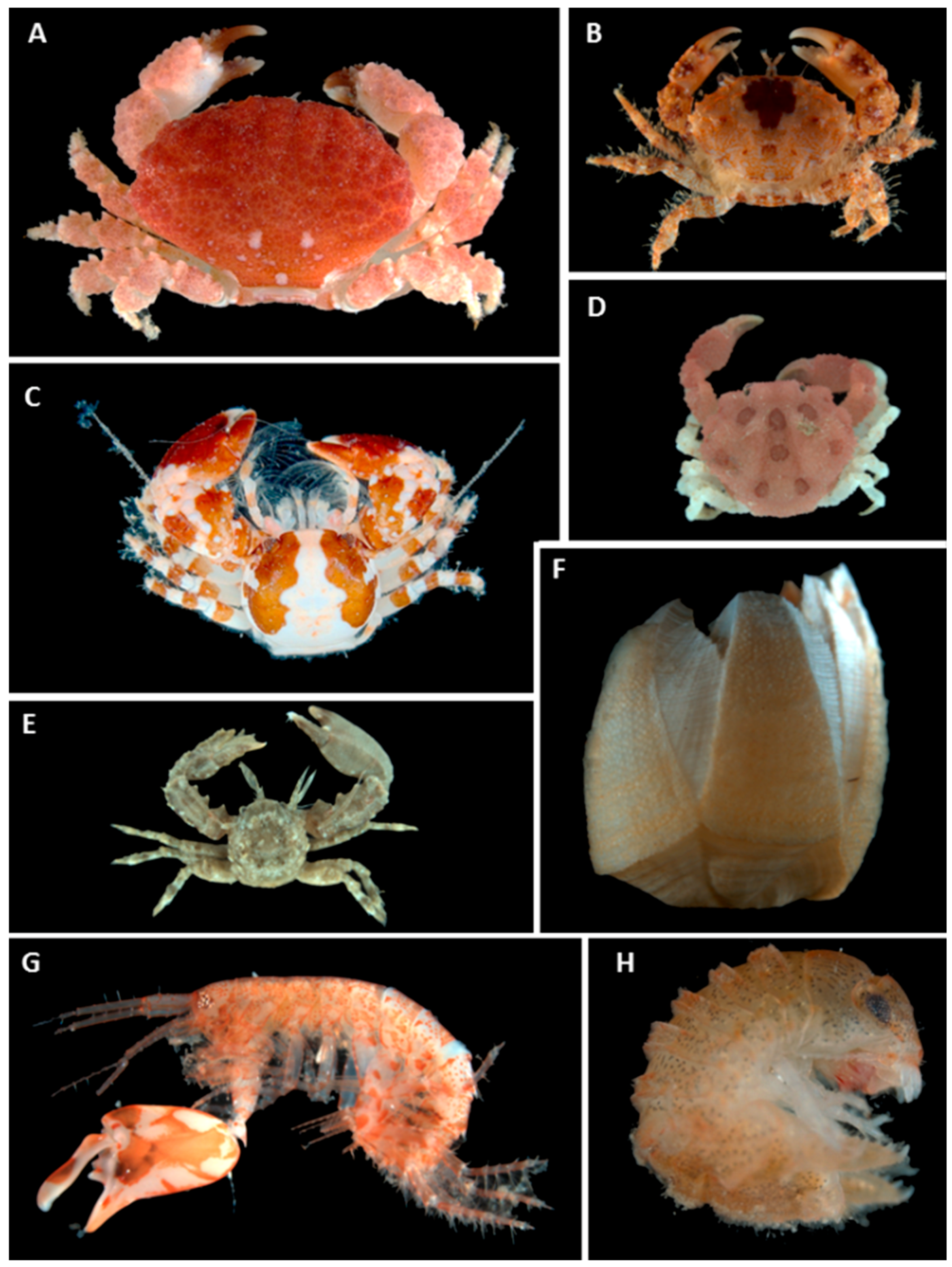
2.6. Crustaceans
2.7. Echinoderms
2.8. Fishes
2.9. Marine Plants
3. Discussion
4. Materials and Methods
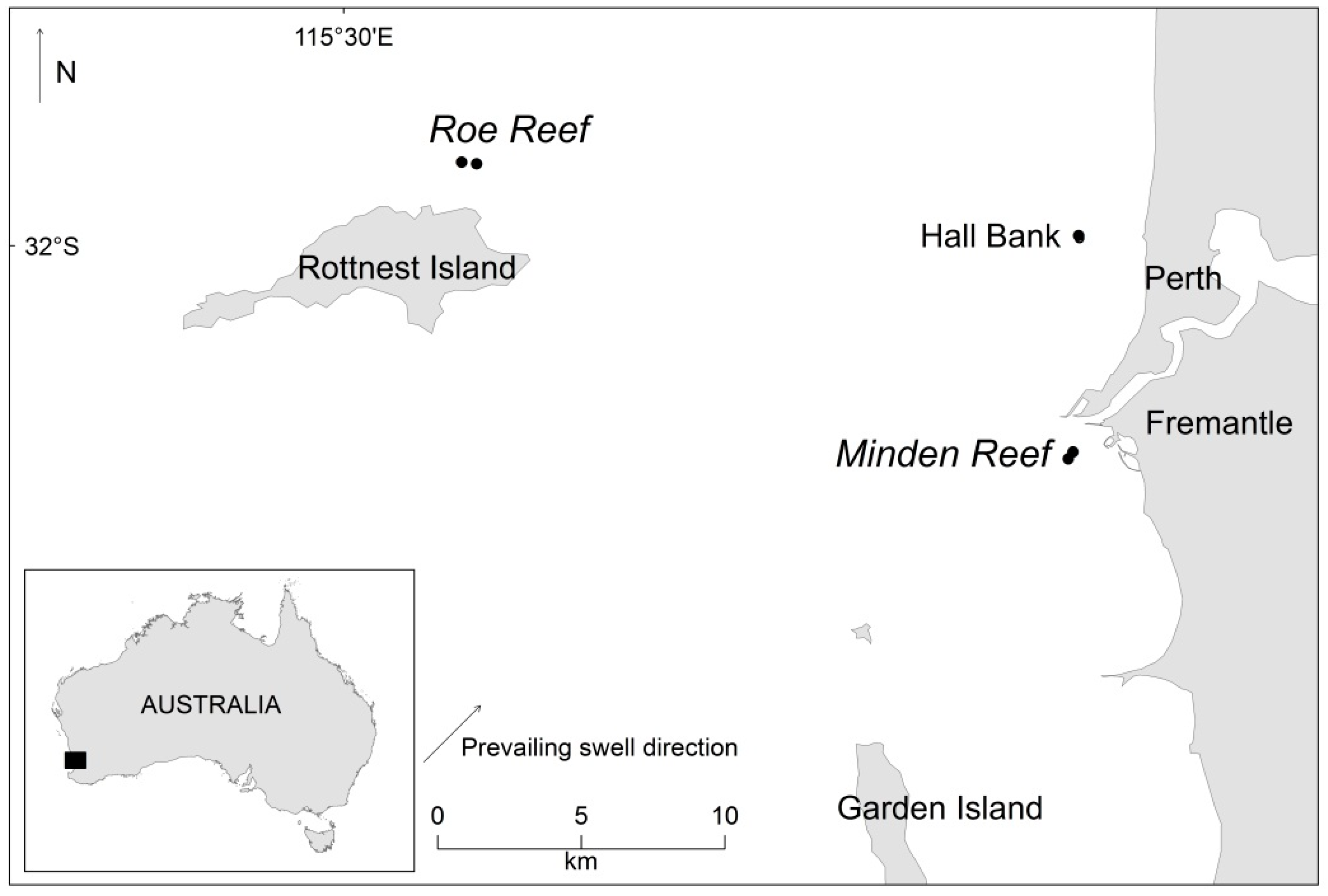
4.1. Site Description
4.2. Field Surveys
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rosenzweig, C.; Neofotis, P. Detection and attribution of anthropogenic climate change impacts. WIREs Clim. Change 2013, 4, 121–150. [Google Scholar] [CrossRef]
- Precht, W.F.; Aronson, R.B. Climate flickers and range shifts of reef corals. Front. Ecol. Environ. 2004, 2, 307–314. [Google Scholar] [CrossRef]
- Ling, S.D.; Johnson, C.R.; Ridgway, K.; Hobday, A.J.; Haddon, M. Climate-driven range extension of a sea urchin: Inferring future trends by analysis of recent population dynamics. Glob. Change Biol. 2009, 15, 719–731. [Google Scholar] [CrossRef]
- Last, P.R.; White, W.T.; Gledhill, D.C.; Hobday, A.J.; Brown, R.; Edgar, G.J.; Pecl, G. Long-term shifts in abundance and distribution of a temperate fish fauna: A response to climate change and fishing practices. Glob. Ecol. Biogeogr. 2011, 20, 58–72. [Google Scholar] [CrossRef]
- Pitt, N.R.; Poloczanska, E.S.; Hobday, A.J. Climate-driven range changes in Tasmanian intertidal fauna. Mar. Freshw. Res. 2010, 61, 963–970. [Google Scholar] [CrossRef]
- Yamano, H.; Sugihara, K.; Nomura, K. Rapid poleward range expansion of tropical reef corals in response to rising sea surface temperatures. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Baird, A.H.; Sommer, B.; Madin, J.S. Pole-ward range expansion of Acropora spp. along the east coast of Australia. Coral Reefs 2012, 31, 1063. [Google Scholar] [CrossRef]
- Nakamura, Y.; Feary, D.A.; Kanda, M.; Yamaoka, K. Tropical fishes dominate temperate reef fish communities within Western Japan. PLoS ONE 2013, 8, e81107. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Harasti, D.; Davis, T.; Smith, S.D. Southernmost records of the host sea anemone, Stichodactyla haddoni, and associated commensal shrimps in a climate change hotspot. Mar. Biodivers. 2014, 45, 1–2. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Willan, R.C.; Smith, S.D. Range extensions for heterobranch sea slugs (formerly opisthobranch) belonging to the families diaphanidae, Plakobranchidae and Facelinidae on the eastern coast of Australia. Mar. Biodivers. Rec. 2015, 8, e76. [Google Scholar] [CrossRef]
- Przeslawski, R.; Ahyong, S.; Byrne, M.; Worheide, G.; Hutchings, P. Beyond coral and fish: The effects of climate change on non-coral benthic invertebrates of tropical reefs. Glob. Change Biol. 2008, 14, 2773–2795. [Google Scholar] [CrossRef]
- Przeslawski, R.; Falkner, I.; Ashcroft, M.B.; Hutchings, P. Using rigorous selection criteria to investigate marine range shifts. Estuar. Coast. Shelf Sci. 2012, 113, 205–212. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Babcock, R.C.; Butler, A.; Hobday, A.J.; Hoegh-Guldberg, O.; Kunz, T.J.; Matear, R.; Milton, D.A.; Okey, T.A.; Richardson, A.J. Climate change and Australian marine life. Oceanogr. Mar. Biol. 2007, 45, 407–478. [Google Scholar]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Wernberg, T.; Russell, B.D.; Moore, P.J.; Ling, S.D.; Smale, D.A.; Campbell, A.; Coleman, M.A.; Steinberg, P.D.; Kendrick, G.A.; Connell, S.D. Impacts of climate change in a global hotspot for temperate marine biodiversity and ocean warming. J. Exp. Mar. Biol. Ecol. 2011, 400, 7–16. [Google Scholar] [CrossRef]
- Cacciapaglia, C.; Woesik, R. Reef-coral refugia in a rapidly changing ocean. Glob. Change Biol. 2015, 21, 2272–2282. [Google Scholar] [CrossRef] [PubMed]
- Waters, J.M.; Wernberg, T.; Connell, S.D.; Thomsen, M.S.; Zuccarello, G.C.; Kraft, G.T.; Sanderson, J.C.; West, J.A.; Gurgel, C.F.D. Australia’s marine biogeography revisited: Back to the future? Aust. Ecol. 2010, 35, 988–992. [Google Scholar] [CrossRef]
- Huber, M. Compendium of Bivalves. A Full Color Guide to 3300 of the World’s Marine Bivalves. A Status on Bivalvia after 250 Years of Research; Conchbooks: Hackenheim, Geramny, 2010; Volume 1, p. 901. [Google Scholar]
- Wilson, B.R.; Allen, G.R. Major components and distribution of marine fauna. In Fauna of Australia; Walton, G.R.D.D.W., Ed.; Australian Government Publishing Service: Canberra, Australia, 1987; Volume 1a, pp. 43–68. [Google Scholar]
- Ponder, W.; Hutchings, P.; Chapman, R. Overview of the Conservation of Australian Marine Invertebrates; A Report for Environment Australia; Australian Museum: Sydney, Australia, 2002; p. 588. [Google Scholar]
- Phillips, J.A. Marine macroalgal biodiversity hotspots: Why is there high species richness and endemism in southern Australian marine benthic flora? Biodivers. Conserv. 2001, 10, 1555–1577. [Google Scholar] [CrossRef]
- Cork, S.; Sattler, P.; Alexandra, J. Biodiversity Theme Commentary Prepared for the 2006 Australian State of the Environment Committee. Available online: http://www.deh.gov.au/soe/2006/commentaries/biodiversity/index.html (accessed on 12 September 2015).
- Pearce, A.; Feng, M. Observations of warming on the Western Australian continental shelf. Mar. Freshw. Res. 2007, 58, 914–920. [Google Scholar] [CrossRef]
- Ling, S.D. Range expansion of a habitat-modifying species leads to loss of taxonomic diversity: A new and impoverished reef state. Oecologia 2008, 156, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.C. Partitioning herbivory and its effects on coral-reef algal communities. Ecol. Monogr. 1986, 56, 345–364. [Google Scholar] [CrossRef]
- Johnson, C.R.; Banks, S.C.; Barrett, N.S.; Cazassus, F.; Dunstan, P.K.; Edgar, G.J.; Frusher, S.D.; Gardner, C.; Haddon, M.; Helidoniotis, F.; et al. Climate change cascades: Shifts in oceanography, species’ ranges and subtidal marine community dynamics in eastern tasmania. J. Exp. Mar. Biol. Ecol. 2011, 400, 17–32. [Google Scholar] [CrossRef]
- Vergés, A.; Steinberg, P.D.; Hay, M.E.; Poore, A.G.B.; Campbell, A.H.; Ballesteros, E.; Heck, K.L.; Booth, D.J.; Coleman, M.A.; Feary, D.A.; et al. The tropicalization of temperate marine ecosystems: Climate-mediated changes in herbivory and community phase shifts. Proc. R. Soc. Lond. B Biol. Sci. 2014, 281, 20140846. [Google Scholar] [CrossRef] [PubMed]
- Caputi, N.; Melville-Smith, R.; Lestang, S.D.; Pearce, A.; Feng, M. The effect of climate change on the Western rock lobster (Panulirus cygnus) fishery of Western Australia. Can. J. Fish. Aquat. Sci. 2010, 67, 85–96. [Google Scholar]
- Stachowicz, J.J.; Terwin, J.R.; Whitlatch, R.B.; Osman, R.W. Linking climate change and biological invasions: Ocean warming facilitates nonindigenous species invasions. Proc. Natl. Acad. Sci. 2002, 99, 15497–15500. [Google Scholar] [CrossRef] [PubMed]
- Thresher, R.; Proctor, C.; Ruiz, G.; Gurney, R.; MacKinnon, C.; Walton, W.; Rodriguez, L.; Bax, N. Invasion dynamics of the european shore crab, Carcinus maenas, in Australia. Mar. Biol. 2003, 142, 867–876. [Google Scholar]
- Zeidberg, L.D.; Robison, B.H. Invasive range expansion by the Humboldt squid, Dosidicus gigas, in the eastern north pacific. Proc. Natl. Acad. Sci. 2007, 104, 12948–12950. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.S.; Hazen, E.L.; Bograd, S.J.; Byrnes, J.E.K.; Foley, D.G.; Gilly, W.F.; Robison, B.H.; Field, J.C. Combined climate- and prey-mediated range expansion of Humboldt squid (Dosidicus gigas), a large marine predator in the california current system. Glob. Change Biol. 2014, 20, 1832–1843. [Google Scholar] [CrossRef] [PubMed]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, B.J.; Pandolfi, J.M. Escaping the heat: Range shifts of reef coral taxa in coastal Western Australia. Glob. Change Biol. 2008, 14, 513–528. [Google Scholar] [CrossRef]
- Eliot, I.; Nutt, C.; Gozzard, B.; Higgins, M.; Buckley, E.; Bowyer, J. Coastal Compartments of Western Australia: A Physical Framework for Marine and Coastal Planning; Report to the Departments of Environment & Conservation, Planning and Transport; Damara W.A. Pty Ltd, Geological Survey of Western Australia; Department of Environment & Conservation: Western Australia, Australia, 2011. [Google Scholar]
- Veron, J.E.N.; Marsh, L.M. Hermatypic corals of Western Australia. Records and annotated species list. Rec. West. Aust. Mus. Suppl. 1988, 29, 1–136. [Google Scholar]
- Thomson, D.P.; Frisch, A.J. Extraordinarily high coral cover on a nearshore, high-latitude reef in south-west Australia. Coral Reefs 2010, 29, 923–927. [Google Scholar] [CrossRef]
- Hutchins, J.B.; Pearce, A.F. Influence of the Leeuwin current on recruitment of tropical reef fishes at Rottnest Island, Western Australia. Bull. Mar. Sci. 1994, 54, 245–255. [Google Scholar]
- Jones, D.S.; Morgan, G.J. An annotated checklist of the crustacea of Rottnest Island, Western Australia. In The Marine Flora and Fauna of Rottnest Island, Western Australia, Proceedings of the Fifth International Marine Biological Workshop, Rottnest Island, Australia, 5 January 1991; Wells, F.E., Walker, D.I., Kirkman, H., Lethbridge, R., Eds.; Western Australian Museum: Perth, Australia, 1993; Volume 1, pp. 135–162. [Google Scholar]
- Huisman, J.M.; Walker, D.I. A catologue of the marine plants from Rottnest Island, Western Australia, with notes on their distribution and biogeography. Kingia 1990, 1, 365–481. [Google Scholar]
- Wells, F.E.; Walker, D.I.; Kirkman, H.; Lethbridge, R. (Eds.) The Marine Flora and Fauna of Rottnest Island, Western Australia: Proceedings of the Fifth International Marine Biological Workshop, Rottnest Island, Australia; Western Australian Museum: Perth, Australia, 1993; Volume 1.
- Wells, F.E.; Walker, D.I.; Kirkman, H.; Lethbridge, R. (Eds.) The Marine Flora and Fauna of Rottnest Island, Western Australia: Proceedings of the Fifth International Marine Biological Workshop, Rottnest Island, Australia; Western Australian Museum: Perth, Australia, 1993; Volume 2.
- Walker, D.I.; Wells, F.E. The Seagrass Flora and Fauna of Rottnest Island, Western Australia; Western Australian Museum: Perth, Australia, 1999; p. 421. [Google Scholar]
- Farrant, P.A. Reproduction in the temperate Australian soft coral Capnella gaboensis. In Proceedings of the Fifth International Coral Reef Congress, Tahiti, 27 May–1 June 1985; Volume 4, pp. 319–324.
- Verseveldt, J. Australian Octocorallia (Coelenterata). Aust. J. Mar. Freshw. Res. 1977, 28, 171–240. [Google Scholar] [CrossRef]
- Alderslade, P. Four new genera of soft corals (coelenterata: Octocorallia), with notes on the classification of some established taxa. Zool. Meded. 2000, 74, 1–17. [Google Scholar]
- Fromont, J.; Huggett, M.J.; Lengger, S.L.; Grice, K.; Schönberg, C.H. Characterisation of Leucetta prolifera, a calcarean cyanosponge from south-Western Australia, and its symbionts. J. Mar. Biol. Assoc. UK 2016, 96, 541–552. [Google Scholar] [CrossRef]
- Hentschel, E. Tetraxonida. In Die Fauna Südwest-Australiens Ergebnisse der Hamburger Südwest-Australischen Forschungsreise, 1905; Michaelsen, W., Hartmeyer, R., Eds.; Fischer: Jena, Germany, 1909; Volume 2, pp. 347–402. [Google Scholar]
- Morrison, H.; Wells, F.E. Colonisation of fremantle harbour and cockburn sound, Western Australia by the eastern Australian scallop Scaeochlamys livida (lamarck, 1819). Molluscan Res. 2008, 28, 107–110. [Google Scholar]
- Knudsen, S.W.; Clements, K.D. Kyphosus gladius, a new species of sea chub from Western Australia (Teleostei: Kyphosidae), with comments on segutilum klunzingeri whitley. Zootaxa 2013, 3599, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.M. Marine Plants of Australia; University of Western Australia Press: Perth, Australia, 2000; p. 300. [Google Scholar]
- Smale, D.A.; Wernberg, T. Extreme climatic event drives range contraction of a habitat-forming species. Proc. Biol. Sci. 2013, 280, 20122829. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.M. The type and Australian species of the red algal genera liagora and ganonema (Liagoraceae, Nemaliales). Aust. Syst. Bot. 2002, 15, 773–838. [Google Scholar] [CrossRef]
- Lin, S.-M.; Yang, S.-Y.; Huisman, J.M. Systematic revision of the genera Liagora and Izziella (Liagoraceae, Rhodophyta) from Taiwan based on lsu and rbcl sequencing analyses and carposporophyte development with description of I. hommersandii sp. nov. and I. kuroshioensis sp. nov. J. Phycol. 2011, 47, 352–365. [Google Scholar] [CrossRef]
- Huisman, J.M. Algae of Australia: Nemaliales; ABRS: Canberra, Australia, 2006. [Google Scholar]
- Hutchings, P.A.; Ahyong, S.T.; Ashcroft, M.B.; McGrouther, M.A.; Reid, A.L. Sydney harbour: Its diverse biodiversity. Aust. Zool. 2013, 36, 255–320. [Google Scholar] [CrossRef]
- Jones, D.S. The barnacles of Rottnest Island, Western Australia, with descriptions of two new species. In The Marine Flora and Fauna of Rottnest Island, Western Australia, Proceedings of the Fifth International Marine Biological Workshop, Rottnest Island, Australia, 5 January 1991; Wells, F.E., Walker, D.I., Kirkman, H., Lethbridge, R., Eds.; Western Australian Museum: Perth, Australia, 1993; Volume 1, pp. 105–133. [Google Scholar]
- Gales, C.F. Mr c.F. Gales report. Western Mail, 1907; 60. [Google Scholar]
- Fromont, J. Reproduction of some demosponges in a temperate Australian shallow water habitat. Mem. Qld. Mus. 1999, 44, 185–192. [Google Scholar]
- Sampey, A.; Fromont, J.; Johnston, D.J. Demersal and epibenthic fauna in a temperate marine embayment, Cockburn Sound, Western Australia: Determination of key indicator species. J. Roy. Soc. West. Aust. 2011, 94, 1–18. [Google Scholar]
- Hall-Spencer, J.M.; Allen, R. The impact of CO2 emissions on “nuisance” marine species. Res. Rep. Biodivers. Stud. 2015, 33, 33–46. [Google Scholar] [CrossRef]
- Feng, M.; Caputi, N.; Pearce, A. Leeuwin current. In In a Marine Climate Change Impacts and Adaptation Report Card for Australia 2012; Poloczanska, E.S., Hobday, A.J., Richardson, A.J., Eds.; CSIRO Marine and Atmospheric Research: Brisbane, Australia, 2012; pp. 61–80. [Google Scholar]
- Hutchins, J.B. Dispersal of tropical fishes to temperate seas in the southern hemisphere. J. R. Soc. West. Aust. 1991, 74, 79–84. [Google Scholar]
- Cure, K.; Hobbs, J.-P.A.; Harvey, E.S. High recruitment associated with increased sea temperatures towards the southern range edge of a Western Australian endemic reef fish Choerodon rubescens (family Labridae). Environ. Biol. Fish. 2014, 98, 1059–1067. [Google Scholar] [CrossRef]
- Pearce, A.F.; Hutchins, J.B. Record-Breaking Temperatures and Tropical Fish Recruitment at Rottnest Island; Fisheries Research Report [Western Australia]; IMAS: Perth, Australia, 2014; pp. 25–26. [Google Scholar]
- Wernberg, T.; Smale, D.A.; Tuya, F.; Thomsen, M.S.; Langlois, T.J.; de Bettignies, T.; Bennett, S.; Rousseaux, C.S. An extreme climatic event alters marine ecosystem structure in a global biodiversity hotspot. Nat. Clim. Change 2013, 3, 78–82. [Google Scholar] [CrossRef]
- Bennett, S.; Wernberg, T.; Harvey, E.S.; Santana-Garcon, J.; Saunders, B.J. Tropical herbivores provide resilience to a climate-mediated phase shift on temperate reefs. Ecol. Lett. 2015, 18, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Thomson, D.P.; Bearham, D.; Graham, F.; Eagle, J.V. High latitude, deeper water coral bleaching at Rottnest Island, Western Australia. Coral Reefs 2011, 30, 1107. [Google Scholar] [CrossRef]
- Moore, J.A.; Bellchambers, L.M.; Depczynski, M.R.; Evans, R.D.; Evans, S.N.; Field, S.N.; Friedman, K.J.; Gilmour, J.P.; Holmes, T.H.; Middlebrook, R.; et al. Unprecedented mass bleaching and loss of coral across 12 of latitude in Western Australia in 2010–2011. PLoS ONE 2012, 7, e51807. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Pearce, A.F. The rise and fall of the “marine heat wave” off Western Australia during the summer of 2010/2011. J. Mar. Syst. 2013, 111–112, 139–156. [Google Scholar]
- Bennett, S.; Wernberg, T.; Connell, S.D.; Hobday, A.J.; Johnson, C.R.; Poloczanska, E.S. The “Great Southern Reef”: Social, ecological and economic value of Australia’s neglected kelp forests. Mar. Freshw. Res. 2015, 67, 47–56. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hard Corals | Octocorals | Sponges | Molluscs | Crustaceans | Echinoderms | Fishes | Marine Plants | Total | |
|---|---|---|---|---|---|---|---|---|---|
| Minden Reef | |||||||||
| # Species | 12 | 4 | 27 | 48 | 34 | 16 | 46 | NA | 188 |
| # Specimens collected | 27 | 7 | 44 | 27 * | 157 | 17 | 0 | NA | 252 ^ |
| # Visual Records | 2 | 0 | 2 | 282 | 0 | 0 | 46 | NA | 332 |
| Roe Reef | |||||||||
| # Species | 9 | 4 | 48 | 40 | 45 | 16 | 56 | 69 | 287 |
| # Specimens collected | 22 | 4 | 38 | 58 * | 140 | 18 | 0 | 2 | 224 ^ |
| # Visual Records | 0 | 0 | 12 | 10 | 1 | 0 | 56 | 67 | 146 |
| Phylum | Species | Putative New Species | Range Extension | Endemic to South-Western Australia |
|---|---|---|---|---|
| Arthropoda | Acasta sp. nov. | • | ||
| Arthropoda | Acasta fenestrata | • | ||
| Arthropoda | Armatobalanus allium | • | ||
| Arthropoda | Cyclax spinicinctus | • | ||
| Arthropoda | Liomera margaritata | • | ||
| Arthropoda | Membranobalanus sp. 1 | • | ||
| Arthropoda | Pyrogoma cancellata | • | ||
| Arthropoda | Pyrogospongia sp. nov. | • | ||
| Arthropoda | Synalpheus bispinosus | • | ||
| Arthropoda | Zalasius dromiaeformis | • | ||
| Chordata | Caesioscorpis theagenes | • | ||
| Chordata | Cheilodactylus gibbosus | • | ||
| Chordata | Choerodon rubescens | • | ||
| Chordata | Chromis klunzingeri | • | ||
| Chordata | Cirripectes hutchinsi | • | ||
| Chordata | Coris auricularis | • | ||
| Chordata | Dasyatis brevicaudata | • | ||
| Chordata | Dotolabrus alleni | • | ||
| Chordata | Epinephelides armatus | • | ||
| Chordata | Glaucosoma hebraicum | • | ||
| Chordata | Halichoeres brownfieldi | • | ||
| Chordata | Kyphosus cornelii | • | ||
| Chordata | Kyphosus gladius | • | ||
| Chordata | Ostorhinchus victoriae | • | ||
| Chordata | Parma mccullochi | • | ||
| Chordata | Parma occidentalis | • | ||
| Chordata | Pseudolabrus biserialis | • | ||
| Chordata | Suezichthys cyanoleamus | • | ||
| Chordata | Trachinops brauni | • | ||
| Chordata | Trygonoptera ovalis | • | ||
| Cnidaria | Alertigorgia mjoebergi | • | ||
| Cnidaria | Capnella gaboensis | • | ||
| Cnidaria | Favites rotundata | • | ||
| Echinodermata | Amphiura brachyactis | • | ||
| Echinodermata | Macrophiothrix michaelseni | • | ||
| Mollusca | Chama pacifica | • | ||
| Porifera | Cymbastela marshae | • | ||
| Porifera | Echinochalina spongiosa | • | ||
| Porifera | Haliclona sp. | • | ||
| Porifera | Leucetta prolifera | • | ||
| Porifera | Protosuberites epiphytum | • | ||
| Plantae | Tricleocarpa new sp. | • | ||
| TOTAL | 3 | 15 | 24 |
| Class | Species | Previous Known Range | km Range Extension | Possible Explanation for Range Extension |
|---|---|---|---|---|
| Arthropoda | Acasta fenestrata | Dampier, WA | ~1265 km | Most likely overlooked |
| Arthropoda | Armatobalanus allium | Abrolhos Is., WA | ~410 km | Most likely overlooked |
| Arthropoda | Cyclax spinicinctus | Abrolhos Is., WA | ~410 km | Most likely overlooked |
| Arthropoda | Liomera margaritata | Abrolhos Is., WA | ~410 km | Most likely overlooked |
| Arthropoda | Membranobalanus sp. 1 | Arafura Sea, NT | ~3380 km | Genus most likely overlooked. Requires further investigation to confirm species |
| Arthropoda | Pyrogoma cancellata | Abrolhos Is., WA | ~410 km | Possibly overlooked or misidentified in past surveys |
| Arthropoda | Synalpheus bispinosus | Dampier, WA | ~1265 km | Possibly overlooked or misidentified in past surveys |
| Arthropoda | Zalasius dromiaeformis | Gulf of Carpentaria, QLD | ~3000 km | Most likely genuine range extension |
| Cnidaria | Alertigorgia mjoebergi | North-West Australia, North of Exmouth, WA | ~1300 km | Most likely genuine range extension |
| Cnidaria | Capnella gaboensis | Southern Queensland, New South Wales, Victoria to South Australia (Waldegrave Island) | ~2300 km | Most likely overlooked but possibly a genuine range extension |
| Cnidaria | Favites rotundata | Indo-west Pacific, including east and western Australia. Most southerly WA record is Abrolhos Is., WA | ~410 km | Possibly overlooked or misidentified in past surveys |
| Echinodermata | Amphiura brachyactis | Roebuck Bay, WA | ~1696 km | Most likely genuine range extension from larval dispersal |
| Mollusca | Chama pacifica | Indo-west Pacific, previously known from Broome, WA | ~1700 km | Most likely genuine range extension from larval dispersal |
| Porifera | Echinochalina spongiosa | Only known from the type locality—Port Phillip Bay, VIC | ~2740 km | Most likely genuine range extension with lack of sampling in between localities causing disjunct distribution |
| Porifera | Protosuberites epiphytum | Great Barrier Reef, Port Phillip Bay VIC and Shark Bay, WA | ~690 km | Most likely genuine range extension with lack of sampling in between localities causing disjunct distribution |
| Location | Site | Aspect | Depth | Latitude (South) | Longitude (East) | Exposure |
|---|---|---|---|---|---|---|
| Minden Reef 13th March 2013 | Site 1 | North | 8–10 m | 32.0401° | 115.4362° | Partially protected from prevailing south-west winds by an interconnecting reef system between Garden and Rottnest Islands. It lies within a sediment deposition zone. |
| Site 2 | South | 8–10 m | 32.0408° | 115.4362° | ||
| Roe Reef 6th February 2014 | Site 1 | East | 20–25 m | 31.9743° | 115.5417° | Protected from prevailing south-west winds and associated wind-driven seas. Subject to winter swells wrapping around the western end of Rottnest Island, which run along the northern side and over the reef. Larger swells break producing high-energy wave action across the reef at these times. |
| Site 2 | Central | 10–12 m | 31.9738° | 115.5370° |
| Data/Taxon | Method | Sample Preservation | Surveyor, Collector And/Or Taxonomist |
|---|---|---|---|
| Benthic Cover | Four 15 m point-intercept transects—every 15 cm under tape = 100 points per transect | NA | Clay Bryce |
| Hard Corals | Four 15 m × 1 m wide belt transects plus 20 min swim off-transect | 100% ethanol (wet); bleach (dry) | Zoe Richards |
| Soft Corals | Four 15 m × 1 m wide belt transects plus 20 min swim off-transect | 70% ethanol | Monika Bryce |
| Sponges | Four 15 m × 1 m wide belt transects | 70% ethanol | Jane Fromont, Oliver Gomez |
| Molluscs | Four 15 m × 1 m wide belt transects plus 20 min swim off-transect | 100% ethanol | Lisa Kirkendale, Corey Whisson, Nerida Wilson, Glad Hansen, Hugh Morrison |
| Crustacea | Four 15 m × 1 m wide belt transects plus representative samples of potential habitat such as sponges, seaweeds and rubble, collected and sorted in the laboratory. Additional ad hoc off-transect collections | 100% ethanol | Andrew Hosie, Ana Hara, Lee Betterridge |
| Echinoderms | Incidental collections (Minden Reef and Roe Reef Site 1 only) | 100% ethanol | Courtney Wood, Jenelle Ritchie, Loisette Marsh |
| Fishes | Six 50 m × 5 m wide belt transects | UVC or photographed | Glenn Moore, Sue Morrison |
| Flora | One hour rapid visual assessment (Roe Reef only) | Pressed; dried in silica gel | John Huisman |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richards, Z.; Kirkendale, L.; Moore, G.; Hosie, A.; Huisman, J.; Bryce, M.; Marsh, L.; Bryce, C.; Hara, A.; Wilson, N.; et al. Marine Biodiversity in Temperate Western Australia: Multi-Taxon Surveys of Minden and Roe Reefs. Diversity 2016, 8, 7. https://doi.org/10.3390/d8020007
Richards Z, Kirkendale L, Moore G, Hosie A, Huisman J, Bryce M, Marsh L, Bryce C, Hara A, Wilson N, et al. Marine Biodiversity in Temperate Western Australia: Multi-Taxon Surveys of Minden and Roe Reefs. Diversity. 2016; 8(2):7. https://doi.org/10.3390/d8020007
Chicago/Turabian StyleRichards, Zoe, Lisa Kirkendale, Glenn Moore, Andrew Hosie, John Huisman, Monika Bryce, Loisette Marsh, Clay Bryce, Ana Hara, Nerida Wilson, and et al. 2016. "Marine Biodiversity in Temperate Western Australia: Multi-Taxon Surveys of Minden and Roe Reefs" Diversity 8, no. 2: 7. https://doi.org/10.3390/d8020007
APA StyleRichards, Z., Kirkendale, L., Moore, G., Hosie, A., Huisman, J., Bryce, M., Marsh, L., Bryce, C., Hara, A., Wilson, N., Morrison, S., Gomez, O., Ritchie, J., Whisson, C., Allen, M., Betterridge, L., Wood, C., Morrison, H., Salotti, M., ... Fromont, J. (2016). Marine Biodiversity in Temperate Western Australia: Multi-Taxon Surveys of Minden and Roe Reefs. Diversity, 8(2), 7. https://doi.org/10.3390/d8020007