
The Chemistry Behind ADCs


Abstract
:1. Introduction
2. ADC Payloads and Their Attachment to the Linker
2.1. Microtubule-Disrupting Drugs
2.1.1. Auristatins
2.1.2. Maytansinoid Derivatives (DM2, DM4)
2.1.3. Tubulysins
2.1.4. Cryptomycins
2.1.5. Antimitotic EG5 Inhibitors
2.2. DNA Damaging Drugs
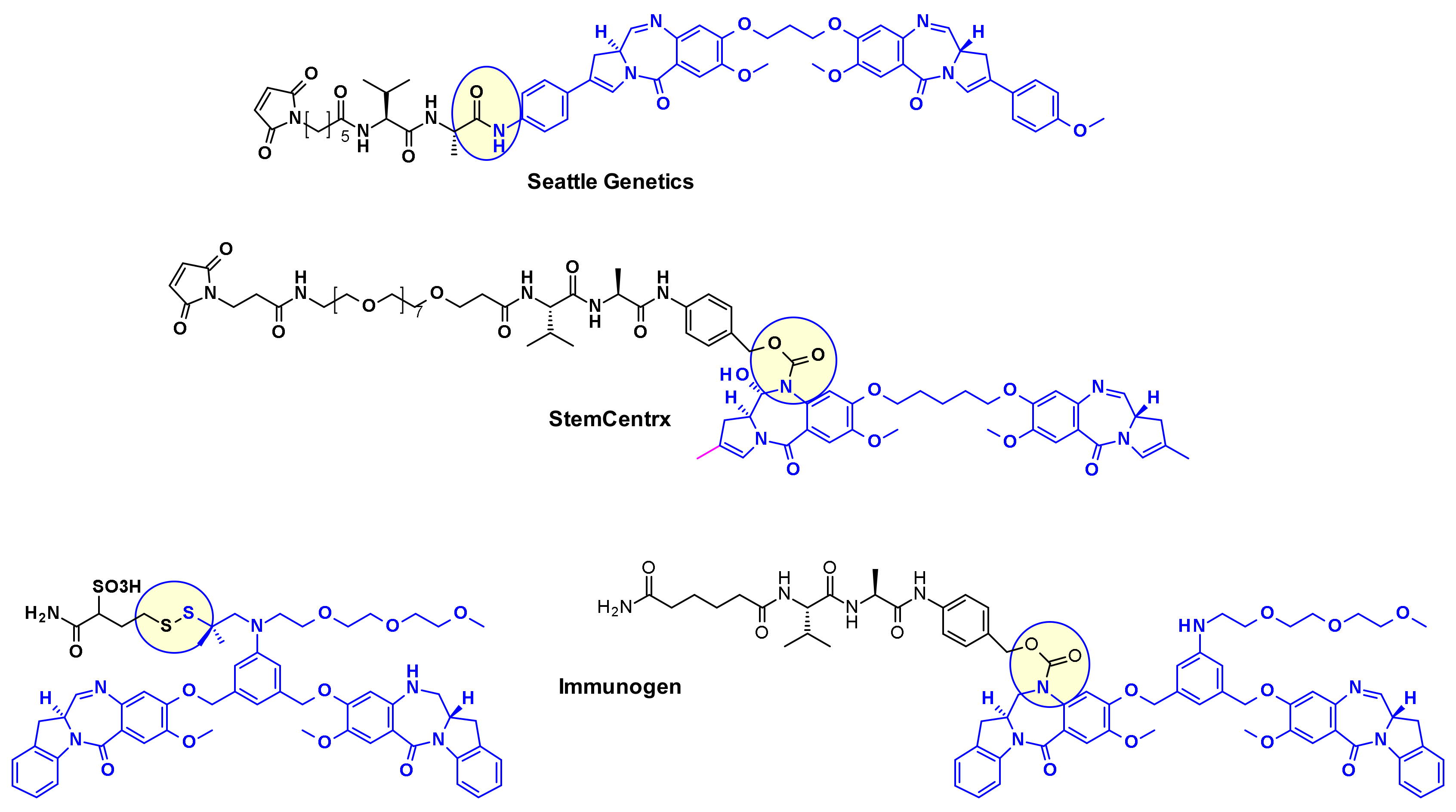
2.2.1. Pyrrolobenzodiazepines and Indolinobenzodiazepine
2.2.2. Duocarmycins
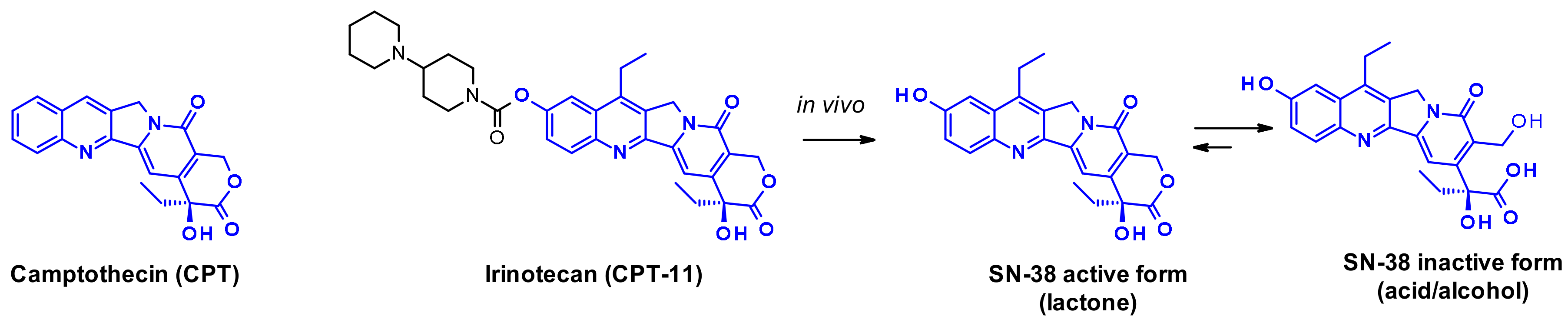
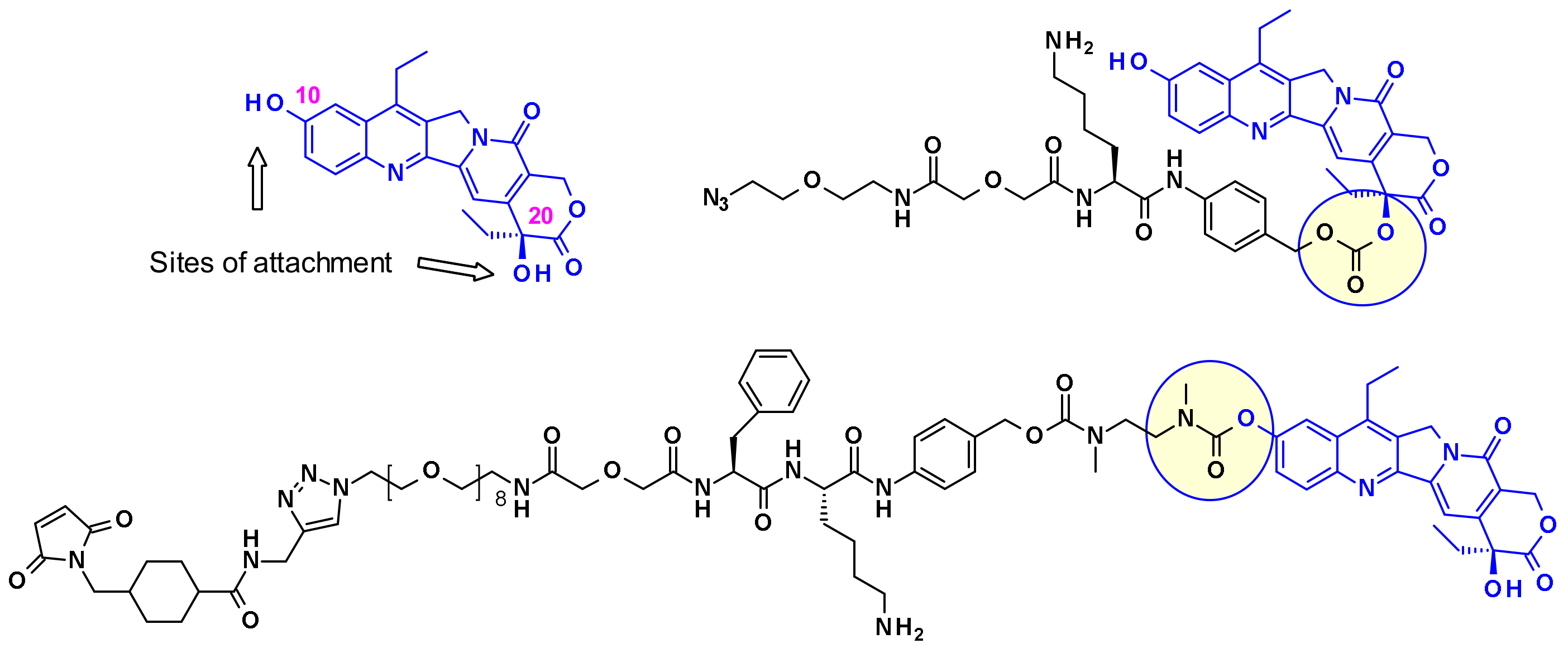
2.2.3. Camptothecin
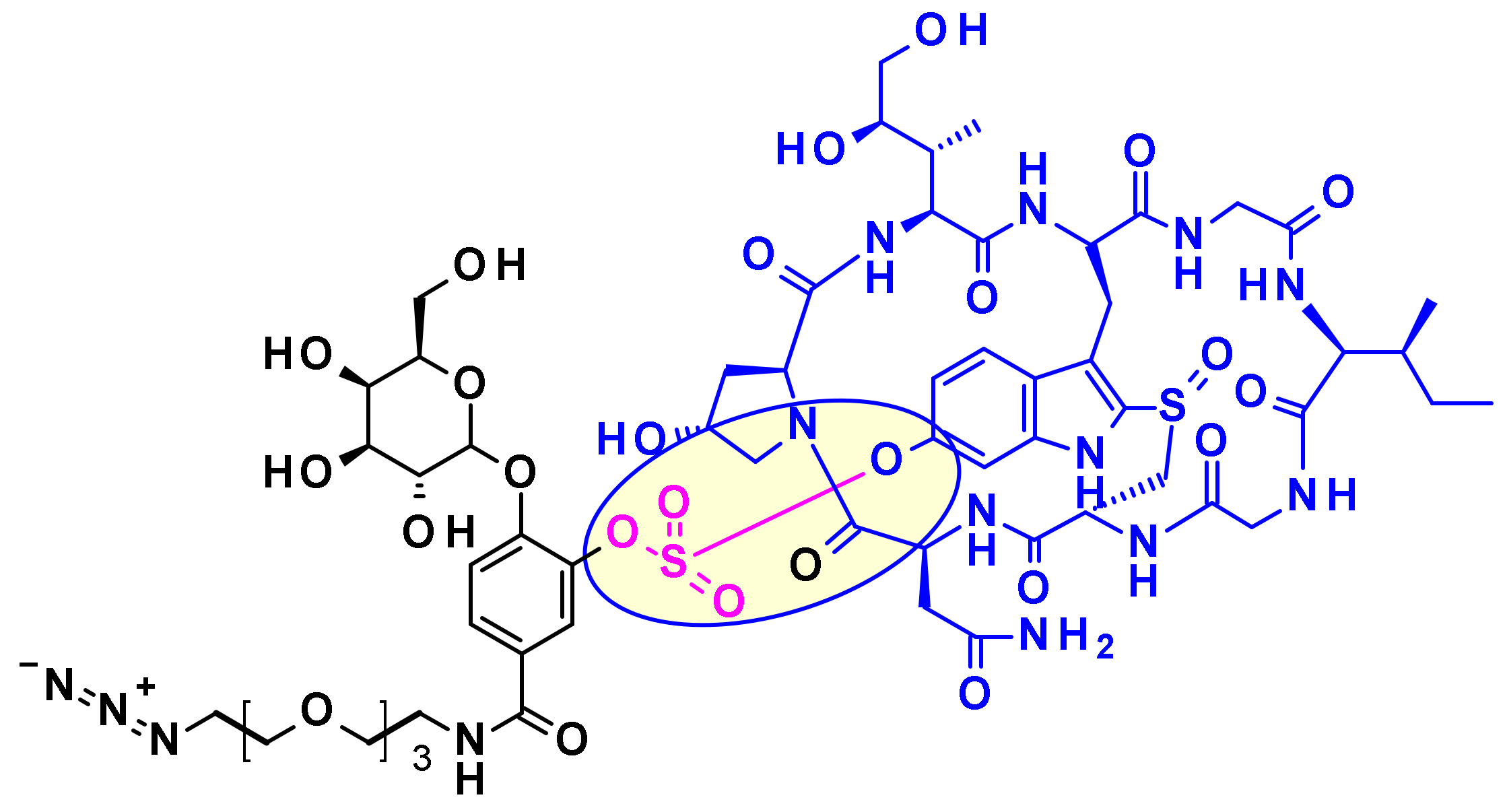
2.2.4. Calicheamicin
2.3. Innovative Drugs
2.3.1. Apoptosis Inducers (Bcl-xL Inhibitors)
2.3.2. Thailanstatin and Analogues
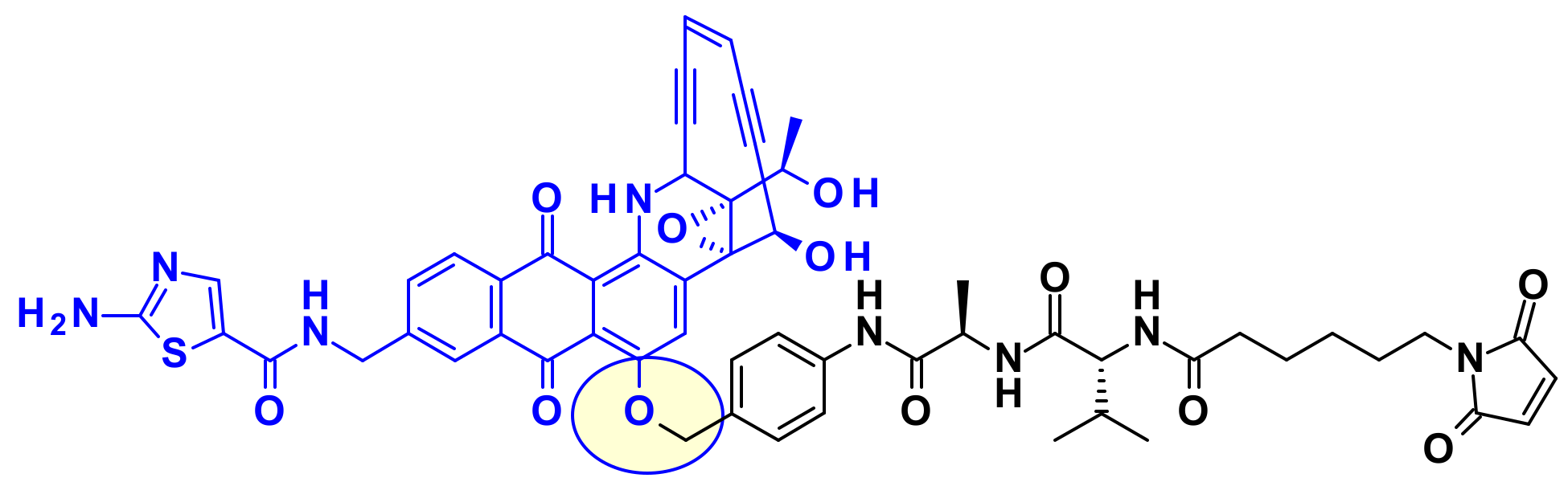
2.3.3. Amatoxins
2.3.4. Inhibition of the Nicotinamide Phosphoribosyltransferase (NAMPT)
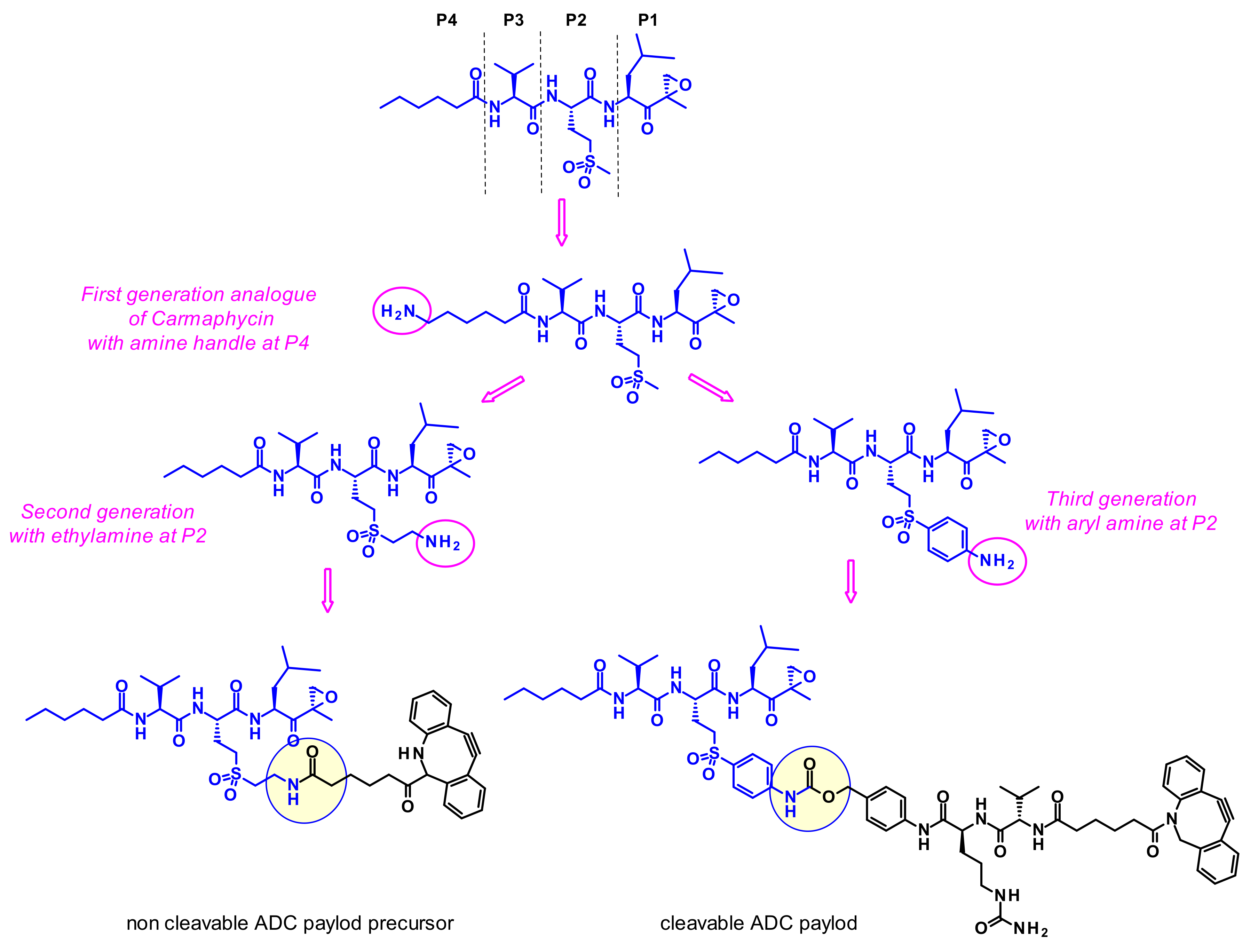
2.3.5. Carmaphycins
3. Linker Types
3.1. Non-Cleavable Linkers
3.2. Cleavable Linkers
3.2.1. Non-Enzymatic Linkers
3.2.2. Enzymatic Cleavage
Cathepsin-B
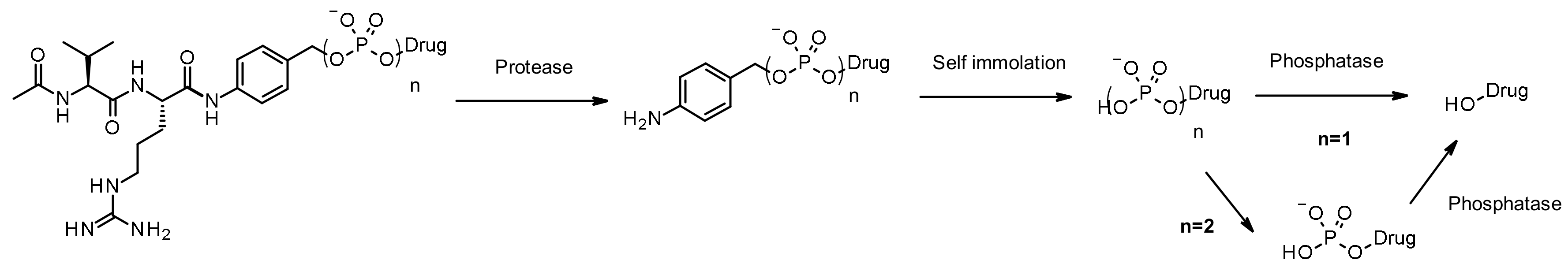
Phosphatase and Pyrophosphatase
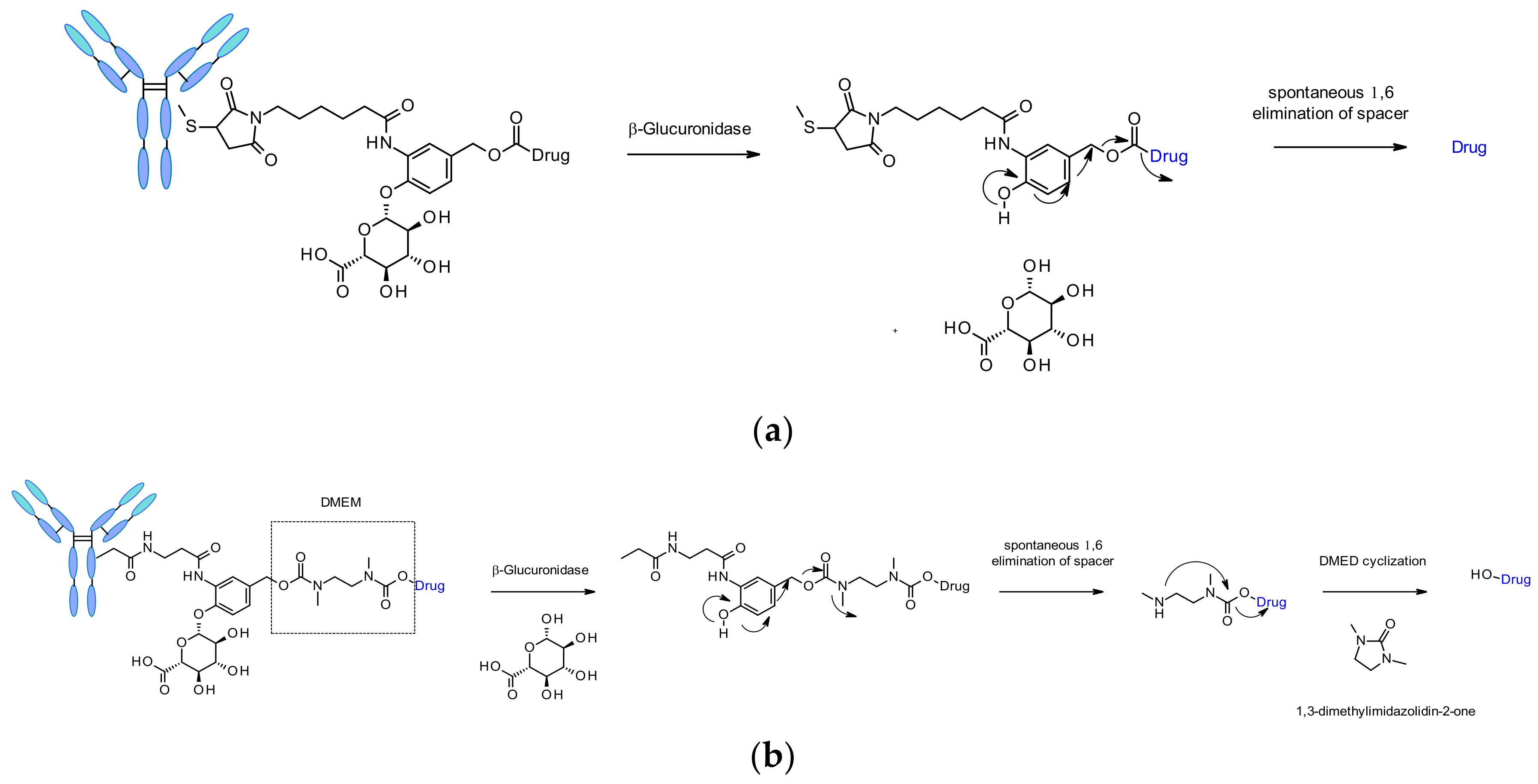
β-Glucuronidases
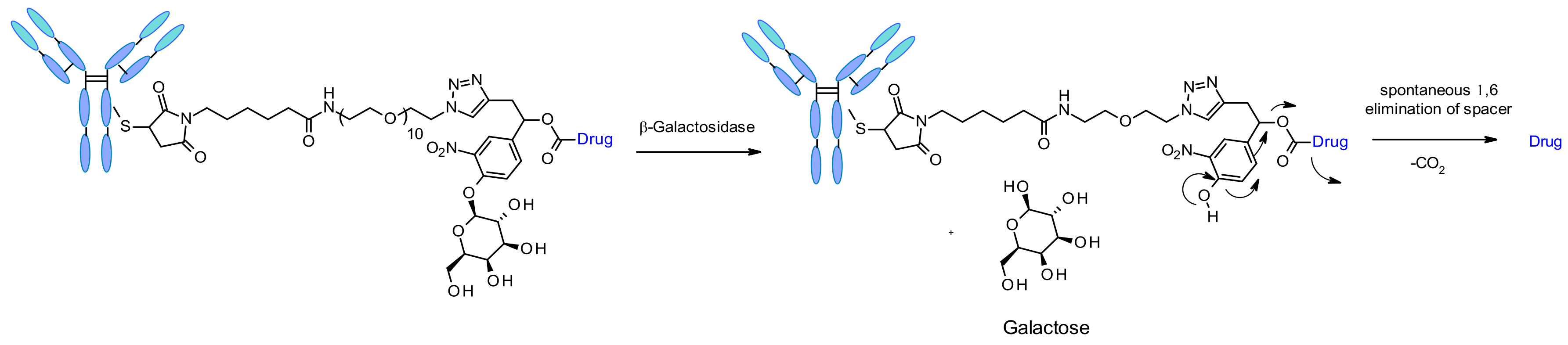
β-Galactosidase
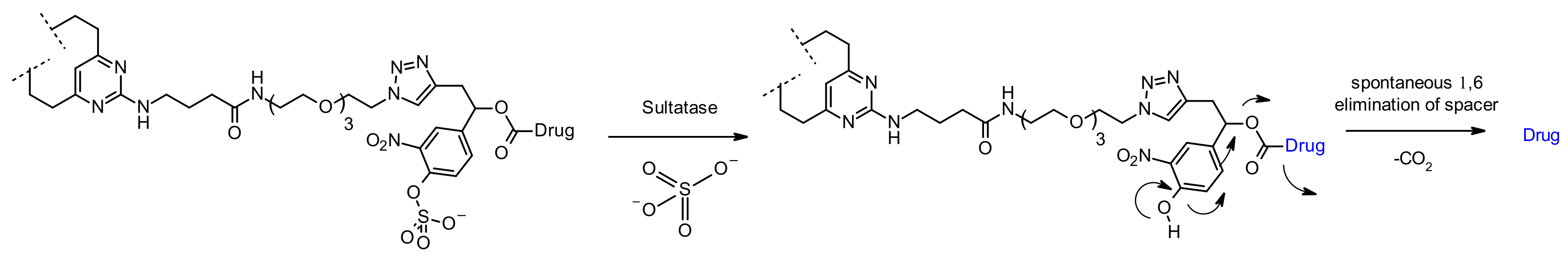
Sulfatase
4. Bioconjugation
4.1. Chemistry Based Site-Specific Modification of Native Antibodies
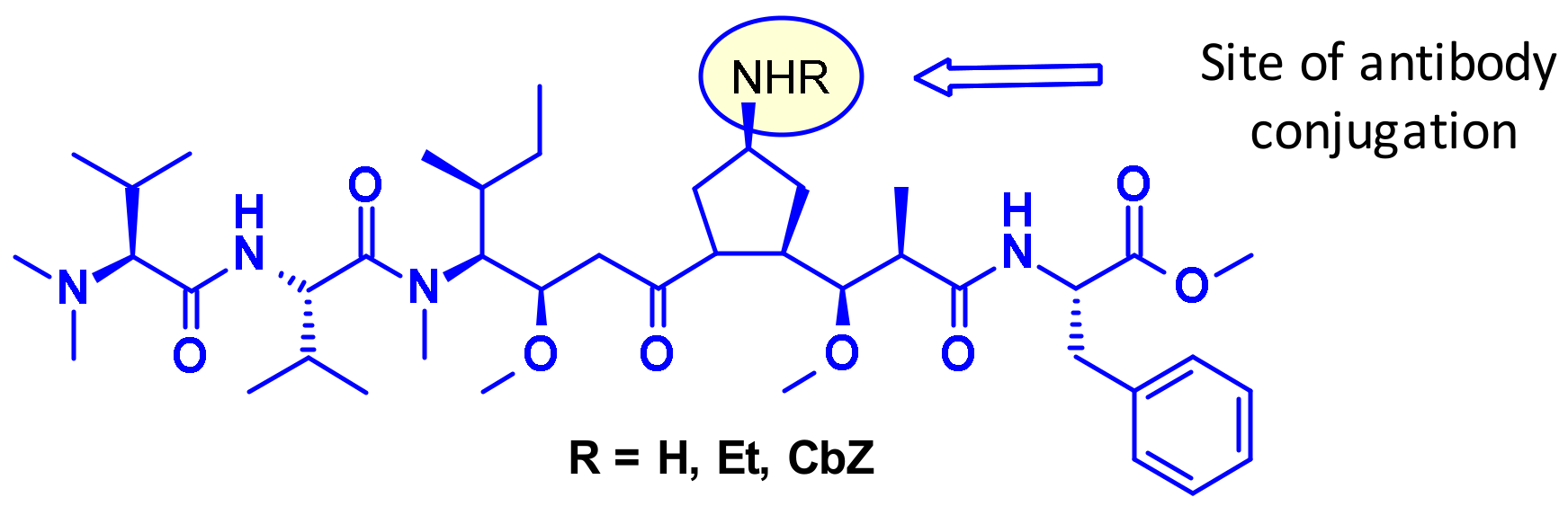
4.1.1. Conjugation to Endogenous Amino Acids
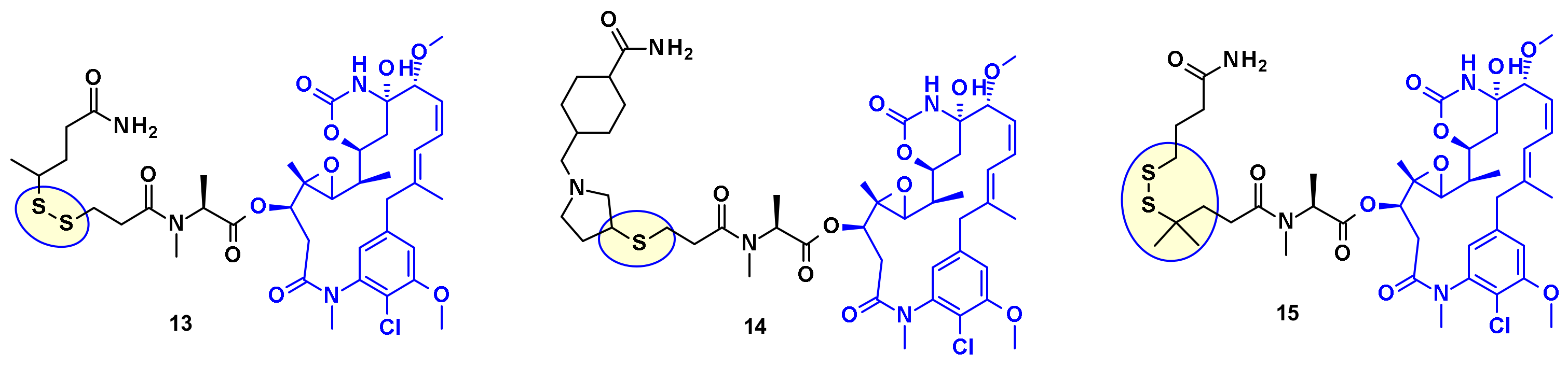
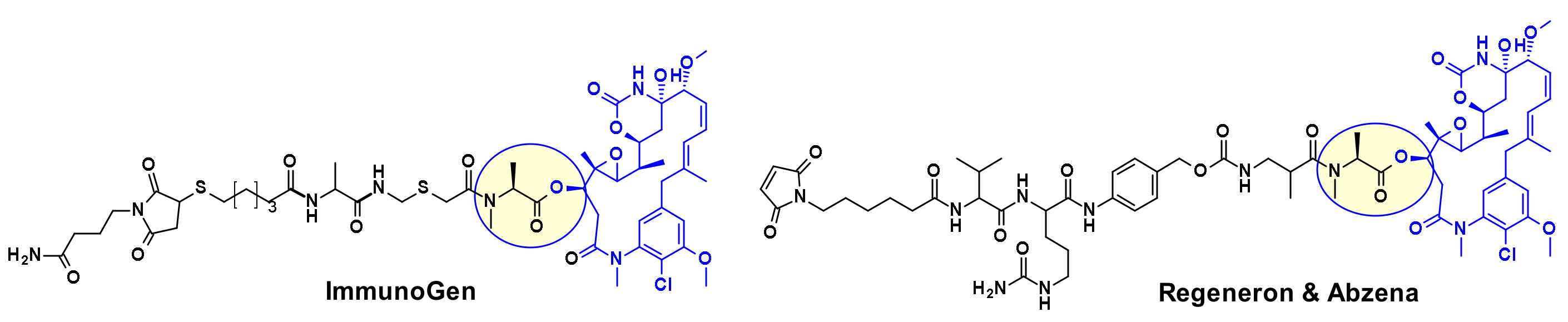
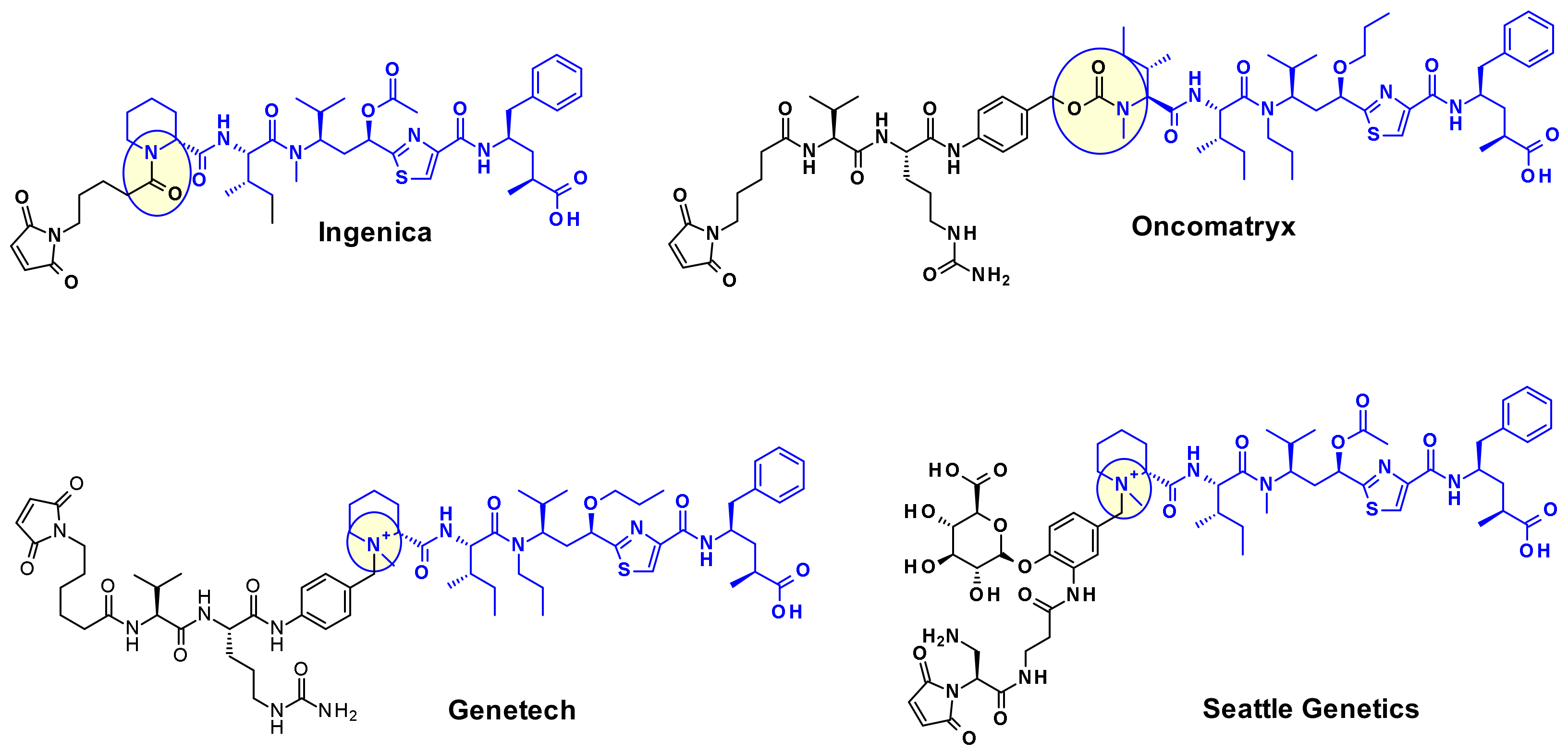
4.1.2. Disulphide Rebridging Strategies
4.1.3. Conjugation to Glycan
4.2. Site-Specific Bioconjugation of Engineered Antibodies
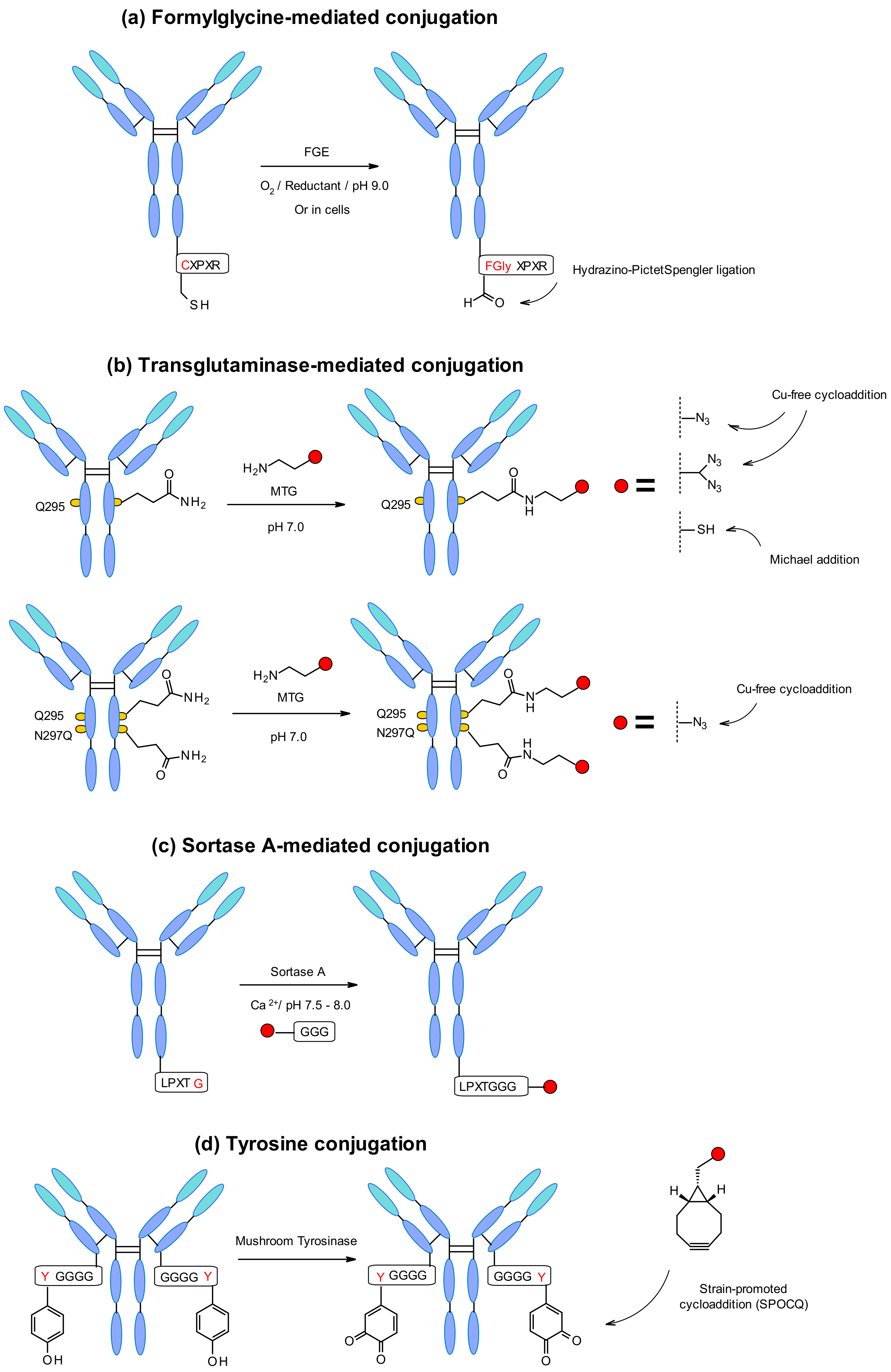
4.2.1. Enzymatic Approaches
4.2.2. Cysteine Engineering: Thiomab Technology
4.2.3. Conjugation to Engineered Unnatural Amino Acids/SelenomAb
5. Overview of the ADCs in Late-Stage Clinical Development
6. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Drago, J.Z.; Modi, S.; Chandarlapaty, S. Unlocking the potential of antibody–drug conjugates for cancer therapy. Nat. Rev. Clin. Oncol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Khongorzul, P.; Ling, C.J.; Khan, F.U.; Ihsan, A.U.; Zhang, J. Antibody–Drug Conjugates: A Comprehensive Review. Mol. Cancer Res. 2020, 18, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joubert, N.; Beck, A.; Dumontet, C.; Denevault-Sabourin, C. Antibody–Drug Conjugates: The Last Decade. Pharmaceuticals 2020, 13, 245. [Google Scholar] [CrossRef]
- Ritchie, M.; Tchistiakova, L.; Scott, N. Implications of receptor-mediated endocytosis and intracellular trafficking dynamics in the development of antibody drug conjugates. mAbs 2013, 5, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, M.J.; Lindsay, A.J. The Endosomal Recycling Pathway—At the Crossroads of the Cell. Int. J. Mol. Sci. 2020, 21, 6074. [Google Scholar] [CrossRef] [PubMed]
- Kaplon, H.; Reichert, J.M. Antibodies to watch in 2019. mAbs 2019, 11, 219–238. [Google Scholar] [CrossRef] [PubMed]
- Maderna, A.; Leverett, C.A. Recent advances in the development of new auristatins: Structural modifications and application in antibody drug conjugates. Mol. Pharm. 2015, 12, 1798–1812. [Google Scholar] [CrossRef] [PubMed]
- Burke, P.J.; Hamilton, J.Z.; Pires, T.A.; Setter, J.R.; Hunter, J.H.; Cochran, J.H.; Waight, A.B.; Gordon, K.A.; Toki, B.E.; Emmerton, K.K.; et al. Development of Novel Quaternary Ammonium Linkers for Antibody-Drug Conjugates. Mol. Cancer Ther. 2016, 15, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Akaiwa, M.; Dugal-Tessier, J.; Mendelsohn, B.A. Antibody-Drug Conjugate Payloads; Study of Auristatin Derivatives. Chem. Pharm. Bull. 2020, 68, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
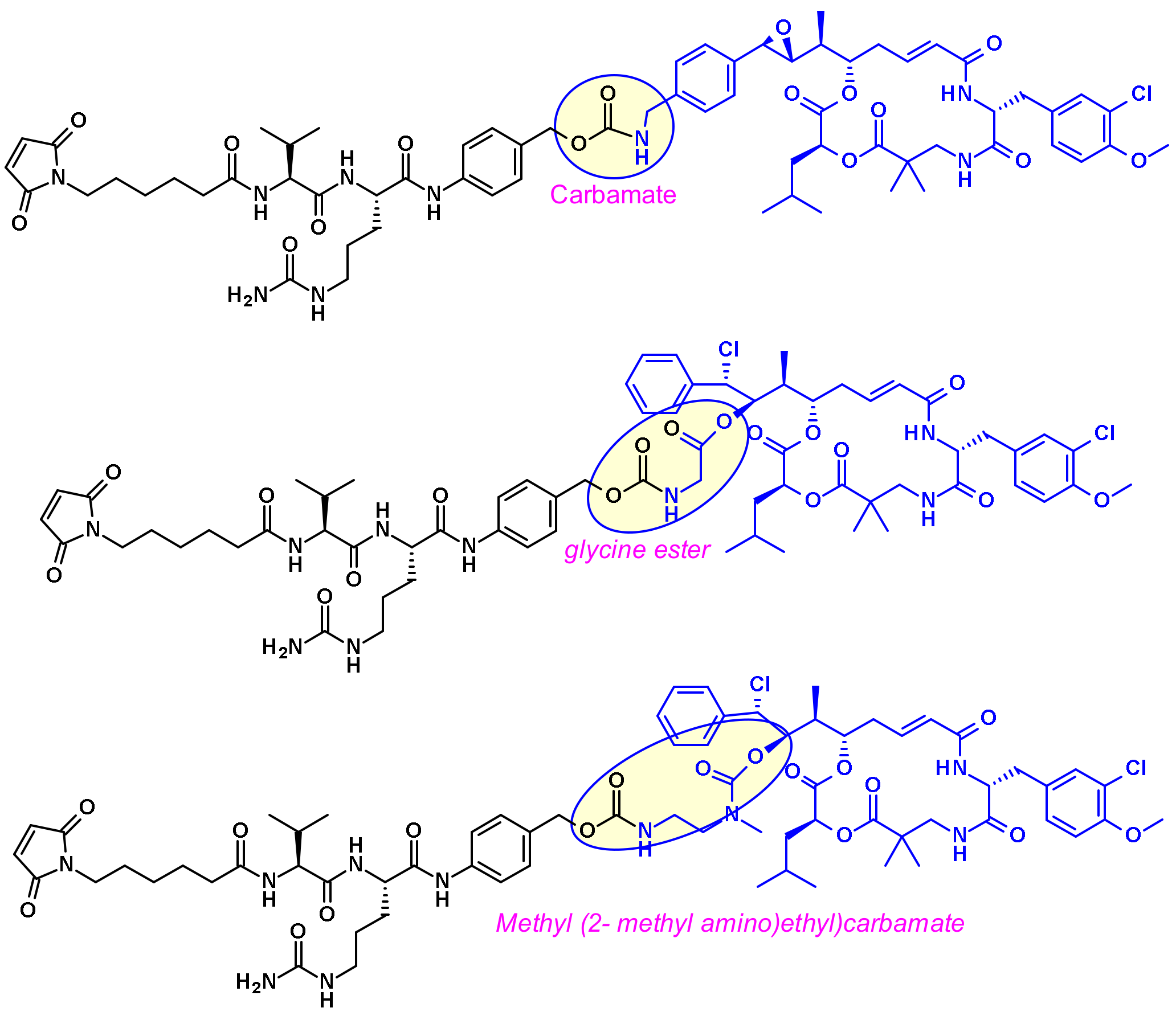
- Kolakowski, R.V.; Haelsig, K.T.; Emmerton, K.K.; Leiske, C.I.; Miyamoto, J.B.; Cochran, J.H.; Lyon, R.P.; Senter, P.D.; Jeffrey, S.C. The Methylene Alkoxy Carbamate Self-Immolative Unit: Utilization for the Targeted Delivery of Alcohol-Containing Payloads with Antibody-Drug Conjugates. Angew. Chem. Int. Ed. Engl. 2016, 55, 7948–7951. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.W.; Fahrner, R.; Shevshenko, D.; Fyrknäs, M.; Larsson, R.; Lehmann, F.; Odell, L.R. Rational Design of Azastatin as a Potential ADC Payload with Reduced Bystander Killing. ChemMedChem 2020, 15, 2500–2512. [Google Scholar] [CrossRef]
- Cassady, J.M.; Chan, K.K.; Floss, H.G.; Leistner, E. Recent developments in the maytansinoid antitumor agents. Chem. Pharm. Bull. 2004, 52, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, C.; Yang, X.; Hou, X.; Zhao, W.; Benson, D.; Yu, J.; Dong, Y. Design, synthesis and evaluation of anti-CD38 antibody drug conjugate based on Daratumumab and maytansinoid. Bioorg. Med. Chem. 2019, 27, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Costoplus, J.A.; Veale, K.H.; Qiu, Q.; Ponte, J.F.; Lanieri, L.; Setiady, Y.; Dong, L.; Skaletskaya, A.; Bartle, L.M.; Salomon, P.; et al. Peptide-Cleavable Self-immolative Maytansinoid Antibody-Drug Conjugates Designed To Provide Improved Bystander Killing. ACS Med. Chem. Lett. 2019, 10, 1393–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nittoli, T.; Kelly, M.P.; Delfino, F.; Rudge, J.; Kunz, A.; Markotan, T.; Spink, J.; Chen, Z.; Shan, J.; Navarro, E.; et al. Antibody drug conjugates of cleavable amino-alkyl and aryl maytansinoids. Bioorg. Med. Chem. 2018, 26, 2271–2279. [Google Scholar] [CrossRef]
- Kaur, G.; Hollingshead, M.; Holbeck, S.; Schauer-Vukasinović, V.; Camalier, R.F.; Dömling, A.; Agarwal, S. Biological evaluation of tubulysin A: A potential anticancer and antiangiogenic natural product. Biochem. J. 2006, 396, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.S. Tubulysins as Antibody–Drug Conjugate (ADC) Payloads. In Cytotoxic Payloads for Antibody–Drug Conjugates; Thurston, D.E., Jackson, P.J.M., Eds.; Royal Society of Chemistry: Cambridge, UK, 2019; Chapter 18; pp. 380–397. ISBN 978-1-78801-077-1. [Google Scholar]
- Morris, M.J.; Vogelzang, N.J.; Sartor, O.; Armour, A.; Groaning, M.; Robarts, A.; Petrylak, D.P.; Tolcher, A.W.; Gordon, M.S.; Babiker, H.M. Phase 1 study of the PSMA-targeted small-molecule drug conjugate EC1169 in patients with metastatic castrate-resistant prostate cancer (mCRPC). J. Clin. Oncol. 2017, 35, 5038. [Google Scholar] [CrossRef]
- Kontermann, R.; Pfizenmaier, K.; Ferrer, C.; Fabre, M.; Simon, L. Antibody-Drug Conjugates and Immunotoxins. Patent No. WO 2015118031, 13 August 2015. [Google Scholar]
- Gingipalli, L.; Toader, D.; Wang, F. Tubulysin Derivatives. Patent No. WO 2015157594, 15 October 2015. [Google Scholar]
- Rao-Naik, C.; Deshpande, S.; Antibodies, S. Conjugatables by Transglutaminase and Conjugates Made TherefromA. Patent No. WO 2016144608, 15 September 2016. [Google Scholar]
- Tumey, L.N.; Leverett, C.A.; Vetelino, B.; Li, F.; Rago, B.; Han, X.; Loganzo, F.; Musto, S.; Bai, G.; Sukuru, S.C.K.; et al. Optimization of Tubulysin Antibody-Drug Conjugates: A Case Study in Addressing ADC Metabolism. ACS Med. Chem. Lett. 2016, 7, 977–982. [Google Scholar] [CrossRef]
- Cheng, H.; Cong, Q.; Dervin, D.; Stevens, A.; Vemuri, K.; Huber, M.; Juliano, J.; Cuison, S.; Sung, J.; Passmore, D.; et al. Synthesis and Biological Evaluation of a Carbamate-Containing Tubulysin Antibody-Drug Conjugate. Bioconjug. Chem. 2020, 31, 2350–2361. [Google Scholar] [CrossRef]
- Jackson, D.Y.; Ha, E. Antibody-Drug Conjugates and Related Compounds, Compositions, and Methods. Patent No. WO 2013085925, 13 June 2013. [Google Scholar]
- Jackson, D.Y.; Ha, E. Novel Linkers for Antibody-drug Conjugates and Related Compounds, Compositions, and Methods of Use. Patent No. WO 2014197854, 11 December 2014. [Google Scholar]
- Jackson, D.Y.; Ha, E. Antibody-Drug Conjugates and Related Compounds, Compositions, and Methods. US Patent No. 20160303247, 20 October 2016. [Google Scholar]
- Staben, L.R.; Koenig, S.G.; Lehar, S.M.; Vandlen, R.; Zhang, D.; Chuh, J.; Yu, S.-F.; Ng, C.; Guo, J.; Liu, Y.; et al. Targeted drug delivery through the traceless release of tertiary and heteroaryl amines from antibody-drug conjugates. Nat. Chem. 2016, 8, 1112–1119. [Google Scholar] [CrossRef]
- Verma, V.A.; Pillow, T.H.; DePalatis, L.; Li, G.; Phillips, G.L.; Polson, A.G.; Raab, H.E.; Spencer, S.; Zheng, B. The cryptophycins as potent payloads for antibody drug conjugates. Bioorg. Med. Chem. Lett. 2015, 25, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Lai, Q.; Wu, M.; Wang, R.; Lai, W.; Tao, Y.; Lu, Y.; Wang, Y.; Yu, L.; Zhang, R.; Peng, Y.; et al. Cryptophycin-55/52 based antibody-drug conjugates: Synthesis, efficacy, and mode of action studies. Eur. J. Med. Chem. 2020, 199, 112364. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.M.; Collins, I. Recent findings and future directions for interpolar mitotic kinesin inhibitors in cancer therapy. Future Med. Chem. 2016, 8, 463–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerchen, H.-G.; Wittrock, S.; Stelte-Ludwig, B.; Sommer, A.; Berndt, S.; Griebenow, N.; Rebstock, A.-S.; Johannes, S.; Cancho-Grande, Y.; Mahlert, C.; et al. Antibody-Drug Conjugates with Pyrrole-Based KSP Inhibitors as the Payload Class. Angew. Chem. Int. Ed. Engl. 2018, 57, 15243–15247. [Google Scholar] [CrossRef]
- Karpov, A.S.; Nieto-Oberhuber, C.M.; Abrams, T.; Beng-Louka, E.; Blanco, E.; Chamoin, S.; Chene, P.; Dacquignies, I.; Daniel, D.; Dillon, M.P.; et al. Discovery of Potent and Selective Antibody-Drug Conjugates with Eg5 Inhibitors through Linker and Payload Optimization. ACS Med. Chem. Lett. 2019, 10, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Mantaj, J.; Jackson, P.J.M.; Rahman, K.M.; Thurston, D.E. From Anthramycin to Pyrrolobenzodiazepine (PBD)-Containing Antibody-Drug Conjugates (ADCs). Angew. Chem. Int. Ed. Engl. 2017, 56, 462–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frigerio, M.; Kyle, A.F. The Chemical Design and Synthesis of Linkers Used in Antibody Drug Conjugates. Curr. Top. Med. Chem. 2017, 17, 3393–3424. [Google Scholar] [CrossRef]
- Archer, K.E.; Reid, E.E.; Shizuka, M.; Woods, J.; Harris, L.; Maloney, E.K.; Bartle, L.M.; Ab, O.; Wilhelm, A.; Setiady, Y.; et al. Synthesis of Highly Potent N-10 Amino-Linked DNA-Alkylating Indolinobenzodiazepine Antibody-Drug Conjugates (ADCs). ACS Med. Chem. Lett. 2019, 10, 1211–1215. [Google Scholar] [CrossRef]
- Gregson, S.J.; Masterson, L.A.; Wei, B.; Pillow, T.H.; Spencer, S.D.; Kang, G.-D.; Yu, S.-F.; Raab, H.; Lau, J.; Li, G.; et al. Pyrrolobenzodiazepine Dimer Antibody-Drug Conjugates: Synthesis and Evaluation of Noncleavable Drug-Linkers. J. Med. Chem. 2017, 60, 9490–9507. [Google Scholar] [CrossRef]
- Boger, D.L.; Ishizaki, T.; Wysocki, R.J.; Munk, S.A.; Kitos, P.A.; Suntornwat, O. Total synthesis and evaluation of (.+-.)-N-(tert-butoxycarbonyl)-CBI, (+-.)-CBI-CDPI1, and (+-.)-CBI-CDPI2: CC-1065 functional agents incorporating the equivalent 1,2,9,9a-tetrahydrocyclopropa[1,2-c]benz[1,2-e]indol-4-one (CBI) left-hand subunit. J. Am. Chem. Soc. 1989, 111, 6461–6463. [Google Scholar] [CrossRef]
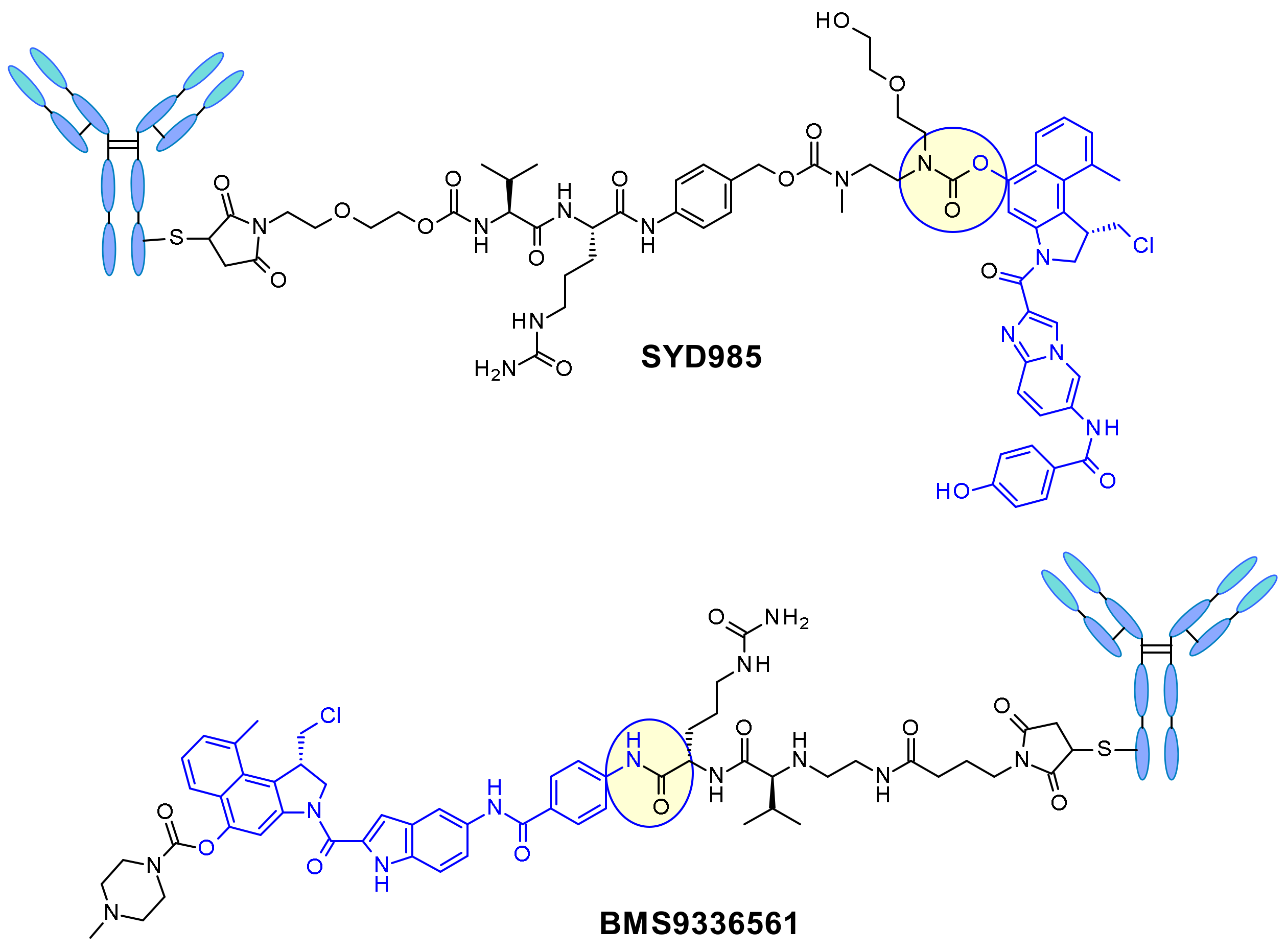
- Elgersma, R.C.; Coumans, R.G.; Huijbregts, T.; Menge, W.M.; Joosten, J.A.; Spijker, H.J.; de Groot, F.M.; van der Lee, M.M.; Ubink, R.; van den Dobbelsteen, D.J.; et al. Design, Synthesis, and Evaluation of Linker-Duocarmycin Payloads: Toward Selection of HER2-Targeting Antibody-Drug Conjugate SYD985. Mol. Pharm. 2015, 12, 1813–1835. [Google Scholar] [CrossRef]
- Owonikoko, T.K.; Hussain, A.; Stadler, W.M.; Smith, D.C.; Kluger, H.; Molina, A.M.; Gulati, P.; Shah, A.; Ahlers, C.M.; Cardarelli, P.M.; et al. First-in-human multicenter phase I study of BMS-936561 (MDX-1203), an antibody-drug conjugate targeting CD70. Cancer Chemother. Pharmacol. 2016, 77, 155–162. [Google Scholar] [CrossRef]
- Govindan, S.V.; Cardillo, T.M.; Goldenberg, D.M. Topoisomerase Inhibitors as Antibody–Drug Conjugate (ADC) Payloads. In Cytotoxic Payloads for Antibody–Drug Conjugates; Thurston, D.E., Jackson, P.J.M., Eds.; Royal Society of Chemistry: Cambridge, UK, 2019; Chapter 8; pp. 166–186. ISBN 978-1-78801-077-1. [Google Scholar]
- Zhao, H.; Lee, C.; Sai, P.; Choe, Y.H.; Boro, M.; Pendri, A.; Guan, S.; Greenwald, R.B. 20-O-Acylcamptothecin Derivatives: Evidence for Lactone Stabilization. J. Org. Chem. 2000, 65, 4601–4606. [Google Scholar] [CrossRef] [PubMed]
- Venditto, V.J.; Simanek, E.E. Cancer therapies utilizing the camptothecins: A review of the in vivo literature. Mol. Pharm. 2010, 7, 307–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogitani, Y.; Hagihara, K.; Oitate, M.; Naito, H.; Agatsuma, T. Bystander killing effect of DS-8201a, a novel anti-human epidermal growth factor receptor 2 antibody-drug conjugate, in tumors with human epidermal growth factor receptor 2 heterogeneity. Cancer Sci. 2016, 107, 1039–1046. [Google Scholar] [CrossRef]
- Nakada, T.; Sugihara, K.; Jikoh, T.; Abe, Y.; Agatsuma, T. The Latest Research and Development into the Antibody-Drug Conjugate, fam-Trastuzumab Deruxtecan (DS-8201a), for HER2 Cancer Therapy. Chem. Pharm. Bull. 2019, 67, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Veale, K.H.; Qiu, Q.; Sinkevicius, K.W.; Maloney, E.K.; Costoplus, J.A.; Lau, J.; Evans, H.L.; Setiady, Y.; Ab, O.; et al. Synthesis and Evaluation of Camptothecin Antibody-Drug Conjugates. ACS Med. Chem. Lett. 2019, 10, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Wang, H.; Taylor, T.; Warabi, K.; Huang, X.-H.; Andersen, R.J. Uncialamycin, a new enediyne antibiotic. Org. Lett. 2005, 7, 5233–5236. [Google Scholar] [CrossRef]
- Chowdari, N.S.; Gangwar, S.; Sufi, B. Enediyne Compounds, Conjugates Thereof, and Uses and Methods Therefor. U.S. Patent 8709431B2, 29 April 2014. [Google Scholar]
- Nicolaou, K.C.; Lu, M.; Mandal, D.; Gangwar, S.; Chowdari, N.S.; Poudel, Y.B. Derivatives of Uncialamycin, Methods of Synthesis and Their Use as Antitumor Agents. Patent No. WO 2015023879 A1, 19 February 2015. [Google Scholar]
- Poudel, Y.B.; Rao, C.; Kotapati, S.; Deshpande, M.; Thevanayagam, L.; Pan, C.; Cardarelli, J.; Chowdari, N.; Kaspady, M.; Samikannu, R.; et al. Design, synthesis and biological evaluation of phenol-linked uncialamycin antibody-drug conjugates. Bioorg. Med. Chem. Lett. 2020, 30, 126782. [Google Scholar] [CrossRef]
- Boghaert, E.R.; Souers, A.J.; PHILLIPS, A.C.; Judd, A.S.; Bruncko, M. Anti-EGFR Antibody Drug Conjugates. Patent No. WO 2017214282Al, 30 January 2019. [Google Scholar]
- Boghaert, E.R.; Souers, A.J.; PHILLIPS, A.C.; Judd, A.S.; Bruncko, M. Anti-EGFR Antibody Drug Conjugates. Patent No. WO 2017214301A1, 14 December 2017. [Google Scholar]
- Butler, M.S. Remediating cancer via splicing modulation. J. Med. Chem. 2013, 56, 6573–6575. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Biswas, S.; Berg, M.G.; Antapli, C.M.; Xie, F.; Wang, Q.; Tang, M.-C.; Tang, G.-L.; Zhang, L.; Dreyfuss, G.; et al. Genomics-guided discovery of thailanstatins A, B, and C As pre-mRNA splicing inhibitors and antiproliferative agents from Burkholderia thailandensis MSMB43. J. Nat. Prod. 2013, 76, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Puthenveetil, S.; Loganzo, F.; He, H.; Dirico, K.; Green, M.; Teske, J.; Musto, S.; Clark, T.; Rago, B.; Koehn, F.; et al. Natural Product Splicing Inhibitors: A New Class of Antibody-Drug Conjugate (ADC) Payloads. Bioconjug. Chem. 2016, 27, 1880–1888. [Google Scholar] [CrossRef] [PubMed]
- Dirico, K.J.; Eustaquio, A.S.; Green, M.E.; He, H.; He, M.; Koehn, F.E.; O’Donnell, C.J.; Puthenveetil, S.; Ratnayake, A.S.; Subramanyam, C. Preparation of Spliceostatin Analog Antibody Conjugates for Cancer Treatment. Patent No. WO 2014068443 A1, 8 May 2014. [Google Scholar]
- Danielczyk, A.; Stahn, R.; Faulstich, D.; Löffler, A.; Märten, A.; Karsten, U.; Goletz, S. PankoMab: A potent new generation anti-tumour MUC1 antibody. Cancer Immunol. Immunother. 2006, 55, 1337–1347. [Google Scholar] [CrossRef]
- Moldenhauer, G.; Salnikov, A.V.; Lüttgau, S.; Herr, I.; Anderl, J.; Faulstich, H. Therapeutic potential of amanitin-conjugated anti-epithelial cell adhesion molecule monoclonal antibody against pancreatic carcinoma. J. Natl. Cancer Inst. 2012, 104, 622–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurston, D.E.; Jackson, P.J.M. (Eds.) Cytotoxic Payloads for Antibody–Drug Conjugates; Royal Society of Chemistry: Cambridge, UK, 2019; pp. 11–21. ISBN 978-1-78801-077-1. [Google Scholar]
- Park, S.; Kim, S.Y.; Cho, J.; Jung, D.; Seo, D.; Lee, J.; Lee, S.; Yun, S.; Lee, H.; Park, O.; et al. Aryl Sulfate is a Useful Motif for Conjugating and Releasing Phenolic Molecules: Sulfur Fluorine Exchange Click Chemistry Enables Discovery of Ortho-Hydroxy-Protected Aryl Sulfate Linker. Bioconjug. Chem. 2019, 30, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Karpov, A.S.; Abrams, T.; Clark, S.; Raikar, A.; D’Alessio, J.A.; Dillon, M.P.; Gesner, T.G.; Jones, D.; Lacaud, M.; Mallet, W.; et al. Nicotinamide Phosphoribosyltransferase Inhibitor as a Novel Payload for Antibody-Drug Conjugates. ACS Med. Chem. Lett. 2018, 9, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.R.; Kale, A.J.; Fenley, A.T.; Byrum, T.; Debonsi, H.M.; Gilson, M.K.; Valeriote, F.A.; Moore, B.S.; Gerwick, W.H. The carmaphycins: New proteasome inhibitors exhibiting an α,β-epoxyketone warhead from a marine cyanobacterium. Chembiochem 2012, 13, 810–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almaliti, J.; Miller, B.; Pietraszkiewicz, H.; Glukhov, E.; Naman, C.B.; Kline, T.; Hanson, J.; Li, X.; Zhou, S.; Valeriote, F.A.; et al. Exploration of the carmaphycins as payloads in antibody drug conjugate anticancer agents. Eur. J. Med. Chem. 2019, 161, 416–432. [Google Scholar] [CrossRef]
- Erickson, H.K.; Widdison, W.C.; Mayo, M.F.; Whiteman, K.; Audette, C.; Wilhelm, S.D.; Singh, R. Tumor delivery and in vivo processing of disulfide-linked and thioether-linked antibody-maytansinoid conjugates. Bioconjug. Chem. 2010, 21, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Kovtun, Y.V.; Audette, C.A.; Ye, Y.; Xie, H.; Ruberti, M.F.; Phinney, S.J.; Leece, B.A.; Chittenden, T.; Blättler, W.A.; Goldmacher, V.S. Antibody-drug conjugates designed to eradicate tumors with homogeneous and heterogeneous expression of the target antigen. Cancer Res. 2006, 66, 3214–3221. [Google Scholar] [CrossRef] [Green Version]
- Walles, M.; Connor, A.; Hainzl, D. ADME and Safety Aspects of Non-cleavable Linkers in Drug Discovery and Development. Curr. Top. Med. Chem. 2017, 17, 3463–3475. [Google Scholar] [CrossRef] [PubMed]
- Saito, G.; Swanson, J.A.; Lee, K.-D. Drug delivery strategy utilizing conjugation via reversible disulfide linkages: Role and site of cellular reducing activities. Adv. Drug Deliv. Rev. 2003, 55, 199–215. [Google Scholar] [CrossRef]
- Kellogg, B.A.; Garrett, L.; Kovtun, Y.; Lai, K.C.; Leece, B.; Miller, M.; Payne, G.; Steeves, R.; Whiteman, K.R.; Widdison, W.; et al. Disulfide-linked antibody-maytansinoid conjugates: Optimization of in vivo activity by varying the steric hindrance at carbon atoms adjacent to the disulfide linkage. Bioconjug. Chem. 2011, 22, 717–727. [Google Scholar] [CrossRef]
- Hong, E.E.; Erickson, H.; Lutz, R.J.; Whiteman, K.R.; Jones, G.; Kovtun, Y.; Blanc, V.; Lambert, J.M. Design of Coltuximab Ravtansine, a CD19-Targeting Antibody-Drug Conjugate (ADC) for the Treatment of B-Cell Malignancies: Structure-Activity Relationships and Preclinical Evaluation. Mol. Pharm. 2015, 12, 1703–1716. [Google Scholar] [CrossRef] [PubMed]
- Sapra, P.; Stein, R.; Pickett, J.; Qu, Z.; Govindan, S.V.; Cardillo, T.M.; Hansen, H.J.; Horak, I.D.; Griffiths, G.L.; Goldenberg, D.M. Anti-CD74 antibody-doxorubicin conjugate, IMMU-110, in a human multiple myeloma xenograft and in monkeys. Clin. Cancer Res. 2005, 11, 5257–5264. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, H.; Viskov, C.; Garcia-Echeverria, C. Antibody-drug conjugates—A new wave of cancer drugs. Bioorg. Med. Chem. Lett. 2014, 24, 5357–5363. [Google Scholar] [CrossRef] [Green Version]
- Hamann, P.R.; Hinman, L.M.; Hollander, I.; Beyer, C.F.; Lindh, D.; Holcomb, R.; Hallett, W.; Tsou, H.-R.; Upeslacis, J.; Shochat, D.; et al. Gemtuzumab ozogamicin, a potent and selective anti-CD33 antibody-calicheamicin conjugate for treatment of acute myeloid leukemia. Bioconjug. Chem. 2002, 13, 47–58. [Google Scholar] [CrossRef]
- Kantarjian, H.M.; DeAngelo, D.J.; Stelljes, M.; Martinelli, G.; Liedtke, M.; Stock, W.; Gökbuget, N.; O’Brien, S.; Wang, K.; Wang, T.; et al. Inotuzumab Ozogamicin versus Standard Therapy for Acute Lymphoblastic Leukemia. N. Engl. J. Med. 2016, 375, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Doronina, S.O.; Toki, B.E.; Torgov, M.Y.; Mendelsohn, B.A.; Cerveny, C.G.; Chace, D.F.; DeBlanc, R.L.; Gearing, R.P.; Bovee, T.D.; Siegall, C.B.; et al. Development of potent monoclonal antibody auristatin conjugates for cancer therapy. Nat. Biotechnol. 2003, 21, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Schröder, B.A.; Wrocklage, C.; Hasilik, A.; Saftig, P. The proteome of lysosomes. Proteomics 2010, 10, 4053–4076. [Google Scholar] [CrossRef] [PubMed]
- Dubowchik, G.M.; Mosure, K.; Knipe, J.O.; Firestone, R.A. Cathepsin B-sensitive dipeptide prodrugs. 2. Models of anticancer drugs paclitaxel (Taxol®), mitomycin C and doxorubicin. Bioorg. Med. Chem. Lett. 1998, 8, 3347–3352. [Google Scholar] [CrossRef]
- Dubowchik, G.M.; Firestone, R.A.; Padilla, L.; Willner, D.; Hofstead, S.J.; Mosure, K.; Knipe, J.O.; Lasch, S.J.; Trail, P.A. Cathepsin B-labile dipeptide linkers for lysosomal release of doxorubicin from internalizing immunoconjugates: Model studies of enzymatic drug release and antigen-specific in vitro anticancer activity. Bioconjug. Chem. 2002, 13, 855–869. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, S.C.; Nguyen, M.T.; Andreyka, J.B.; Meyer, D.L.; Doronina, S.O.; Senter, P.D. Dipeptide-based highly potent doxorubicin antibody conjugates. Bioorg. Med. Chem. Lett. 2006, 16, 358–362. [Google Scholar] [CrossRef]
- Dal Corso, A.; Cazzamalli, S.; Gébleux, R.; Mattarella, M.; Neri, D. Protease-Cleavable Linkers Modulate the Anticancer Activity of Noninternalizing Antibody-Drug Conjugates. Bioconjug. Chem. 2017, 28, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fan, S.; Zhong, W.; Zhou, X.; Li, S. Development and Properties of Valine-Alanine based Antibody-Drug Conjugates with Monomethyl Auristatin E as the Potent Payload. Int. J. Mol. Sci. 2017, 18, 1860. [Google Scholar] [CrossRef]
- Bryden, F.; Martin, C.; Letast, S.; Lles, E.; Viéitez-Villemin, I.; Rousseau, A.; Colas, C.; Brachet-Botineau, M.; Allard-Vannier, E.; Larbouret, C.; et al. Impact of cathepsin B-sensitive triggers and hydrophilic linkers on in vitro efficacy of novel site-specific antibody-drug conjugates. Org. Biomol. Chem. 2018, 16, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Ogitani, Y.; Aida, T.; Hagihara, K.; Yamaguchi, J.; Ishii, C.; Harada, N.; Soma, M.; Okamoto, H.; Oitate, M.; Arakawa, S.; et al. DS-8201a, A Novel HER2-Targeting ADC with a Novel DNA Topoisomerase I Inhibitor, Demonstrates a Promising Antitumor Efficacy with Differentiation from T-DM1. Clin. Cancer Res. 2016, 22, 5097–5108. [Google Scholar] [CrossRef] [Green Version]
- Andrikopoulou, A.; Zografos, E.; Liontos, M.; Koutsoukos, K.; Dimopoulos, M.-A.; Zagouri, F. Trastuzumab Deruxtecan (DS-8201a): The Latest Research and Advances in Breast Cancer. Clin. Breast Cancer 2020. [Google Scholar] [CrossRef]
- Kern, J.C.; Dooney, D.; Zhang, R.; Liang, L.; Brandish, P.E.; Cheng, M.; Feng, G.; Beck, A.; Bresson, D.; Firdos, J.; et al. Novel Phosphate Modified Cathepsin B Linkers: Improving Aqueous Solubility and Enhancing Payload Scope of ADCs. Bioconjug. Chem. 2016, 27, 2081–2088. [Google Scholar] [CrossRef]
- Kern, J.C.; Cancilla, M.; Dooney, D.; Kwasnjuk, K.; Zhang, R.; Beaumont, M.; Figueroa, I.; Hsieh, S.; Liang, L.; Tomazela, D.; et al. Discovery of Pyrophosphate Diesters as Tunable, Soluble, and Bioorthogonal Linkers for Site-Specific Antibody-Drug Conjugates. J. Am. Chem. Soc. 2016, 138, 1430–1445. [Google Scholar] [CrossRef]
- Jeffrey, S.C.; Andreyka, J.B.; Bernhardt, S.X.; Kissler, K.M.; Kline, T.; Lenox, J.S.; Moser, R.F.; Nguyen, M.T.; Okeley, N.M.; Stone, I.J.; et al. Development and properties of beta-glucuronide linkers for monoclonal antibody-drug conjugates. Bioconjug. Chem. 2006, 17, 831–840. [Google Scholar] [CrossRef]
- Jeffrey, S.C.; Nguyen, M.T.; Moser, R.F.; Meyer, D.L.; Miyamoto, J.B.; Senter, P.D. Minor groove binder antibody conjugates employing a water soluble beta-glucuronide linker. Bioorg. Med. Chem. Lett. 2007, 17, 2278–2280. [Google Scholar] [CrossRef]
- Burke, P.J.; Senter, P.D.; Meyer, D.W.; Miyamoto, J.B.; Anderson, M.; Toki, B.E.; Manikumar, G.; Wani, M.C.; Kroll, D.J.; Jeffrey, S.C. Design, synthesis, and biological evaluation of antibody-drug conjugates comprised of potent camptothecin analogues. Bioconjug. Chem. 2009, 20, 1242–1250. [Google Scholar] [CrossRef]
- Jeffrey, S.C.; Brabander, J.; de Miyamoto, J.; Senter, P.D. Expanded Utility of the β-Glucuronide Linker: ADCs That Deliver Phenolic Cytotoxic Agents. ACS Med. Chem. Lett. 2010, 1, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Lyon, R.P.; Bovee, T.D.; Doronina, S.O.; Burke, P.J.; Hunter, J.H.; Neff-LaFord, H.D.; Jonas, M.; Anderson, M.E.; Setter, J.R.; Senter, P.D. Reducing hydrophobicity of homogeneous antibody-drug conjugates improves pharmacokinetics and therapeutic index. Nat. Biotechnol. 2015, 33, 733–735. [Google Scholar] [CrossRef] [PubMed]
- Kolodych, S.; Michel, C.; Delacroix, S.; Koniev, O.; Ehkirch, A.; Eberova, J.; Cianférani, S.; Renoux, B.; Krezel, W.; Poinot, P.; et al. Development and evaluation of β-galactosidase-sensitive antibody-drug conjugates. Eur. J. Med. Chem. 2017, 142, 376–382. [Google Scholar] [CrossRef]
- Papot, S.; Tranoy, I.; Tillequin, F.; Florent, J.-C.; Gesson, J.-P. Design of selectively activated anticancer prodrugs: Elimination and cyclization strategies. Curr. Med. Chem. Anticancer Agents 2002, 2, 155–185. [Google Scholar] [CrossRef] [PubMed]
- Legigan, T.; Clarhaut, J.; Tranoy-Opalinski, I.; Monvoisin, A.; Renoux, B.; Thomas, M.; Le Pape, A.; Lerondel, S.; Papot, S. The first generation of β-galactosidase-responsive prodrugs designed for the selective treatment of solid tumors in prodrug monotherapy. Angew. Chem. Int. Ed. Engl. 2012, 51, 11606–11610. [Google Scholar] [CrossRef] [PubMed]
- Bargh, J.D.; Walsh, S.J.; Isidro-Llobet, A.; Omarjee, S.; Carroll, J.S.; Spring, D.R. Sulfatase-cleavable linkers for antibody-drug conjugates. Chem. Sci. 2020, 11, 2375–2380. [Google Scholar] [CrossRef] [Green Version]
- Albin, N.; Massaad, L.; Toussaint, C.; Mathieu, M.C.; Morizet, J.; Parise, O.; Gouyette, A.; Chabot, G.G. Main drug-metabolizing enzyme systems in human breast tumors and peritumoral tissues. Cancer Res. 1993, 53, 3541–3546. [Google Scholar] [PubMed]
- Rabuka, D.; Lui, J.; Chuprakov, S. Sulfatase-Cleavable Linkers for Antibody-Drug-Conjugates. Patent No. WO2020/096775, 14 May 2020. [Google Scholar]
- Yamada, K.; Ito, Y. Recent Chemical Approaches for Site-Specific Conjugation of Native Antibodies: Technologies toward Next-Generation Antibody–Drug Conjugates. ChemBioChem 2019, 20, 2729–2737. [Google Scholar] [CrossRef] [PubMed]
- Hamann, P.R. The genesis of the antibody conjugate gemtuzumab ozogamicin (Mylotarg ® for acute myeloid leukemia. In Accounts in Drug Discovery: Case Studies in Medicinal Chemistry; Barrish, J.C., Carter, P.H., Cheng, P.T.W., Zahler, R., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2011; Chapter 5; pp. 103–119. [Google Scholar]
- Chen, Y.; Kim, M.T.; Zheng, L.; Deperalta, G.; Jacobson, F. Structural characterization of cross-linked species in Trastuzumab emtansine (Kadcyla). BioconjugChem 2016, 27, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.J.; Oliveira, B.L.; Martínez-Sáez, N.; Gunerreiro, A.; Cal, P.M.S.D.; Bertoldo, J.; Maneiro, M.; Perkins, E.; Howard, J.; Deery, M.K.; et al. Chemo- and Regioselective Lysine Modification on Native Proteins. J. Am. Chem. Soc. 2018, 140, 4004–4017. [Google Scholar] [CrossRef] [PubMed]
- Adusumalli, S.R.; Rawale, D.G.; Singh, U.; Tripathi, P.; Paul, R.; Kalra, N.; Mishra, R.K.; Shukla, S.; Rai, V. Single-Site Labeling of Native Proteins Enabled by a Chemoselective and Site-Selective Chemical Technology. J. Am. Chem. Soc. 2018, 140, 15114–15123. [Google Scholar] [CrossRef] [PubMed]
- Adusumalli, S.R.; Rawale, D.G.; Thakur, K.; Purushottam, L.; Reddy, N.C.; Kalra, N.; Shukla, S.; Rai, V. Chemoselective and Site-Selective Lysine-Directed Lysine Modification Enables Single-Site Labeling of Native Proteins. Angew. Chem. Int. Ed. Engl. 2020, 59, 10332–10336. [Google Scholar] [CrossRef] [PubMed]
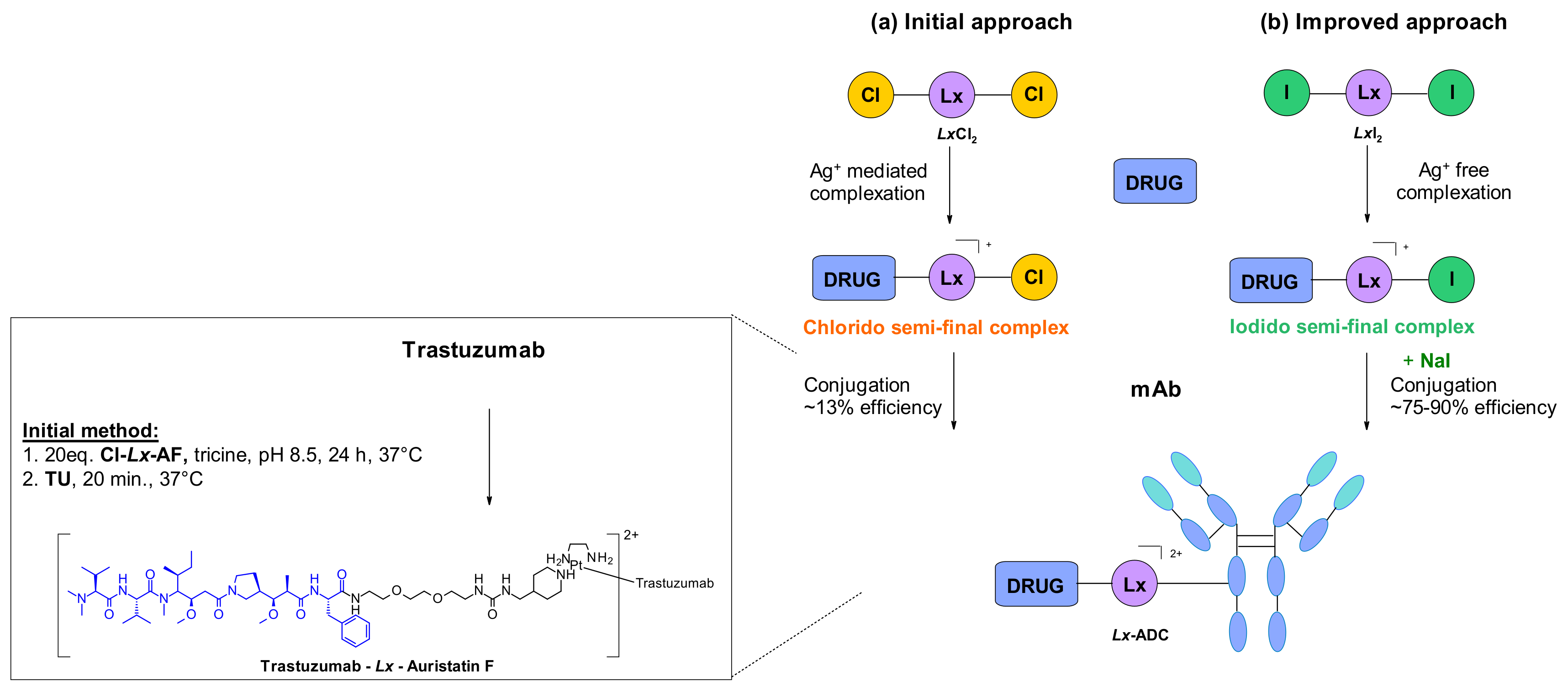
- Merkul, E.; Muns, J.A.; Sijbrandi, N.J.; Houthoff, H.-J.; Nijmeijer, B.; Rheenen, G.V.; Reedijk, J.; Dongen, G.A.M.S.V. An Efficient Conjugation Approach for Coupling Drugs to Native Antibodies via the PtII Linker Lx for Improved Manufacturability of Antibody–Drug Conjugates. Angew. Chem. Int. Ed. 2021, 60, 2–10. [Google Scholar]
- Merkul, E.; Sijbrandi, N.J.; Muns, J.A.; Aydin, I.; Adamzek, K.; Houthoff, H.-J.; Nijmeijer, B.; Dongen, G.A.M.S.V. First platinum(II)-based metal-organic linker technology (Lx®) for a plug-and-play development of antibody-drug conjugates (ADCs). Expert Opin. Drug Deliv. 2019, 16, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Merkul, E.; Sijbrandi, N.J.; Aydin, I.; Muns, J.A.; Peters, R.J.R.W.; Laarhoven, P.; Houthoff, H.-J.; van Dongen, G.A.M.S. A successful search for new, efficient, and silver-free manufacturing processes for key platinum(II) intermediates applied in antibody–drug conjugate (ADC) production. Green Chem. 2020, 22, 2203–2212. [Google Scholar] [CrossRef]
- Liu, H.; May, K. Disulfide bond structures of IgG molecules: Structural variations, chemical modifications and possible impacts to stability and biological function. MAbs 2012, 4, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamblett, K.J.; Senter, P.D.; Chace, D.F.; Sun, M.M.C.; Lenox, J.; Cerveny, C.G.; Kissler, K.M.; Bernhardt, S.X.; Kopcha, A.K.; Zabinski, R.F.; et al. Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate. Clin. Cancer Res. 2004, 10, 7063–7070. [Google Scholar] [CrossRef] [Green Version]
- Beckley, N.S.; Lazzareschi, K.P.; Chih, H.-W.; Sharma, V.K.; Flores, H.L. Investigation into Temperature-Induced Aggregation of an Antibody Drug Conjugate. Bioconjug. Chem. 2013, 24, 1674–1683. [Google Scholar] [CrossRef]
- Badescu, G.; Bryant, P.; Bird, M.; Henseleit, K.; Swierkosz, J.; Parekh, V.; Tommasi, R.; Pawlisz, E.; Jurlewicz, K.; Farys, M.; et al. Bridging disulfides for stable and defined antibody drug conjugates. Bioconjug. Chem. 2014, 25, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Riegger, A.; Lamla, M.; Wiese, S.; Oeckl, P.; Otto, K.-M.; Wu, Y.; Fischer, S.; Barth, H.; Kuan, S.L.; et al. Water-soluble allyl sulfones for dual site-specific labelling of proteins and cyclic peptides. Chem. Sci. 2016, 7, 3234–3239. [Google Scholar] [CrossRef] [Green Version]
- Griebenow, N.; Dilmaç, A.M.; Greven, S.; Bräse, S. Site-Specific Conjugation of Peptides and Proteins via Rebridging of Thiol Disulfide Bonds Using the Thiol-Yne Coupling Reaction. Bioconjug. Chem. 2016, 27, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, F.F.; Nunes, J.P.M.; Maruani, A.; Chudasama, V.; Smith, M.E.B.; Chester, K.A.; Baker, J.R.; Caddick, S. Next generation maleimides enable the controlled assembly of antibody-drug conjugates via native disulfide bond bridging. Org. Biomol. Chem. 2014, 12, 7261–7269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuillâtre, O.; Gély, C.; Huvelle, S.; Baltus, C.B.; Juen, L.; Joubert, N.; Desgranges, A.; Viaud-Massuard, M.-C.; Martin, C. Impact of Maleimide Disubstitution on Chemical and Biological Characteristics of HER2 Antibody–Drug Conjugates. ACS Omega 2020, 5, 1557–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruani, A.; Smith, M.E.B.; Miranda, E.; Chester, K.A.; Chudasama, V.; Caddick, S. A plug-and-play approach to antibody-based therapeutics via a chemoselective dual click strategy. Nat. Commun. 2015, 6, 6645. [Google Scholar] [CrossRef] [PubMed]
- Bahou, C.; Richards, D.A.; Maruani, A.; Love, E.A.; Javaid, F.; Caddick, S.; Baker, J.R.; Chudasama, V. Highly homogeneous antibody modification through optimisation of the synthesis and conjugation of functionalised dibromopyridazinediones. Org. Biomol. Chem. 2018, 16, 1359–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.J.; Omarjee, S.; Galloway, W.R.J.D.; Kwan, T.T.-L.; Sore, H.F.; Parker, J.S.; Hyvönen, M.; Carroll, J.S.; Spring, D.R. A general approach for the site-selective modification of native proteins, enabling the generation of stable and functional antibody–drug conjugates. Chem. Sci. 2019, 10, 694–700. [Google Scholar] [CrossRef] [Green Version]
- Walsh, S.J.; Iegre, J.; Seki, H.; Bargh, J.D.; Sore, H.F.; Parker, J.S.; Carroll, J.S.; Spring, D.R. General dual functionalisation of biomacromolecules via a cysteine bridging strategy. Org. Biomol. Chem. 2020, 18, 4224–4230. [Google Scholar] [CrossRef] [PubMed]
- Counsell, A.J.; Walsh, S.J.; Robertson, N.S.; Sore, H.F.; Spring, D.R. Efficient and selective antibody modification with functionalised divinyltriazines. Org. Biomol. Chem. 2020, 18, 4739–4743. [Google Scholar] [CrossRef] [PubMed]
- Toda, N.; Asano, S.; Barbas, C.F., 3rd. Rapid, stable, chemoselective labeling of thiols with Julia-Kocieński-like reagents: A serum-stable alternative to maleimide-based protein conjugation. Angew. Chem. Int. Ed. Engl. 2013, 52, 12592–12596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, J.T.; Asano, S.; Li, X.; Rader, C.; Barbas, C.F., 3rd. Improving the serum stability of site-specific antibody conjugates with sulfone linkers. Bioconjug. Chem. 2014, 25, 1402–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adumeau, P.; Davydova, M.; Zeglis, B.M. Thiol-Reactive Bifunctional Chelators for the Creation of Site-Selectively Modified Radioimmunoconjugates with Improved Stability. Bioconjug. Chem. 2018, 29, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Sarbisheh, E.K.; Dewaele-Le Roi, G.; Shannon, W.E.; Tan, S.; Xu, Y.; Zeglis, B.M.; Price, E.W. DiPODS: A Reagent for Site-Specific Bioconjugation via the Irreversible Rebridging of Disulfide Linkages. Bioconjug. Chem. 2020, 31, 2789–2806. [Google Scholar]
- Davydova, M.; Dewaele Le Roi, G.; Adumeau, P.; Zeglis, B.M. Synthesis and Bioconjugation of Thiol-Reactive Reagents for the Creation of Site-Selectively Modified Immunoconjugates. J. Vis. Exp. 2019, 145, 59063. [Google Scholar] [CrossRef] [PubMed]
- Bahou, C.; Love, E.A.; Leonard, S.; Spears, R.J.; Maruani, A.; Armour, K.; Baker, J.R.; Chudasama, V. Disulfide Modified IgG1: An Investigation of Biophysical Profile and Clinically Relevant Fc Interactions. Bioconjug. Chem. 2019, 30, 1048–1054. [Google Scholar] [CrossRef]
- Zuberbühler, K.; Casi, G.; Bernardes, G.J.L.; Neri, D. Fucose-specific conjugation of hydrazide derivatives to a vascular-targeting monoclonal antibody in IgG format. Chem. Commun. 2012, 48, 7100–7102. [Google Scholar] [CrossRef]
- Okeley, N.M.; Toki, B.E.; Zhang, X.; Jeffrey, S.C.; Burke, P.J.; Alley, S.C.; Senter, P.D. Metabolic Engineering of Monoclonal Antibody Carbohydrates for Antibody–Drug Conjugation. Bioconjug. Chem. 2013, 24, 1650–1655. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Stefano, J.E.; Manning, C.; Kyazike, J.; Chen, B.; Gianolio, D.A.; Park, A.; Busch, M.; Bird, J.; Zheng, X.; et al. Site-Specific Antibody–Drug Conjugation through Glycoengineering. Bioconjug. Chem. 2014, 25, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, T.; Boons, G.-J. Preparation of well-defined antibody-drug conjugates through glycan remodeling and strain-promoted azide-alkyne cycloadditions. Angew. Chem. Int. Ed. 2014, 53, 7179–7182. [Google Scholar] [CrossRef] [Green Version]
- Boeggeman, E.; Ramakrishnan, B.; Pasek, M.; Manzoni, M.; Puri, A.; Loomis, K.H.; Waybright, T.J.; Qasba, P.K. Site specific conjugation of fluoroprobes to the remodeled Fc N-glycans of monoclonal antibodies using mutant glycosyltransferases: Application for cell surface antigen detection. Bioconjug. Chem. 2009, 20, 1228–1236. [Google Scholar] [CrossRef] [Green Version]
- Zeglis, B.M.; Davis, C.B.; Aggeler, R.; Kang, H.C.; Chen, A.; Agnew, B.J.; Lewis, J.S. Enzyme-mediated methodology for the site-specific radiolabeling of antibodies based on catalyst-free click chemistry. Bioconjug. Chem. 2013, 24, 1057–1067. [Google Scholar] [CrossRef]
- Zhu, Z.; Ramakrishnan, B.; Li, J.; Wang, Y.; Feng, Y.; Ponraj Prabakaran, P.; Colantonio, S.; Dyba, M.A.; Qasba, P.K.; Dimitrov, D.S. Site-specific antibody-drug conjugation through an engineered glycotransferase and a chemically reactive sugar. MAbs 2014, 6, 1190–1200. [Google Scholar] [CrossRef] [Green Version]
- Sjögren, J.; Cosgrave, E.F.J.; Allhorn, M.; Nordgren, M.; Björk, S.; Olsson, F.; Fredriksson, S.; Collin, M. EndoS and EndoS2 hydrolyze Fc-glycans on therapeutic antibodies with different glycoform selectivity and can be used for rapid quantification of high-mannose glycans. Glycobiology 2015, 25, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- van Geel, R.; Wijdeven, M.A.; Heesbeen, R.; Verkade, J.M.M.; Wasiel, A.A.; van Berkel, S.S.; van Delft, F.L. Chemoenzymatic Conjugation of Toxic Payloads to the Globally Conserved N-Glycan of Native mAbs Provides Homogeneous and Highly Efficacious Antibody-Drug Conjugates. Bioconjug. Chem. 2015, 26, 2233–2242. [Google Scholar] [CrossRef]
- Albers, A.E.; Garofalo, A.W.; Drake, P.M.; Kudirka, R.W.; de Hart, G.; Barfield, R.M.; Baker, J.; Banas, S.; Rabuka, D. Exploring the effects of linker composition on site-specifically modified antibody-drug conjugates. Eur. J. Med. Chem. 2014, 88, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Falck, G.; Müller, K.M. Enzyme-Based Labeling Strategies for Antibody-Drug Conjugates and Antibody Mimetics. Antibodies 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Dennler, P.; Chiotellis, A.; Fischer, E.; Brégeon, D.; Belmant, C.; Gauthier, L.; Lhospice, F.; Romagne, F.; Schibli, R. Transglutaminase-based chemo-enzymatic conjugation approach yields homogeneous antibody-drug conjugates. Bioconjug. Chem. 2014, 25, 569–578. [Google Scholar] [CrossRef]
- Anami, Y.; Xiong, W.; Gui, X.; Deng, M.; Zhang, C.C.; Zhang, N.; An, Z.; Tsuchikama, K. Enzymatic conjugation using branched linkers for constructing homogeneous antibody-drug conjugates with high potency. Org. Biomol. Chem. 2017, 15, 5635–5642. [Google Scholar] [CrossRef]
- Lhospice, F.; Brégeon, D.; Belmant, C.; Dennler, P.; Chiotellis, A.; Fischer, E.; Gauthier, L.; Boëdec, A.; Rispaud, H.; Savard-Chambard, S.; et al. Site-Specific Conjugation of Monomethyl Auristatin E to Anti-CD30 Antibodies Improves Their Pharmacokinetics and Therapeutic Index in Rodent Models. Mol. Pharm. 2015, 12, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Dickgiesser, S.; Rieker, M.; Mueller-Pompalla, D.; Schröter, C.; Tonillo, J.; Warszawski, S.; Raab-Westphal, S.; Kühn, S.; Knehans, T.; Könning, D.; et al. Site-Specific Conjugation of Native Antibodies Using Engineered Microbial Transglutaminases. Bioconjug. Chem. 2020, 31, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Spidel, J.L.; Vaessen, B.; Albone, E.F.; Cheng, X.; Verdi, A.; Kline, J.B. Site-Specific Conjugation to Native and Engineered Lysines in Human Immunoglobulins by Microbial Transglutaminase. Bioconjug. Chem. 2017, 28, 2471–2484. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.J.; Bargh, J.D.; Dannheim, F.M.; Hanby, A.R.; Seki, H.; Counsell, A.J.; Ou, X.; Fowler, E.; Ashman, N.; Takada, Y.; et al. Site-selective modification strategies in antibody-drug conjugates. Chem. Soc. Rev. 2021, 50, 1305–1353. [Google Scholar] [CrossRef]
- Beerli, R.R.; Hell, T.; Merkel, A.S.; Grawunder, U. Sortase Enzyme-Mediated Generation of Site-Specifically Conjugated Antibody Drug Conjugates with High In Vitro and In Vivo Potency. PLoS ONE 2015, 10, e0131177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefan, N.; Gébleux, R.; Waldmeier, L.; Hell, T.; Escher, M.; Wolter, F.I.; Grawunder, U.; Beerli, R.R. Highly Potent, Anthracycline-based Antibody-Drug Conjugates Generated by Enzymatic, Site-specific Conjugation. Mol. Cancer Ther. 2017, 16, 879–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruins, J.J.; Westphal, A.H.; Albada, B.; Wagner, K.; Bartels, L.; Spits, H.; van Berkel, W.J.H.; van Delft, F.L. Inducible, Site-Specific Protein Labeling by Tyrosine Oxidation-Strain-Promoted (4 + 2) Cycloaddition. Bioconjug. Chem. 2017, 28, 1189–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tumey, L.N.; Li, F.; Rago, B.; Han, X.; Loganzo, F.; Musto, S.; Graziani, E.I.; Puthenveetil, S.; Casavant, J.; Marquette, K.; et al. Site Selection: A Case Study in the Identification of Optimal Cysteine Engineered Antibody Drug Conjugates. AAPS J. 2017, 19, 1123–1135. [Google Scholar] [CrossRef]
- Junutula, J.R.; Bhakta, S.; Raab, H.; Ervin, K.E.; Eigenbrot, C.; Vandlen, R.; Scheller, R.H.; Lowman, H.B. Rapid identification of reactive cysteine residues for site-specific labeling of antibody-Fabs. J. Immunol. Methods 2008, 332, 41–52. [Google Scholar] [CrossRef]
- Ohri, R.; Bhakta, S.; Fourie-O’Donohue, A.; Dela Cruz-Chuh, J.; Tsai, S.P.; Cook, R.; Wei, B.; Ng, C.; Wong, A.W.; Bos, A.B.; et al. High-Throughput Cysteine Scanning to Identify Stable Antibody Conjugation Sites for Maleimide- and Disulfide-Based Linkers. Bioconjug. Chem. 2018, 29, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Junutula, J.R.; Raab, H.; Clark, S.; Bhakta, S.; Leipold, D.D.; Weir, S.; Chen, Y.; Simpson, M.; Tsai, S.P.; Dennis, M.S.; et al. Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index. Nat. Biotechnol. 2008, 26, 925–932. [Google Scholar] [CrossRef]
- Shen, B.Q.; Xu, K.; Liu, L.; Raab, H.; Bhakta, S.; Kenrick, M.; Parsons-Reponte, K.L.; Tien, J.; Yu, S.F.; Mai, E.; et al. Conjugation site modulates the in vivo stability and therapeutic activity of antibody-drug conjugates. Nat. Biotechnol. 2012, 30, 184–189. [Google Scholar] [CrossRef]
- Lyon, R.P.; Setter, J.R.; Bovee, T.D.; Doronina, S.O.; Hunter, J.H.; Anderson, M.E.; Balasubramanian, C.L.; Duniho, S.M.; Leiske, C.I.; Li, F.; et al. Self-hydrolyzing maleimides improve the stability and pharmacological properties of antibody-drug conjugates. Nat. Biotechnol. 2014, 32, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Kemp, G.C.; Tiberghien, A.C.; Patel, N.V.; D’Hooge, F.; Nilapwar, S.M.; Adams, L.R.; Corbett, S.; Williams, D.G.; Hartley, J.A.; Howard, P.W. Synthesis and in vitro evaluation of SG3227, a pyrrolobenzodiazepine dimer antibody-drug conjugate payload based on sibiromycin. Bioorganic Med. Chem. Lett. 2017, 27, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Nunes, J.P.M.; Vassileva, V.; Robinson, E.; Morais, M.; Smith, M.E.B.; Pedley, R.B.; Caddick, S.; Baker, J.R.; Chudasama, V. Use of a next generation maleimide in combination with THIOMAB™ antibody technology delivers a highly stable, potent and near homogeneous THIOMAB™ antibody-drug conjugate (TDC). RSC Adv. 2017, 7, 24828–24832. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, B.L.; Stenton, B.J.; Unnikrishnan, V.B.; De Almeida, C.R.; Conde, J.; Negrão, M.; Schneider, F.S.S.; Cordeiro, C.; Ferreira, M.G.; Caramori, G.F.; et al. Platinum-Triggered Bond-Cleavage of Pentynoyl Amide and N-Propargyl Handles for Drug-Activation. J. Am. Chem. Soc. 2020, 142, 10869–10880. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.J.; Navo, C.D.; Hakala, T.; Ferhati, X.; Guerreiro, A.; Hartmann, D.; Bernardim, B.; Saar, K.L.; Companñón, I.; Corzana, F.; et al. Quaternization of Vinyl/Alkynyl Pyridine Enables Ultrafast Cysteine-Selective Protein Modification and Charge Modulation. Angew. Chem. Int. Ed. 2019, 58, 6640–6644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kline, T.; Steiner, A.R.; Penta, K.; Sato, A.K.; Hallam, T.J.; Yin, G. Methods to Make Homogenous Antibody Drug Conjugates. Pharm. Res. 2015, 32, 3480–3493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kularatne, S.A.; Deshmukh, V.; Ma, J.; Tardif, V.; Lim, R.K.V.; Pugh, H.M.; Sun, Y.; Manibusan, A.; Sellers, A.J.; Barnett, R.S.; et al. A CXCR4-targeted site-specific antibody-drug conjugate. Angew. Chem. Int. Ed. 2014, 53, 11863–11867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandish, P.E.; Palmieri, A.; Antonenko, S.; Beaumont, M.; Benso, L.; Cancilla, M.; Cheng, M.; Fayadat-Dilman, L.; Feng, G.; Figueroa, I.; et al. Development of Anti-CD74 Antibody-Drug Conjugates to Target Glucocorticoids to Immune Cells. Bioconjug. Chem. 2018, 29, 2357–2369. [Google Scholar] [CrossRef] [PubMed]
- VanBrunt, M.P.; Shanebeck, K.; Caldwell, Z.; Johnson, J.; Thompson, P.; Martin, T.; Dong, H.; Li, G.; Xu, H.; D’Hooge, F.; et al. Genetically Encoded Azide Containing Amino Acid in Mammalian Cells Enables Site-Specific Antibody-Drug Conjugates Using Click Cycloaddition Chemistry. Bioconjug. Chem. 2015, 26, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Roy, G.; Reier, J.; Garcia, A.; Martin, T.; Rice, M.; Wang, J.; Prophet, M.; Christie, R.; Dall’Acqua, W.; Ahuja, S.; et al. Development of a high yielding expression platform for the introduction of non-natural amino acids in protein sequences. MAbs 2020, 12, 1684749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oller-Salvia, B.; Kym, G.; Chin, J.W. Rapid and Efficient Generation of Stable Antibody-Drug Conjugates via an Encoded Cyclopropene and an Inverse-Electron-Demand Diels-Alder Reaction. Angew. Chem. Int. Ed. 2018, 57, 2831–2834. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Nelson, C.G.; Nair, R.R.; Hazlehurst, L.; Moroni, T.; Martinez-Acedo, P.; Nanna, A.R.; Hymel, D.; Burke, T.R.; Rader, C. Stable and Potent Selenomab-Drug Conjugates. Cell Chem. Biol. 2017, 24, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, R.; Lemos, A.; Preto, A.J.; Almeida, J.G.; Correia, J.D.G.; Sensoy, O.; Moreira, I.S. Computational Approaches in Antibody-drug Conjugate Optimization for Targeted Cancer Therapy. Curr. Top. Med. Chem. 2018, 18, 1091–1109. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.J.M.; Thurston, D.E. Use of Molecular Modelling Techniques in Antibody–Drug Conjugate (ADC) Payload Discovery and Development. In Cytotoxic Payloads for Antibody–Drug Conjugates; Jackson, P.J.M., Thurston, D.E., Eds.; Royal Society of Chemistry: London, UK, 2019; pp. 57–72. [Google Scholar]













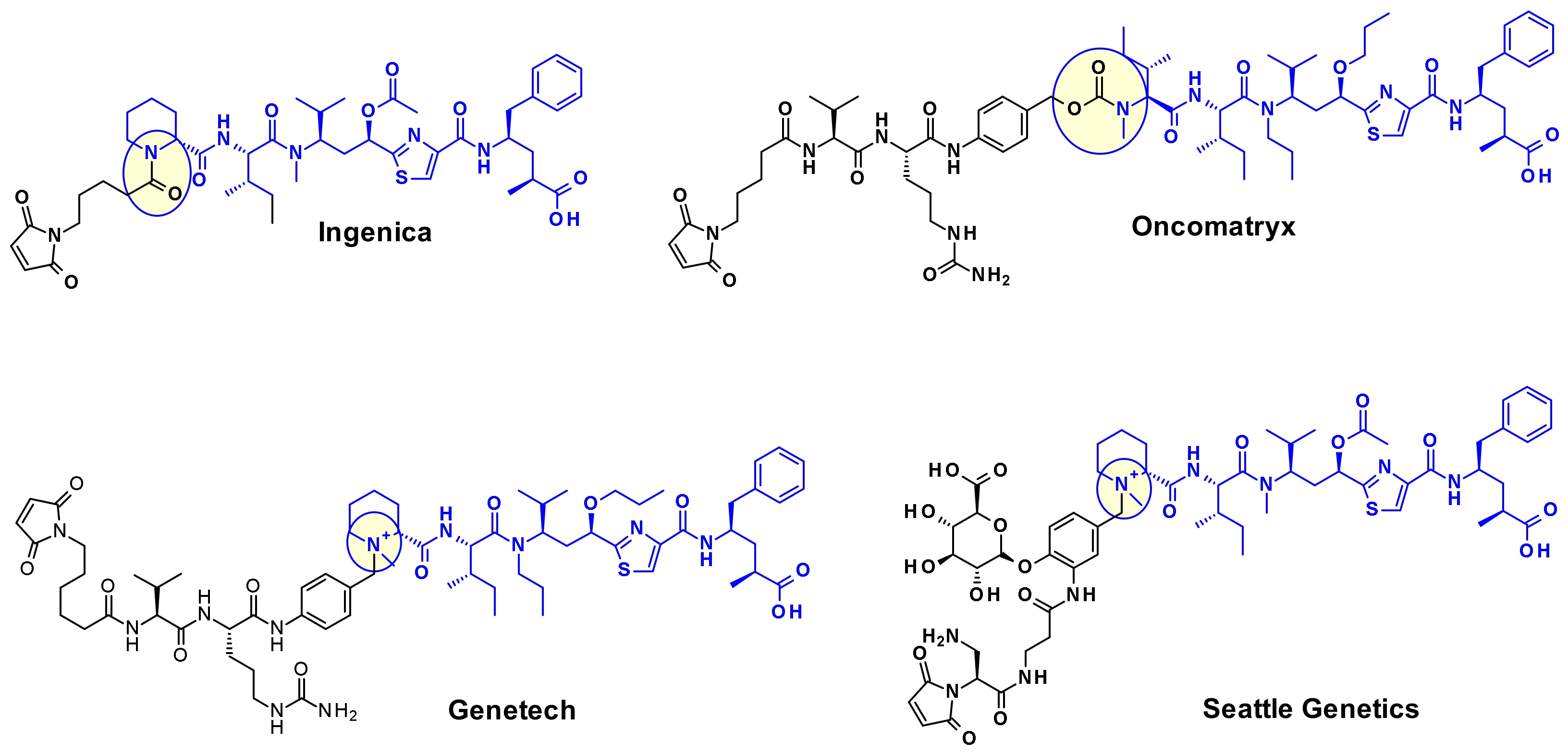














































{kind=link}
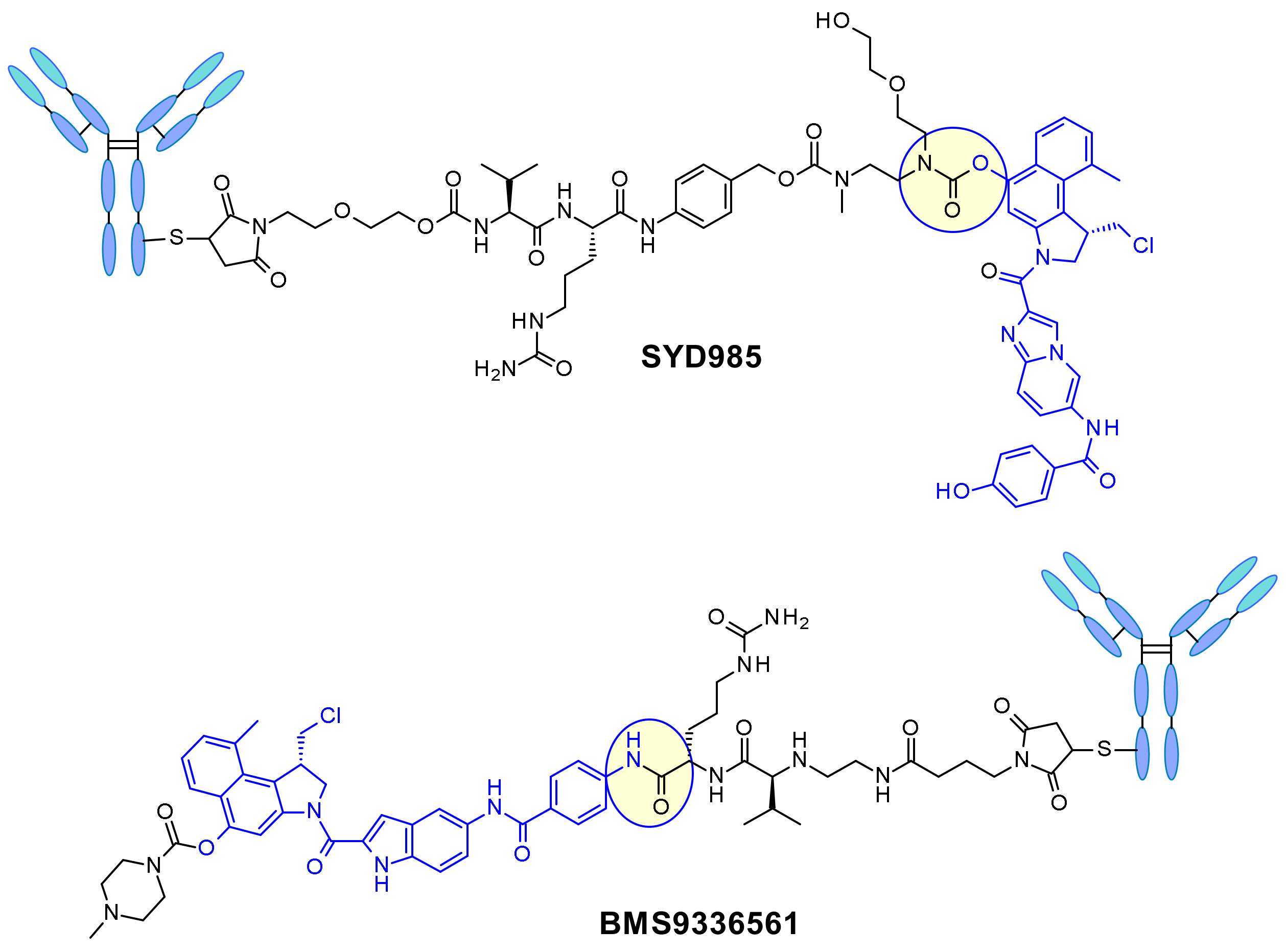
{kind=link}
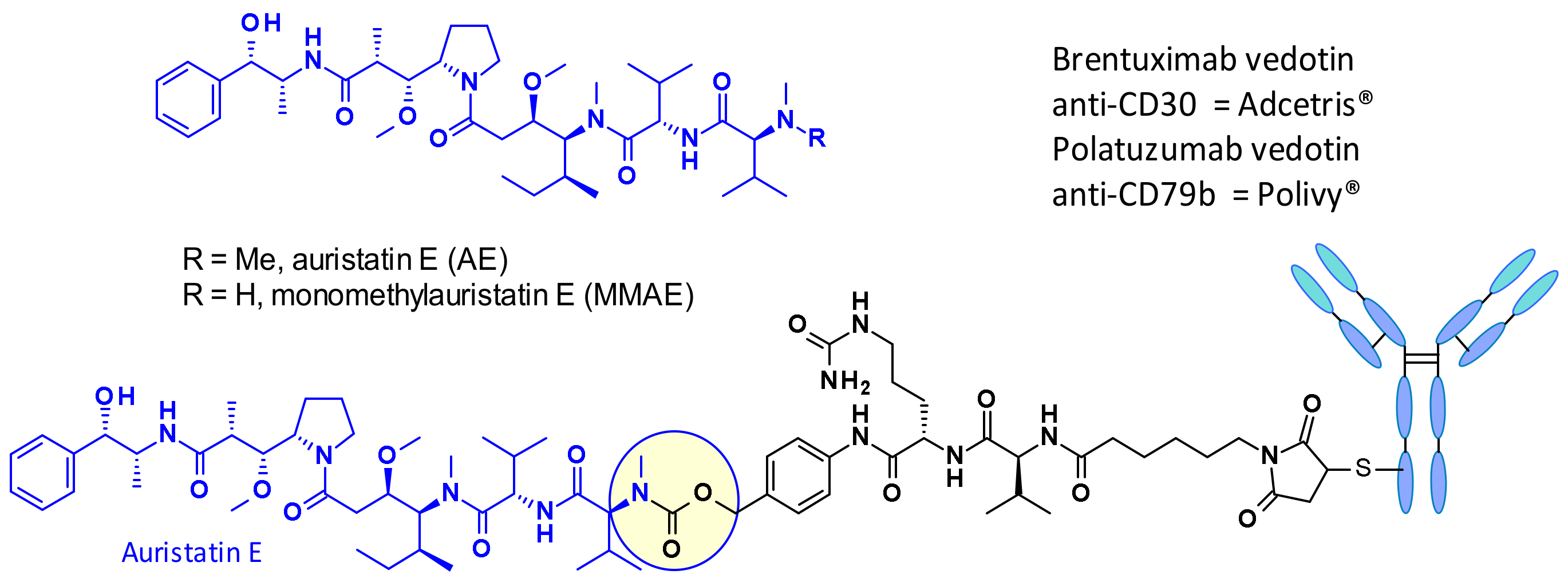
{kind=link}
{kind=link}
{kind=link}
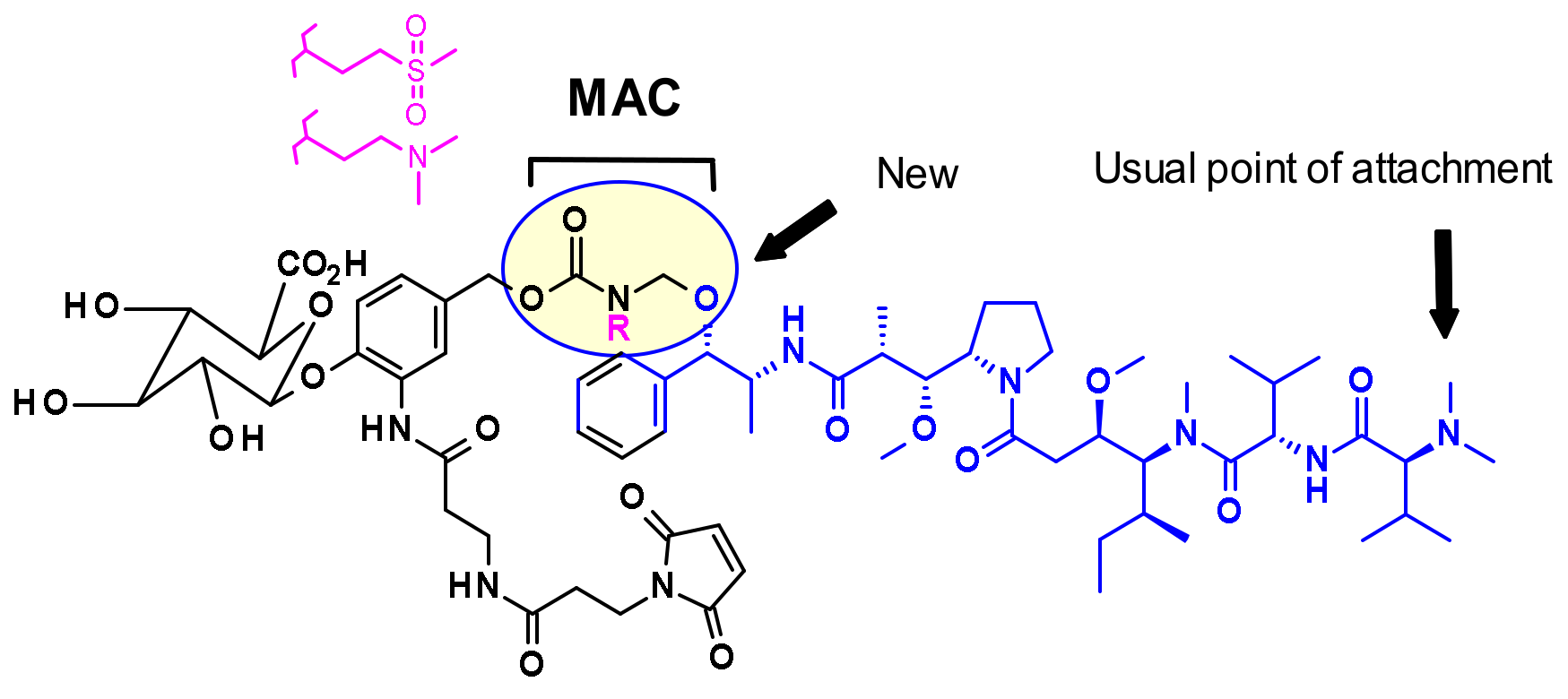{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ADC | Target | Payload | Linker | Indication |
|---|---|---|---|---|
| gemtuzumab ozogamicin (Mylotarg, 1) | CD33 | Calicheamicin | Cleavable, Hydrazone | Acute myeloid leukemia |
| brentuximab vedotin (Adcetris, 2) | CD30 | MMAE 1 | Cleavable, Peptide | Hodgkin leukemia; Systemic anaplastic large-cell lymphoma |
| trastuzumab emtansine (Kadcyla, 3) | HER2 | DM1 | Non-cleavable, Thioether | Breast cancer |
| inotuzumab ozogamicin (Besponsa, 4) | CD22 | Calicheamicin | Cleavable, Hydrazone | B-cell Acute lymphocytic leukemia |
| polatuzumab vedotin (Polivy, 5) | CD79b | MMAE | Cleavable, Peptide | Diffuse large B-cell lymphoma |
| enfortumab vedotin (Padcev, 6) | Nectin-4 | MMAE | Cleavable, Peptide | Urothelial cancer |
| trastuzumab deruxtecan (Enhertu, 7) | HER2 | deruxtecan | Cleavable, Peptide | Breast cancer |
| sacituzumab govitecan (TRODELVY, 8) | TROP-2 | SN-38 | Cleavable, Peptide | Breast cancer |
| belantamab mafodotin (BLENREP, 9) | BCMA | MMAF | Non-cleavable, Thioether? | Multiple myeloma |
| moxetumomab pasudotox (Lumoxiti, 10) | CD22 | PE38 | Fusion protein | Hairy cell leukemia |
| tagraxofusp (Elzonris, 11) | IL-3 | Diphteria toxin | Fusion protein | Blastic plasmacytoid dendritic cell neoplasm |
| ibritumomab tiuxetan (ZEVALIN, 12) | CD20 | 90-yttrium | tiuxetan | Multiple hematological disorders |
 | ||||
|---|---|---|---|---|
| R1 | R2 | R3 | R4 | |
| AstraZeneca |  | Et | Ac | (CH2)4NH2 |
| Bristol-Myers Squibb |  | nPr | Me | (CH2)4NH2CONH2 |
| Pfizer |  | Me | CONHEt | H |
| ADC Name | Target | Payload | Linker | Phase |
|---|---|---|---|---|
| BAT8001 | HER-2 | Maytansinoid | non-cleavable | 3 |
| disitamab vedotin | HER-2 | MMAE | Val-Cit-PABA | 3 |
| DS-1062a | TROP-2 | DXd | Gly-Gly-Phe-Gly | 3 |
| loncastuximab Tesirine | CD19 | SG3199 | Val-Ala-PABA | 3 |
| mirvetuximab soravtansine | FOLR-α | DM4 | sulfo-SPDB 1 | 3 |
| ZRC-3256 2 | HER-2 | DM1 | SMCC 3 | 3 |
| ANG1005 4 | LRP1 | Paclitaxel | covalent | 3 |
| SAR408701 | CEACAM5 | DM4 | SPDB 5 | 3 |
| TAA013 | HER-2 | DM1 | SMCC 3 | 3 |
| trastuzumab duocarmazine | HER-2 | seco-DUBA | Val-Cit-PABA | 3 |
| tisotumab vedotin | TF | MMAE | Val-Cit-PABA | 3 |
| ARX788 | HER-2 | Amberstatin269 | oxime | 2/3 |
| ABBV-3373 | TNF-α | Steroid | Ala-Ala-PABA | 2 |
| anetumab ravtansine | MSLN | DM4 | SPDB 5 | 2 |
| BA3011 | Axl | MMAE | cleavable | 2 |
| camidanlumab Tesirine | CD25 | SG3199 | Val-Ala-PABA | 2 |
| labetuzumab Govitecan | CEACAM5 | SN-38 | CL2A 6 | 2 |
| ladiratuzumab vedotin | LIV-1 | MMAE | Val-Cit | 2 |
| MRG003 | EGFR | MMAE | Val-Cit-PABA | 2 |
| naratuximab emtansine | CD37 | DM1 | SMCC 3 | 2 |
| patritumab Deruxtecan | HER-3 | DXd | Gly-Gly-Phe-Gly | 2 |
| praluzatamab ravtansine | CD166 | DM4 | SPDB 5 | 2 |
| telisotuzumab vedotin | c-MET | MMAE | Val-Cit-PABA | 2 |
| VLS-101 | ROR1 | MMAE | Val-Cit-PABA | 2 |
| ADC Name | Target | Payload | Linker | Phase |
|---|---|---|---|---|
| cintredekin besudotox 1 | IL-13R | Pseudomonas exotoxin A | fusion | 3 |
| E7777 2 | IL-2R | Diptheria Toxin A,B | fusion | 3 |
| oportuzumab monatox 3 | EpCAM | Pseudomonas exotoxin A | fusion | 3 |
| T-Guard 4 | CD7 and CD3 | Ricin A | undisclosed | 3 |
| naptumomab estafenatox 5 | 5T4 | Staphylococcal Enterotoxin E | fusion | 2/3 |
| Proxinium 6 | EpCAM | Pseudomonas exotoxin A | fusion | 2/3 |
| EP-100 7 | LHRH | CLIP-71 | fusion | 2 |
| L-DOS47 8 | CEACAM6 | Urease | SIAB | 2 |
| LMB-2 9 | CD25 | Pseudomonas exotoxin | fusion | 2 |
| MDNA55 10 | IL-4R | Pseudomonas exotoxin A | fusion | 2 |
| MT-3724 11 | CD20 | Shiga-like toxin A | fusion | 2 |
| Resimmune 12 | CD3 | Diphtheria toxin | fusion | 2 |
| RO6927005 13 | MSLN | Pseudomonas exotoxin A | fusion | 2 |
| ADC Name | Target | Payload | Linker | Phase |
|---|---|---|---|---|
| TLX250-CDx | carbonic anhydrase IX | 89-Zr | DOFA 1 | 3 |
| IMMU-107 | MUC-1 | 90-Y | DOTA 2 | 3 |
| Iomab-B | CD45 | 131-Iodine | direct iodination | 3 |
| 131I-8H9 | B7-H3 | 131-Iodine | direct iodination | 2/3 |
| 111In-J591 | PSMA | 111-In | DOTA 2 | 2 |
| 177Lu-DOTA-girentuximab | carbonic anhydrase IX | 177-Lu | DOTA 2 | 2 |
| 177Lu-DOTA-Rosopatamab | FOLH1 | 177-Lu | DOTA 2 | 2 |
| Betalutin | CD37 | 177-Lu | p-SCN-Bn-DOTA | 2 |
| CLR 131 3 | Lipid raft | 131-I | direct iodination | 2 |
| 64Cu-DOTA-trastuzumab | HER-2 | 64-Cu | DOTA 2 | 2 |
| 111 In-ibritumomab tiuxetan | CD20 | 111-In | modified-DTPA | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostova, V.; Désos, P.; Starck, J.-B.; Kotschy, A. The Chemistry Behind ADCs. Pharmaceuticals 2021, 14, 442. https://doi.org/10.3390/ph14050442
Kostova V, Désos P, Starck J-B, Kotschy A. The Chemistry Behind ADCs. Pharmaceuticals. 2021; 14(5):442. https://doi.org/10.3390/ph14050442
Chicago/Turabian StyleKostova, Vesela, Patrice Désos, Jérôme-Benoît Starck, and Andras Kotschy. 2021. "The Chemistry Behind ADCs" Pharmaceuticals 14, no. 5: 442. https://doi.org/10.3390/ph14050442
APA StyleKostova, V., Désos, P., Starck, J.-B., & Kotschy, A. (2021). The Chemistry Behind ADCs. Pharmaceuticals, 14(5), 442. https://doi.org/10.3390/ph14050442