Central and Eastern European Spring Pollen Allergens and Their Expression Analysis—State of the Art

Abstract
:1. North, Central and Eastern Europe Spring Pollinosis Sources
2. Spring Tree Pollinosis—Actual Allergen Nucleic Acids Sequences and Function Knowledge
3. Birch Pollen Allergens—Actual DNA/RNA Sequence Data
4. Minor Central and Eastern Pollen Allergens—Actual DNA/RNA Sequence Data
5. Hazelnut Pollen Allergens—Actual DNA/RNA Sequence Data
6. Spring Tree Pollinosis—Expression Analysis of Bet v1, Bet v2, Cor a 1 and Cor a 2 in Pollen Grains from DifferentIn Situ Conditions
7. Conclusions
Acknowledgments
Conflicts of Interest
List of Abbreviations
| BLAST | Basic Local Alignment Search Tool |
| HRM | High Resolution Melting |
| qRT-PCR | quantitative Real Time Polymerase Chain Reaction |
| NCBI | National Center for Biotechnology Information |
| PCR | Polymerase Chain Reaction |
References
- M’Rabet, L.; Vos, A.P.; Boehm, G.; Garssen, J. Breast-Feeding and Its Role in Early Development of the Immune System in Infants: Consequences for Health Later in Life. J. Nutr. 2008, 138, 1782S–1790S. [Google Scholar] [PubMed]
- Kay, A.B. Allergy and Allergic Diseases; Blackwell Publishing Ltd.: Oxford, UK, 2008. [Google Scholar]
- Buters, J.T.; Kasche, A.; Weichenmeier, I. Year-to-year variation in release of Bet v1 allergen from birch pollen: Evidence for geographical differences between West and South Germany. Int. Arch. Allergy Immunol. 2007, 145, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.E.; Jacobson, K.W.; House, J.M.; Glovsky, M.M. Links between pollen, atopy and the asthma epidemic. Int. Arch. Allergy Immunol. 2007, 144, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Rittenour, W.R.; Hamilton, R.G.; Beezhold, D.H.; Green, B.J. Immunologic, spectrophotometric and nucleic acid based methods for the detection and quantification of airborne pollen. J. Immunol. Methods 2012, 383, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Grote, M.; Vrtala, S.; Valenta, R. Monitoring of two allergens, Bet v1 and profillin, in dry and rehydrated birch pollen by immunogold electron microscopy and immunoblotting. J. Histochem. Cytochem. 1993, 41, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lopez, J.C.; Morales, S.; Castro, A.J. Characterization of Profilin Polymorphism in Pollen with a Focus on Multifunctionality. PLoS ONE 2012, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Marth, K.; Germatuik, T.; Swoboda, I.; Valenta, R. Tree pollen allergens. In Allergens and Allergen Immunotherapy: Subcutaneous, Sublingual, and Oral, 5th ed.; Lockey, R.F., Ledford, D.K., Eds.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Rodríguez, R.; Villalba, M.; Monsalve, R.I.; Batanero, E. The spectrum of olive pollen allergens. Int. Arch. Allergy Immunol. 2001, 125, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.G.N.; Horton, P.; Gray, J.E. The Arabidopsis Cyclophilin Gene Family. Plant Physiol. 2004, 134, 1268–1282. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Roy, S.; Singh, P.; Singla-Pareek, S.; Pareek, A. Cyclophilins: Proteins in search of function. Plant Signal. Behav. 2013, 8, e22734. [Google Scholar] [CrossRef] [PubMed]
- Luttkopf, D.; Muller, U.; Skov, P.S. Comparison of four variants of a major allergen in hazelnut (Corylusavellana) Cor a 1.04 with the major hazel pollen allergen Cor a 1.01. Mol. Immunol. 2002, 38, 515–525. [Google Scholar] [CrossRef]
- Pokoj, S.; Lauer, I.; Fotisch, K. Pichiapastoris is superior to E-coli for the production of recombinant allergenic non-specific lipid-transfer proteins. Protein Expr. Purif. 2010, 69, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Hayek, B.; Vangelista, L.; Pastore, A. Molecular and immunological characterization of a highly cross-reactive two EF-hand calcium-binding alder pollen allergens, Aln g 4: Structural basis for calcium-modulated IgE recognition. J. Immunol. 1998, 161, 7031–7039. [Google Scholar]
- Crameri, R. High throughput screening: A rapid way to recombinant allergens. Allergy 2001, 56, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.S.; Shen, H.H.; Zheng, M.; Frewer, L.J.; Gilissen, W.J. Multidisciplinary Approaches to Allergies; Zhejiang University Press: Hangzhou, China, 2012. [Google Scholar]
- Breiteneder, H.; Pettenburger, K.; Bito, A. The gene coding for the major birch pollen allergen, Bet v 1, is highly homologous to a pea disease resistance response gene. EMBO J. 1989, 8, 1935–1938. [Google Scholar] [PubMed]
- Breiteneder, H.; Ferreira, F.; Hoffmann-Sommergruber, K. Four recombinant isoforms of Cora I, the major allergen of hazel pollen, show different IgE-binding properties. Eur. J. Biochem. 1993, 212, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Seiberler, S.; Scheiner, O.; Kraft, D. Characterization of a birch pollen allergen, Bet v III, representing a novel class of Ca2+ binding proteins; specific expression in mature pollen and dependence of patients’ IgE binding on protein-bound Ca2+. EMBO J. 1994, 13, 3481–3486. [Google Scholar] [PubMed]
- Twardosz, A.; Hayek, B.; Seiberler, S. Molecular characterization, expression in Escherichia coli and epitope analysis of a two EF-hand calcium-binding birch pollen allergen, Bet v 4. Biochem. Biophys. Res. Commun. 1997, 239, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Zuidmeer-Jongejan, L.; Fernández-Rivas, M.; Winter, M.G.T. Oil body-associated hazelnut allergens including oleosins are underrepresented in diagnostic extracts but associated with severe symptoms. Clin. Trans. Allergy 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Liscombe, D.K.; MacLeod, B.P.; Loukanina, N. Evidence for the monophyletic evolution of benzylisoquinoline alkaloid biosynthesis in angiosperms. Phytochemistry 2005, 66, 2501–2520. [Google Scholar] [CrossRef] [PubMed]
- Markovic-Housley, Z.; Degano, M.; Lamba, D. Crystal structure of a hypoallergenic isoform of the major birch pollen allergen Bet v 1 and its likely biological function as a plant steroid carrier. J. Mol. Biol. 2003, 325, 123–133. [Google Scholar] [CrossRef]
- Roth-Walter, F.; Gomez-Casado, C.; Pacios, L.F. Bet v 1 from birch pollen is a lipocalin-like protein acting as allergen only when devoid of iron by promoting Th2lymphocytes. J. Biol. Chem. 2014, 289, 17416–17421. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Kim, K.J.; Shin, R. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. Plant J. 2004, 37, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Osmark, P.; Boyle, B.; Brisson, N. Sequential and structural homology between intracellular pathogenesis-related proteins and a group of latex proteins. Plant Mol. Biol. 1998, 38, 1243–1246. [Google Scholar] [CrossRef] [PubMed]
- Vieths, S.; Scheurer, S.; Ballmer-Weber, B. Current understanding of cross-reactivity of food allergens and pollen. Ann. N. Y. Acad. Sci. 2002, 964, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Samanani, N.; Liscombe, D.K.; Facchini, P.J. Molecular cloning and characterization of norcoclaurine synthase, an enzyme catalyzing the first committed step in benzylisoquinoline alkaloid biosynthesis. Plant J. 2004, 40, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.; Ebner, C.; Kramer, B. Modulation of IgE reactivity of allergens by site-directed mutagenesis: Potential use of hypoallergenic variants for immunotherapy. FASEB J. 1998, 12, 231–242. [Google Scholar]
- Valenta, R.; Breiteneder, V.; Petternburger, K. Homology of the major birch-pollen allergen, Bet v I, with the major pollen allergens of alder, hazel, and hornbeam at the nucleic acid level as determined by cross-hybridization. J. Allergy Clin. Immun. 1991, 87, 677–682. [Google Scholar] [CrossRef]
- Mas, S.; Torres, M.; Garrodo-Arandia, M. Ash pollen immunoproteomics: Identification, Immunologic characterization, and sequencing of 6 new allergens. J. Allergy Clin. Immun. 2014, 133, 923–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenta, R.; Ferreira, F.; Grote, M. Identification of profilin as an actin-binding protein in higher plants. J. Biol. Chem. 1993, 268, 22777–22781. [Google Scholar] [PubMed]
- Ren, H.; Xiang, Y. The function of actin-binding proteins in pollen tube growth. Protoplasma 2007, 230, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. Plant profiling isovariants are distinctly regulated in vegetative and reproductive tissues. Cell Motil. Cytoskelet. 2002, 52, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Kovar, D.R.; Drøbak, B.K.; Staiger, C.J. Maize profilin isoforms are functionally distinct. Plant Cell 2000, 12, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Lauer, I.; Alessandri, S.; Pokoj, S. Expression and characterization of three important panallergens from hazelnut. Mol. Nutr. Food Res. 2008, 52, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Garino, C.; Locatelli, M.; Coïsson, J.D. Gene transcription analysis of hazelnut (Corylusavellana L.) allergens Cor a 1, Cora 8 and Cor a 11: A comparative study. Int. J. Food Sci. Technol. 2013, 48, 1208–1217. [Google Scholar] [CrossRef]
- Longhi, S.; Cristofori, A.; Gatto, P. Biomolecular identification of allergenic pollen: A new perspective for aerobiological monitoring? Ann. Allergy Asthma Immunol. 2009, 103, 508–514. [Google Scholar] [CrossRef]
- Aguerri, M.; Calzada, D.; Montaner, D. Differential gene-expression analysis defines a molecular pattern related to olive pollen allergy. J. Biol. Regul. Homeost. Agents 2013, 27, 337–350. [Google Scholar] [PubMed]
- Žiarovská, J.; Labajová, M.; Ražná, K. Changes in expression of BetV1 allergen of silver birch pollen in urbanized area of Ukraine. J. Environ. Sci. Health 2013, 48, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Ražná, K.; Bežo, M.; Nikolaieva, N. Variability of Corylus avellana, L. CorA and profilin pollen allergens expression. J. Environ. Sci. Health 2014, 49, 639–645. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Liccardi, G.; D’Amato, M.; Holgatew, S. Environmental risk factors and allergic bronchial asthma. Clin. Exp. Allergy 2005, 35, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, T.; Thomas, S. Gene expression and characterization of a stress-induced tyrosine decarboxylase from Arabidopsis thaliana. FEBS Lett. 2009, 583, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Pochop, J.; Kačániová, M.; Hleba, L. Detection of Listeria monocytogenes in ready-to-eat food by step one real-time polymerase chain reaction. J. Environ. Sci. Health B 2012, 47, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Farrokhi, R.; Joozani, R.J. Identification of pork genome in commercial meat extracts for Halal authentication by SYBR green I real-time PCR. Int. J. Food Sci. Technol. 2011, 46, 951–955. [Google Scholar] [CrossRef]
- Kang, I.H.; Srivastava, P.; Ozias-Akins, P.; Gallo, M. Temporal and Spatial Expression of the Major Allergens in Developing and Germinating Peanut Seed. Plant Physiol. 2007, 144, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Platteau, C.; De Loose, M.; De Meulenaer, B.; Taverniers, I. Detection of allergenic ingredients using real-time PCR: A case study on hazelnut (Corylus avellena) and soy (Glycine max). J. Agric. Food Chem. 2011, 59, 10803–10814. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.; Amin, Z. Airborne Pollen Allergy—Impact on Human Health; Lambert Academic Publishing: Saarbrücken, Germany, 2012. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xie, L.; Chen, J. A novel procedure for absolute real-time quantification of gene expression patterns. Plant Methods 2012, 8, 9. [Google Scholar] [CrossRef] [PubMed]
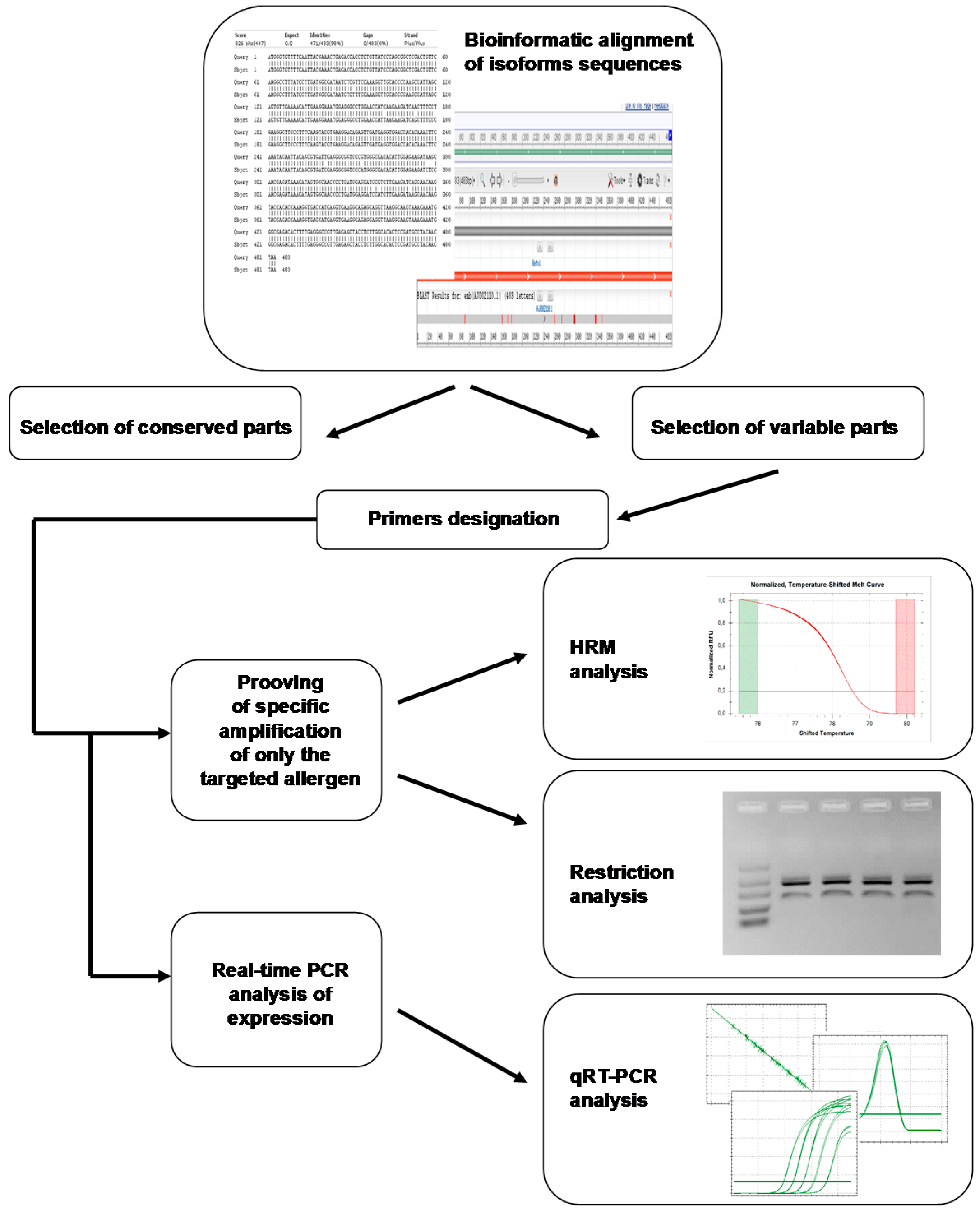
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; D’Auria, J.C.; Luck, K.; Gershenzon, J. Evaluation of Candidate Reference Genes for Real-Time Quantitative PCR of Plant Samples Using Purified cDNA as Template. Plant Mol. Biol. Rep. 2009, 27, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mauriat, M.; Guenin, S. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.F.; Sun, S.S.M.; Yuan, D.Y. Validation of Candidate Reference Genes for the accurate normalization of Real-Time Quantitative RT-PCR Data in Rice during seed development. Plant Mol. Biol. Rep. 2010, 28, 49–57. [Google Scholar] [CrossRef]
- Nicot, N.; Hausman, J.F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Sahoo, A.; Tyagi, A.K.; Jain, M. Validation of internal control genes for quantitative gene expression studies in chickpea (Cicer arietinum L.). Biochem. Biophys. Res. Commun. 2010, 396, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Žiarovská, J.; Nikolajeva, N.; Garkava, K.; Brindza, J. Validation of HMG CoA reductase as internal control for hazelnut pollen allergens expression analysis. Austin J. Genet. Genom. Res. 2015, 2, 1–4. [Google Scholar]




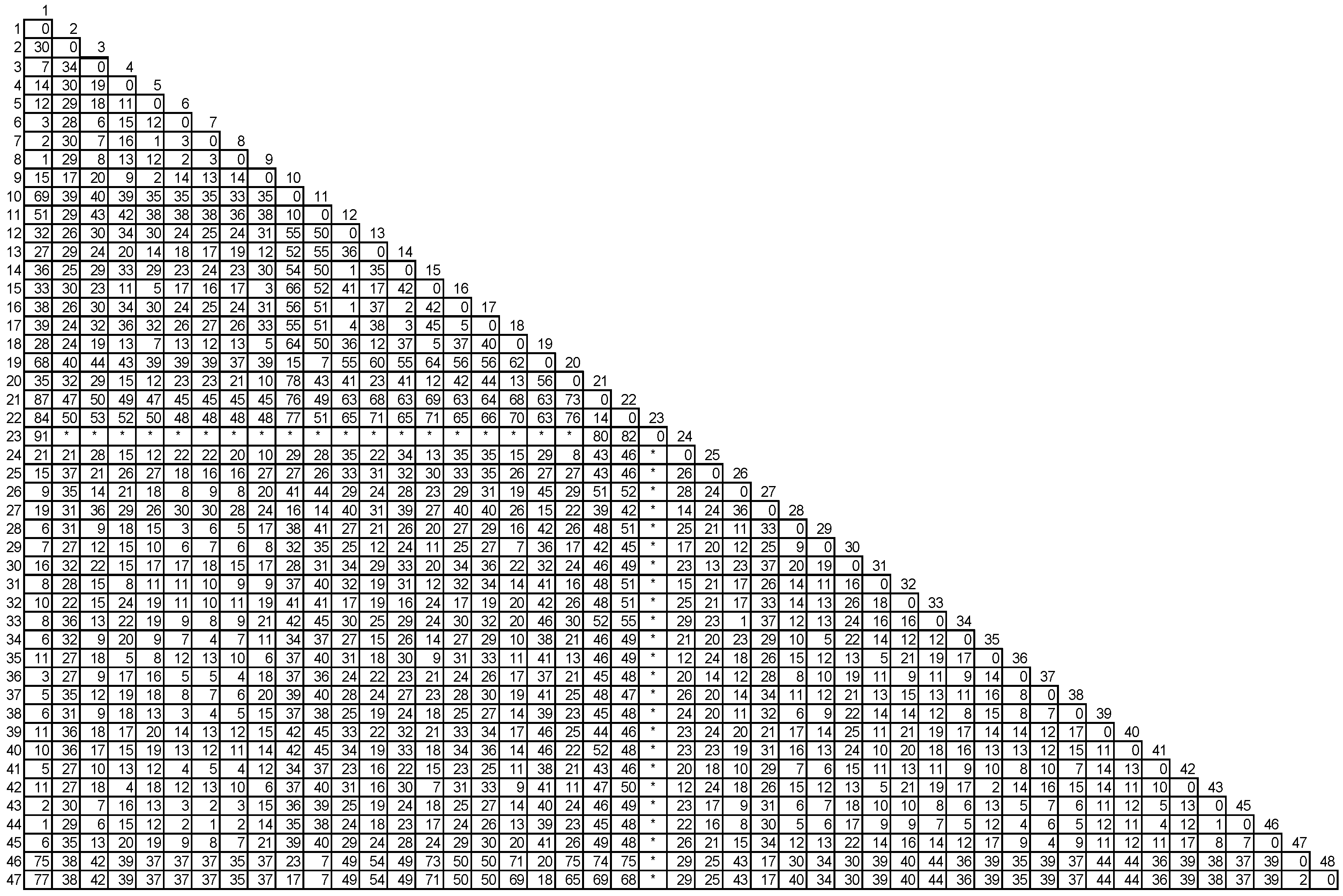{kind=link}
{kind=link}
{kind=link}
| Allergen Type/Number | Occurrence of Homologs | Biological Function |
|---|---|---|
| Bet v1 type | alder, hazel 1, birch | pathogenesis related protein [8] |
| Profilin (Bet v2 type) | hazel, birch | actin-binding protein [8] |
| EF hands type | alder, birch | Ca2+-binding protein [8] |
| olive pollen allergen | ash, London plane tree | unknown [9] |
| cyclophilin | birch, oriental plane | immunosuppressant receptors–immunophilins [9,10,11] |
| lipid transfer protein | Hazel 2, London plane tree, oriental plane | lipid transport [8] |
| Species | Pollen Allergen * | Nucleotide Accession Code | Type of Nucleic Acid | Function or Similarity * |
|---|---|---|---|---|
| Alnus glutinosa | Aln g 1 | S50892.1 | mRNA | PR10; Bet v1 type |
| Aln g 4 | Y17713.1 | mRNA | Ca2+-binding protein | |
| Corylus avellana | Cor a 1 a | Z72440.1Z72439.1 | DNA | PR 10; Bet v1 type |
| Cor a 2 | AF327623.1AF327622.1 | mRNA | profilin; Bet v2 type | |
| Cor a 8 b | AF329829.1 | mRNA | lipid transfer protein precursor | |
| Cor a 9 b,c | KF494372 | mRNA | 11S globulin (legumin-like) | |
| Cor a 10 | AJ295617.1 | mRNA | luminal binding protein | |
| Cor a 11 b | AF441864.1 | mRNA | 11S globulin (vicilin-like) | |
| Cor a 12 | AY224599 | mRNA | oleosin | |
| Cor a 13 | ||||
| Cor a 14 b | FJ358504 | mRNA | 2S albumin | |
| Fraxinus excelsior | Fra e 1 d | AY377127.1 | mRNA | Ole e 1 related (glycosylated protein) |
| Fra e 2 | KC920922.1 | mRNA | profilin | |
| Fra e 3 | KC920923.1 | mRNA | Ca binding protein | |
| Fra e 6 | KC920921.1 | mRNA | not identified | |
| Fra e 9 | KC920916.1 | mRNA | not identified | |
| Fra e 10 | KC920924.1 | mRNA | not identified | |
| Fra e 11 | KC920915.1 | mRNA | not identified | |
| Fra e 12 | EF626802.1 | mRNA | not identified | |
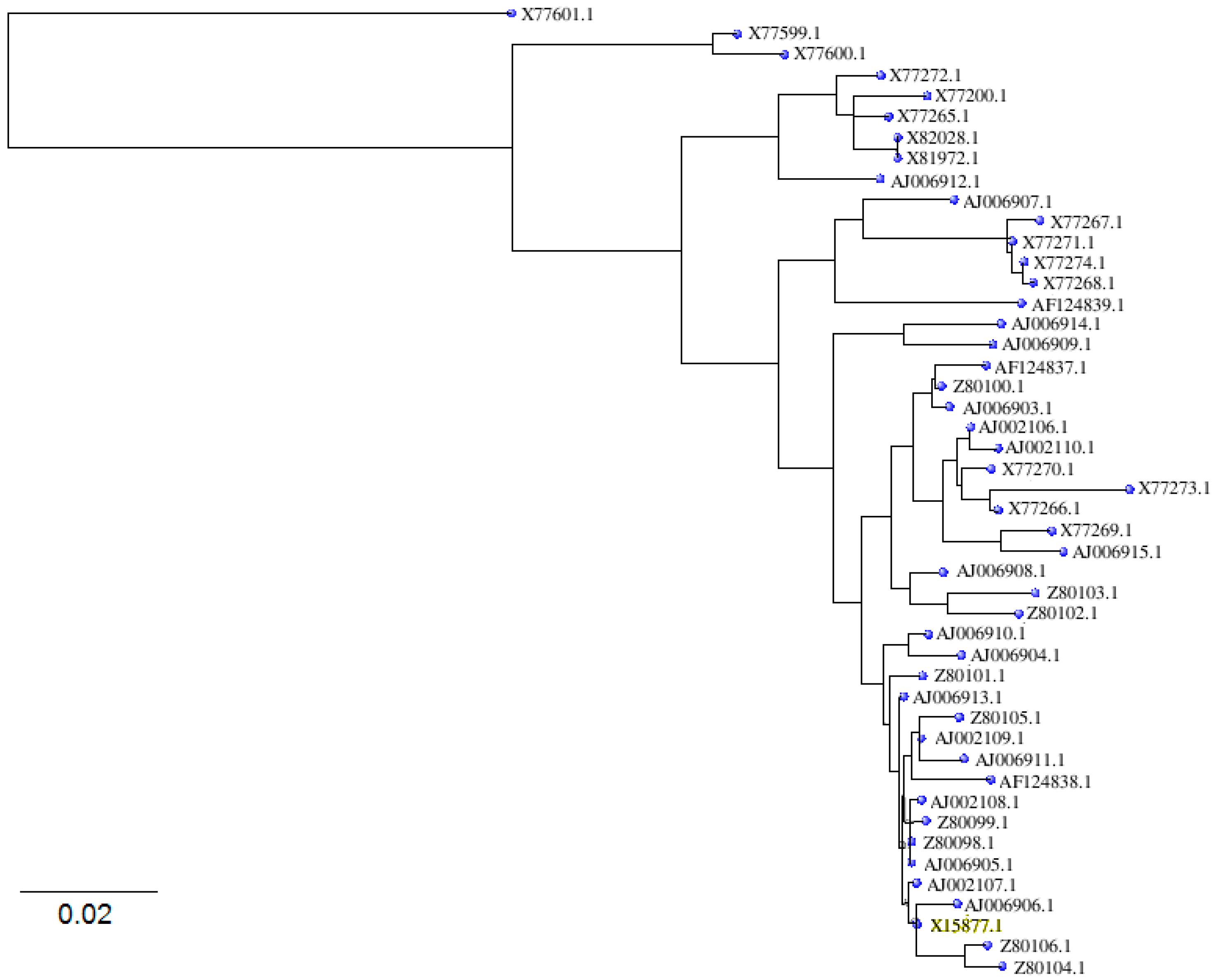
| Betula verrucosa | Bet v1 e | X15877.1 | mRNA | PR10 |
| Bet v2 | M65179 | mRNA | profilin | |
| Bet v3 | X79267 | mRNA | Ca2+-binding protein | |
| Bet v4 | X87153 | mRNA | Ca2+-binding protein | |
| Bet v5 | - | - | isoflavonereductase | |
| Bet v6 | AF135127AF282850 | mRNA | isoflavonereductase | |
| Bet v7 | AJ311666.1 | mRNA | cyclophilin | |
| Bet v8 | - | - | pectin esterase | |
| Platanus acerifolia | Pla a 1 | AJ427413.2 | mRNA | invertase inhibitor |
| Pla a 2 | AJ586898.1 | mRNA | polymethylgalacturonase | |
| Pla a 3 | - | - | Lipid transfer protein | |
| Pla a 8 | ||||
| Ole e 1 | KM397755.1 | mRNA | pollen allergen | |
| Platanus orientalis | Pla or 1 | EU296476.1 | mRNA | plant invertase/pectin methylesterase inhibitor (cyclophilin) |
| Pla or 2 | EU296477 | mRNA | polygalacturonase | |
| Pla or 3 | EU296478 | mRNA | lipid transfer protein precursor |
| Isoform/Nucleotide Position * | 1 | 2 | 3 | 27 | 27 | 33 | 34 | 36 | 37 | 55 | 73 | 85 | 96 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AY652744.1 | G | A | G | T | T | C | T | C | G | T | G | G | A |
| AY377127.1 | x | x | x | C | A | T | C | T | A | C | C | A | G |
| isoform/nucleotide position * | 102 | 103 | 104 | 108 | 112 | 131 | 135 | 148 | 150 | 165 | 166 | 182 | 203 |
| AY652744.1 | T | C | A | C | G | A | G | A | A | A | G | C | G |
| AY377127.1 | A | A | C | T | A | G | C | G | C | G | A | G | T |
| nucleotide position * | 206 | 208 | 214 | 235 | 240 | 241 | 242 | 243 | 244 | 255 | 256 | 264 | 271 |
| AY652744.1 | A | T | C | C | T | G | T | A | C | C | A | A | C |
| AY377127.1 | G | G | T | T | C | A | C | G | T | T | G | G | A |
| isoform/nucleotide position * | 284 | 285 | 300 | 306 | 309 | 324 | 339 | 348 | 391 | 397 | 409 | 416 | - |
| AY652744.1 | C | C | G | G | G | C | C | G | A | C | T | T | - |
| AY377127.1 | T | T | A | A | A | T | A | C | C | G | C | C | - |
| Group No. | Accession Code for Cor a 1 | Type of Nucleic Acid | Sequences Similarity within the Group |
|---|---|---|---|
| 1 | AF323975.1 | mRNA | 99% |
| AF323974.1 | |||
| AF323973.1 | |||
| AF136945.1 | |||
| 2 | Z72440.1 | DNA | 75% |
| Z72439.1 | |||
| 3 | X71000.1 | mRNA | 96%–99% |
| X70999.1 | |||
| X70998.1 | |||
| X70997.1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Žiarovská, J.; Zeleňáková, L. Central and Eastern European Spring Pollen Allergens and Their Expression Analysis—State of the Art. Diversity 2016, 8, 19. https://doi.org/10.3390/d8040019
Žiarovská J, Zeleňáková L. Central and Eastern European Spring Pollen Allergens and Their Expression Analysis—State of the Art. Diversity. 2016; 8(4):19. https://doi.org/10.3390/d8040019
Chicago/Turabian StyleŽiarovská, Jana, and Lucia Zeleňáková. 2016. "Central and Eastern European Spring Pollen Allergens and Their Expression Analysis—State of the Art" Diversity 8, no. 4: 19. https://doi.org/10.3390/d8040019
APA StyleŽiarovská, J., & Zeleňáková, L. (2016). Central and Eastern European Spring Pollen Allergens and Their Expression Analysis—State of the Art. Diversity, 8(4), 19. https://doi.org/10.3390/d8040019