Abstract
Ulcerative colitis (UC) is an inflammatory ailment of the intestine associated with the upregulation of oxidative stress and pro-inflammatory cytokines. Here, we aimed to assess the consequences of Encephalartos villosus (EV) Lem extract on acetic acid (AA)-induced UC. Rats were randomly classified into five groups, as follows: control, AA, AA + mesalazine, AA + EV (50 mg/kg), and AA + EV (100 mg/kg) groups. EV (50 mg/kg and 100 mg/kg) and mesalzine (100 mg/kg) were administered orally for 14 days before the induction of UC. On the last day of the experiment, colitis was provoked via the intra-rectal delivery of 3% AA. Then, after 24 h, the rats were sacrificed and their colon tissues were isolated and inspected. Interestingly, EV pretreatment substantially (p < 0.05) reduced the elevated colon weight/length ratio and ulcer area and normalized the histological changes and immunohistochemical features. In addition, EV efficiently reduced the levels of myeloperoxidase (MPO) and increased the activity of glutathione peroxidase (GS-PX) and catalase (CAT). EV (100 mg/kg) resulted in a downregulation of toll-like receptor 4 (TLR-4) and upregulation of heme oxygenase 1 (HO-1) and occludin expression levels. Concerning the anti-inflammatory mechanisms, EV reduced the levels of tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and nuclear transcription factor kappa B (NF-ĸB) and inhibited cyclooxygenase-2 (COX-2) expression levels. It also decreased caspase-3 levels. Our results indicate that the oral intake of EV improves AA-induced colitis in rats through its antioxidative effects and the modulation of pro-inflammatory cytokines, as well as the restoration of mucosal integrity. Consequently, EV may be an efficient therapeutic candidate for UC.
1. Introduction
Ulcerative colitis (UC) and Crohn’s disease (CD) are ailments of the gastrointestinal tract (GIT). Repeated and non-specific inflammation of the intestine usually occurs in these diseases [1,2]. It is hypothesized that the interaction between the environment and genes triggers an immunological response in the mucosa of the intestine, provoking remarkable inflammation, as well as injury, in the GIT [3].
Toll-like receptors (TLRs) are components of the immune system that play an important role in intestinal inflammation [4,5]. There are many toll-like receptor subtypes, and TLR-4 is one of these subtypes, which plays a major role in inflammatory bowel diseases (IBD). In these conditions, the level of TLR-4 increases [4]. As a result of this increase, upregulation typically occurs in the downstream signaling pathways, including the nuclear factor binding kappa light-chain (NF-ĸB) signaling pathway [6]. As a result, this prompts the release of different pro-inflammatory cytokines, like tumor necrosis factor-alpha (TNF-α) [7].
NF-kB has an important role in the pathogenesis of UC by triggering the release of inflammatory mediators, like cyclooxygenase-2 (COX-2) [8,9]. COX-2 then induces the release of prostaglandins through the metabolism of arachidonic acid [10].
The activation of inducible nitric oxide synthase (iNOS), as well as COX-2, usually results in the destruction of intestinal mucosa by inducing the production of free radicals and inhibiting the anti-oxidative system [11,12]. Indeed, the excessive production of reactive oxygen species (ROS) negatively affects the defense system of the mucosa [13].
The increased release of pro-inflammatory mediators, like TNF-α, can trigger apoptosis, which has an important role in animal colitis models. It is reported that apoptosis is associated with the pathophysiology of IBD [14,15]. The inflammatory response changes the function of the mucosal barrier and affects intestinal integrity, leading to the induction of apoptosis [16]. Consequently, finding novel alternatives that inhibit the inflammatory cascade is a favorable approach to improving the pathological consequences of IBD, which is an inflammatory-mediated ailment [17].
Encephalartos villosus Lem. is a decorative dwarf cycad often referred to as the poor man’s cycad. A previous phytochemical study on E. villosus leaves documented the isolation of four flavone glycosides: luteolin-7-glucoside, luteolin-7-rutinoside, apigenin-7-glucoside, and luteolin-7-rhamnoside. This plant exhibited promising anti-inflammatory effects [18]. Therefore, we aimed to elucidate the anti-inflammatory potential of EV in acetic-acid-induced colitis in rats and to explore the potential underlying mechanisms of such consequences.
2. Results
2.1. HPLC Analysis
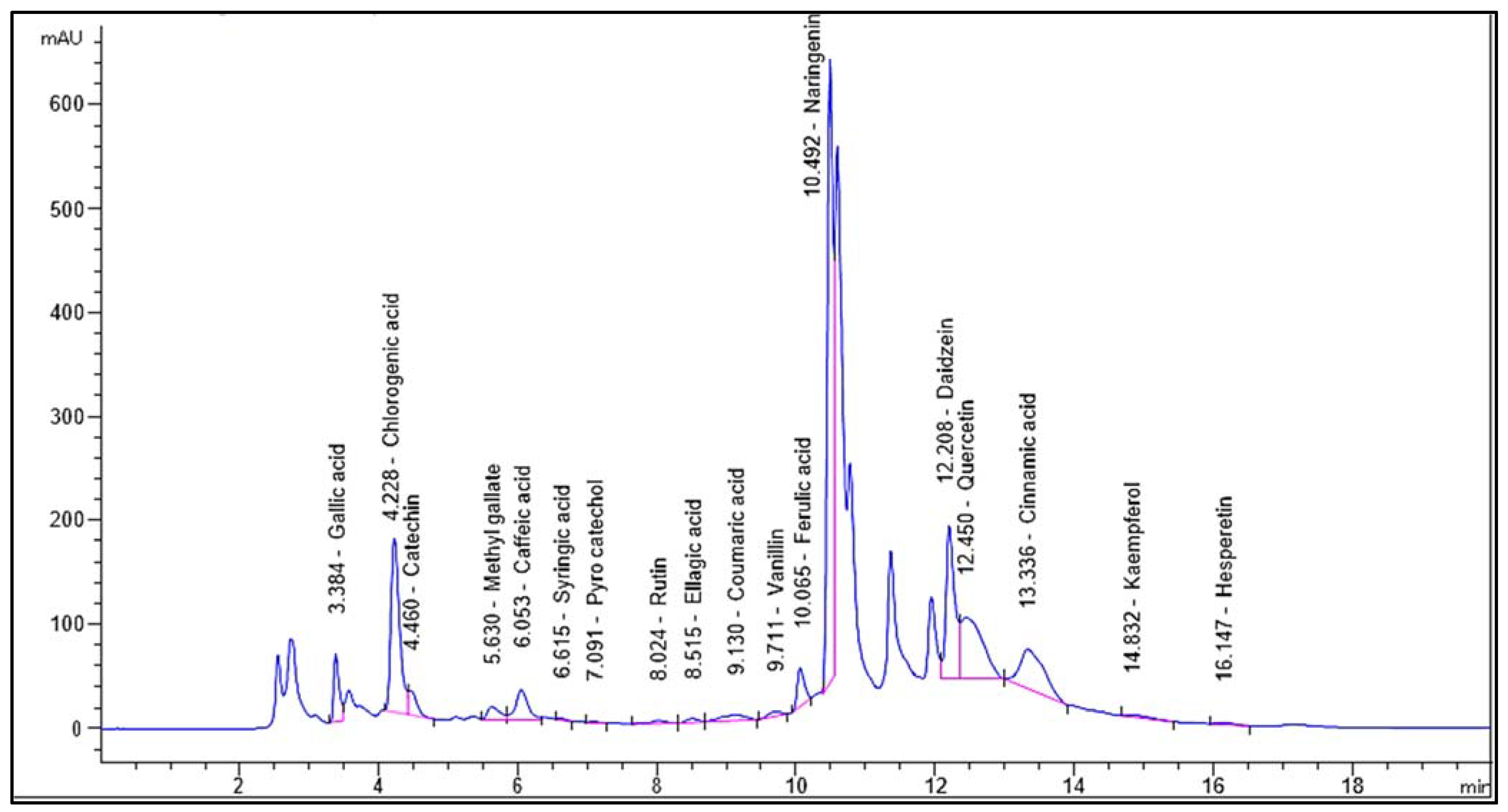
Identification and quantification of the chemical composition of Encephalartos villosus extract was performed using HPLC against standard compounds (Figure S1). Figure 1 shows the HPLC chromatogram for the identified flavonoids and phenolic compounds in the Encephalartos villosus extract. The HPLC analysis identified 18 compounds (Table 1). The most abundant phenolic compound was chlorogenic acid (7595.27 μg/g), and the major flavonoid was naringenin (11,476.89 μg/g).
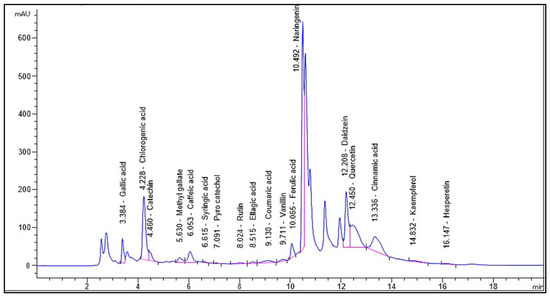
Figure 1.
Chromatogram for the identified flavonoids and phenolic compounds of Encephalartos villosus.

Table 1.
Chemical composition analysis of the phenolic and flavonoid compounds in Encephalartos villosus by HPLC.
2.2. Colon Weight
AA group displayed a noteworthy increase in colon weight (24.82%) compared to the control group. Mesalazine and EV 100 displayed a pronounced reduction (p < 0.05) in the colon weight (24.96 and 20.87%, respectively) compared to the AA group. The effect was non-significant in the EV 50 group (Table 2).
Table 2.
Influence of EV treatment on colon weight, colon length, and colon weight/length ratio in AA-induced UC in rats.
2.3. Colon Length
The acetic acid (AA) group displayed a noteworthy reduction in colon length (21.7%) compared to the control group. Mesalazine and EV 100 presented a substantial rise (p < 0.05) in colon length (18.81 and 23.76%, respectively) compared to the AA group. EV 50 presented no substantial change (4.95%) compared to the AA group (Table 2).
2.4. Colon Weight/Length Ratio
Table 2 reveals that the AA group had a substantial rise in the colon weight/length ratio (60.68%) compared to the control group. Mesalazine, EV 50, and EV 100 showed a substantial decline (p < 0.05) in the colon weight/length ratio (37.12, 17.89, and 36.5%, respectively) when compared to the AA group, with a more pronounced effect in the EV 100 group compared to the EV 50 group (Table 2).
2.5. Colon Catalase (CAT) Activity
The AA group displayed a significant decrease (p < 0.05) in colonic CAT activity (30.93%) compared to the control group. Treatment with mesalazine and EV 100 successfully restored CAT activity (19.7, 7.76, and 24.65%, respectively) compared to the AA group. Moreover, EV 100 showed a more substantial increase (p < 0.05) in CAT activity compared to the AA group (24.65%) (Figure 2A).
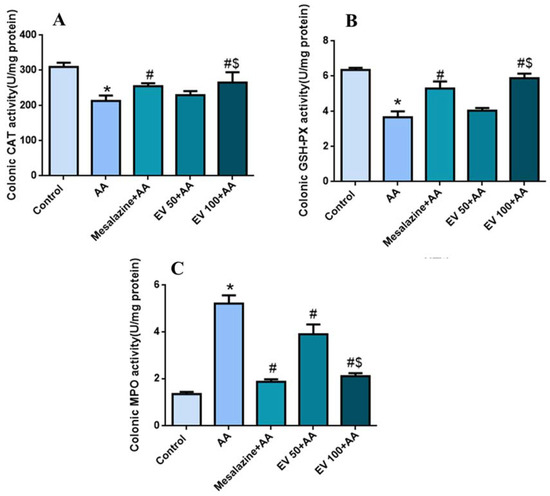
Figure 2.
Influence of EV extract on colonic (A) CAT (B) GSH-PX (C) MPO activity in AA-induced UC. Significant difference vs. * respective control, # respective AA group, and $ respective EV 50 group. p < 0.05.
2.6. Colon Glutathione Peroxidase (GSH-PX) Activity
Figure 2B shows a marked reduction in colonic GSH-PX activity in the AA group (42.2%) compared to the control group. The mesalazine-treated group effectively reestablished GSH-PX activity (44.68%) compared to the AA group. EV 50 and EV 100 also induced a substantial increase (p < 0.05) in GSH-PX activity compared to the AA group (10.35 and 60.21%, respectively) (Figure 2B); p < 0.05.
2.7. Colon Myeloperoxidase (MPO) Activity
As shown in Figure 2C, the AA group showed a substantial escalation (p < 0.05) in MPO activity (289.55%) compared to the control group. Mesalazine, EV 50, and EV 100 showed a substantial decline (p < 0.05) in MPO activity (64.17, 25.28, and 59.57%, respectively) compared to the AA group. MPO activity in the EV 100 group was significantly reduced (88.10%) (Figure 2C).
2.8. Colon Gene Expression of TLR-4, Hemoxygenase (HO-1), and Occludin
The AA group displayed a noteworthy rise (p < 0.05) in colonic TLR-4 gene expressions (110%) compared to the control group. Mesalazine, EV 50, and EV 100 induced a noteworthy decline (p < 0.05) in TLR-4 expression (33.33, 9.5, and 38%, respectively) compared to the AA group. There was no substantial alteration in the TLR-4 expression level in the EV 50 group (6.52%) compared to the AA group (Figure 3A).
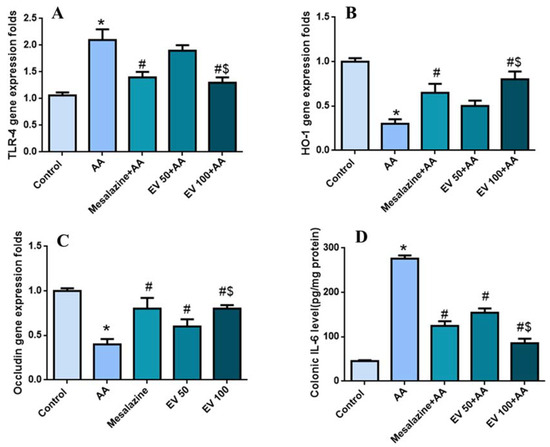
Figure 3.
Influence of EV extract on colonic (A) TLR-4 gene expression, (B) HO-1 gene expression, (C) occludin expression, and (D) IL-6 level in AA-induced UC. Significant difference vs. * respective control, # respective AA group, and $ respective EV 50 group; p < 0.05.
Also, in Figure 3B, the AA group revealed a noteworthy decline in colonic HO-1 gene expressions (233%) compared to the control group. Mesalazine, EV 50, and EV 100 considerably boosted (p < 0.05) the HO-1 expression levels (100, 66.6, and 166.6%, respectively) compared to the AA group. (Figure 3B).
Figure 3C shows that AA persuaded a remarkable injury in the tight junction barrier expressed by a substantial decline (p < 0.05) in the colonic occludin gene expressions (130%) compared to the control group. Mesalazine, EV 50, and EV 100 considerably restored (p < 0.05) the occludin expression levels (100, 50, and 100%, respectively) compared to the AA group (Figure 3C).
2.9. Colon Interleukin-6 (IL-6) Level
The AA group exhibited a substantial rise (p < 0.05) in IL-6 level (508.86%) compared to the control group. Mesalazine treatment considerably diminished (p < 0.05) the colonic IL-6 level (54.89%) compared to the AA group. Furthermore, the EV 50 and EV 100 groups also showed a noteworthy decline in the IL-6 level (44.2 and 68.98%, respectively) compared to the AA group (Figure 3D).
2.10. Histopathological Examination
The histological features of the colon of the tested groups are exposed in Figure 4.
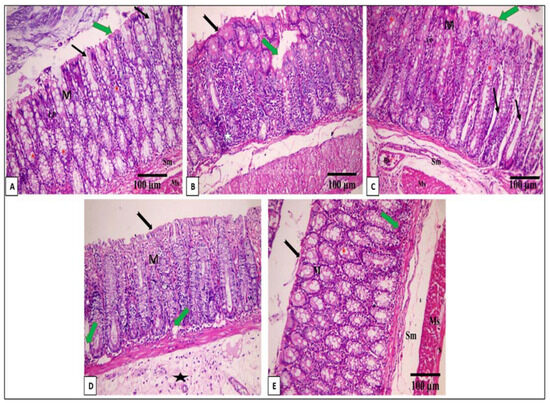
Figure 4.
A photomicrograph of a section in the distal colon of an adult male albino rat: (A) control group I reveals mucosa (M), submucosa (Sm), and musculosa (Ms). Lamina propria (LP) of the mucosa contains closely packed crypts of Lieberkuhn (black arrow). The mucosa is lined by the surface epithelium with a brush border (green arrow) and crypts with abundant goblet cells (*) (H & E × 200, scale bar = 100 μm). (B) Sections in the distal colon of an adult male albino rat of the AA group showing focal loss of surface epithelium and part of the lamina propria (green arrow) detachment of the surface epithelium in some areas (black arrow) in addition to dense inflammatory cellular infiltration (star) (H & E × 200, scale bar = 100 μm). (C) Mesalazine group reveals normal histological structure of the colon as control group I: (H & E × 200, scale bar = 100 μm). (D) The EV 50 group shows apparently normal mucosa (M), intact surface epithelium (black arrow), and focal inflammatory cellular infiltrations with bleeding in lamina propria. Moreover, there are crypt cysts (green arrow), a mildly decreased number of goblet glands, and an increase in the thickness of the submucosa (star) (H & E × 200, scale bar = 100 μm). (E) The EV 100 group reveals the regaining of the normal appearance of the colon in the form of mucosa (M), submucosa (Sm), and musculosa (Ms). Lamina propria (LP) of the mucosa with few inflammatory cellular infiltrations (green arrow). The mucosa is lined by the surface epithelium, which has a brush border (black arrow), and crypts have abundant goblet cells (*) (H & E × 200, scale bar = 100 μm).
2.11. Immunohistochemical Features of Caspase-3, NF-kB, TNF-α, and COX-2
Caspase-3, NF-kB, TNF-α, and COX-2 immunohistochemical features of the colon of the different experimental groups are shown in Figure 5, Figure 6, Figure 7 and Figure 8.
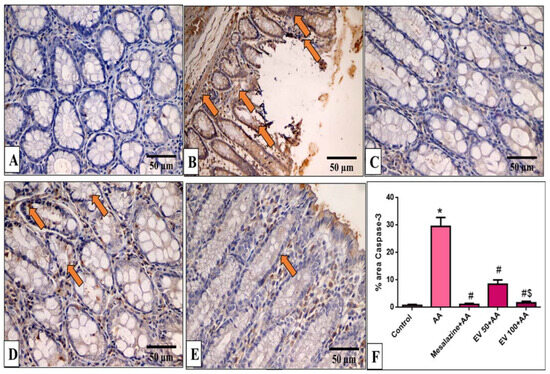
Figure 5.
A photomicrograph of sections in the distal colon of adult male albino rats of various studied groups stained by caspase-3. (A) The normal control group shows a colonic gland with no caspase-3 immune reaction in all cells. (B) The AA group shows a strong positive caspase-3 immune reaction in epithelial and glandular cells (arrows). (C) The mesalazine group shows no caspase-3 immune reaction in all cells. (D) The EV 50 group IV moderate caspase-3 immune reaction in glandular cells (arrows). (E) The EV 100 group shows no caspase-3 immune reaction in the majority of cells and a weak immune reaction in a few surface epithelial cells (caspase-3 immune reaction in cells immunostaining × 400, scale bar = 50 μm). (F) Percent area of caspase-3-positive cells/1000 cells. Significant difference vs. * respective control, # respective AA group, $ respective EV 50 group, each at p < 0.05.
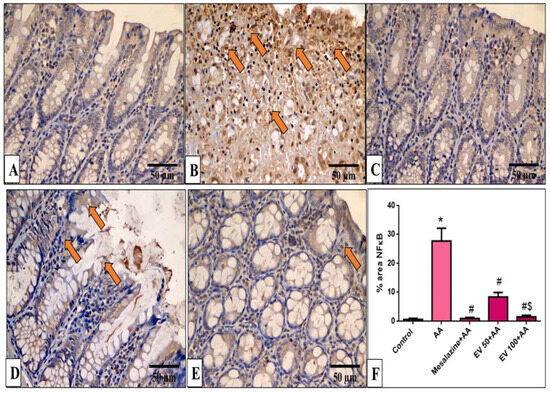
Figure 6.
A photomicrograph of sections in the distal colon of adult male albino rats of various studied groups stained by NF-κB. (A) The normal control group shows a colonic gland with no NF-κB immune reaction in all cells. (B) The AA group shows a strong positive NF-κB immune reaction in surface epithelial and glandular cells (arrows). (C) The mesalazine group shows no NF-κB immune reaction in all cells. (D) The EV 100 group shows a moderate NF-κB immune reaction in surface epithelial cells (arrows). (E) The EV 100 group shows no NF-κB immune reaction in the majority of cells and a weak immune reaction in a few cells. (NF-κB immune reaction in cells immunostaining × 400, scale bar = 50 μm). (F) Percent area of NF-κB-positive cells/1000 cells. Significant difference vs. * respective control, # respective AA group, $ respective EV 50 group, each at p < 0.05.
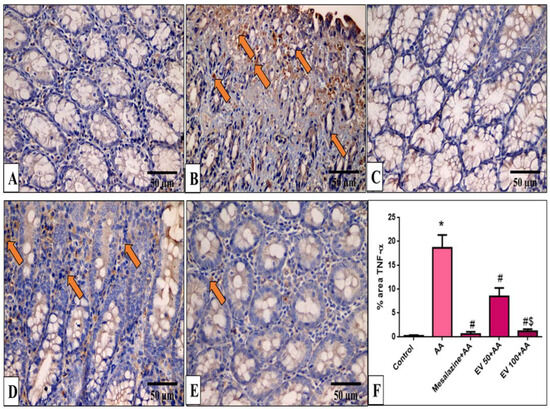
Figure 7.
A photomicrograph of sections in the distal colon of adult male albino rats of various studied groups stained by TNF-α. (A) The normal control group shows a colonic gland with no TNF-α immune reaction in all cells. (B) The AA group shows a strong positive TNF-α immune reaction in surface epithelial and glandular cells (arrows). (C) The mesalazine group shows no TNF-α immune reaction in all cells. (D) The EV 50 group shows a moderate TNF-α immune reaction in some cells (arrows). (E) The EV 100 group shows no TNF-α immune reaction in the majority of cells and a weak immune reaction in a few cells (TNF-α immune reaction in cells immunostaining × 400, scale bar = 50 μm). (F) Percent area of TNF-α-positive cells/1000 cells. Significant difference vs. * respective control, # respective AA group, $ respective EV 50 group, each at p < 0.05.
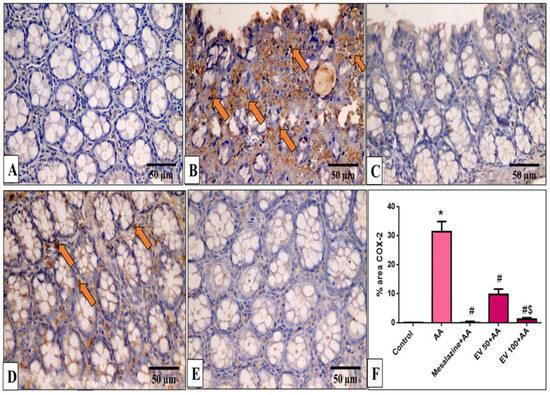
Figure 8.
A photomicrograph of sections in the distal colon of adult male albino rats of various studied groups stained by COX-2 immunostaining. (A) The normal control group shows no COX-2 immune reaction in all cells. (B) The AA group shows a strong positive COX-2 immune reaction in surface epithelial and glandular cells (arrows). (C) The mesalazine group shows no COX-2 immune reaction in all cells. (D) The EV 50 group shows a moderate immune reaction in some glandular cells (arrows). (E) The EV 100 group shows no COX-2 immune reaction in all cells (COX-2 immunostaining × 400, scale bar = 50 μm). (F) Percent area of COX-2-positive cells/1000 cells. Significant difference vs. * respective control, # respective AA group, $ respective EV 50 group, each at p < 0.05.
Results of immune-staining quantification revealed that the AA group showed strong Capase-3 immunostaining (41.04-fold increase). EV 50- and EV 100-treated groups substantially reduced caspase-3 immunostaining by 71.39% and 94.3%, respectively, relative to the AA group (Figure 5F, p < 0.05).
In Figure 6F, the AA group significantly increased NF-κB immunostaining (41.3-fold increase). EV 50 and EV 100 treatments significantly decreased NF-ĸB immunostaining by 69.9% and 94.34%, respectively, with a more pronounced effect in the EV 100 group (Figure 6F, p < 0.05).
Also, as displayed in Figure 7F, the AA group induced a pronounced increase in TNF-α staining (71.89-fold increase). EV 50 and EV 100 treatment showed a significant reduction in TNF-α immunostaining by 54.44% and 93.5% compared to AA (Figure 7F, p < 0.05).
Results indicated that the AA group induced a marked increase in COX-2 staining (232-fold increase). EV 50 and EV 100 pre-treated groups significantly suppressed COX-2 immunostaining by 68.86% and 95.66% compared to AA (Figure 8F, p < 0.05).
3. Discussion
UC is a repeated colon inflammation characterized by the infiltration of inflammatory cells and expression of NF-kB-dependent pro-inflammatory biomarkers like TNF-α and IL-6 [19,20]. Also, ROS are released in this inflammatory disease. In addition, the mucosal integrity of the colon is usually lost [14,21]. Despite the great efforts that are currently being made to cure UC, it is still a poorly treatable ailment. This is in addition to the high risk of its progression to colon cancer. Thus, it is important to find novel therapies to obtain better clinical results.
Here, the AA-induced UC model was utilized to reveal the potential consequences of EV on colitis. Remarkably, there was a substantial reduction (p < 0.05) in the colon length and a substantial rise (p < 0.05) in the colon weight and colon weight/length ratio.
There was a vast release of ROS in the colon tissue after AA administration, and this is attributed to the destruction of the mucosa. This is demonstrated by the considerable reduction of the antioxidant enzymes (CAT and GSH-PX). EV extract-treated groups significantly replenished the capacity of antioxidant enzymes and alleviated oxidative stress, and this is consistent with a previous report [22].
According to the reported IBD pathophysiology, TLRs mediate innate protection in the bowel. Inappropriate levels of TLRs result in stimulation of pro-apoptotic signaling pathways and chronic inflammation [23,24]. Activation of TLR4 causes a rise in NF-kB expression (TLR4/NF-kB pathway) and, as a result, a vast release of inflammatory cytokines and enzymes like IL-1, IL-6, TNF-α, and COX-2 usually occurs, which have a major role in IBD [25,26]. So, it has been proposed that the TLR4/NF-ĸB pathway could be an objective for the treatment of IBD. Here, we found that AA-induced colitis has led to a rise in the expression of TLR4 by qRT-PCR and NF-kB immunostaining, which is in line with previous studies [27,28,29].
The EV 100-treated group displayed a pronounced decrease in TLR-4 expression and NF-kB immunostaining. HPLC of EV identified 18 compounds belonging to flavonoids and polyphenolic subclasses. This investigation revealed that EVME possesses antimicrobial and anti-inflammatory activities, which are likely due to the presence of several active constituents, such as phenolics, flavonoids, and glycoside derivatives. Naringenin is the most abundant flavonoid metabolite, followed by daidzein and quercetin [22,30].
Naringenin was reported to significantly improve colitis in dextran sulfate sodium (DSS)-induced UC in mice by impeding NF-κB and TLR4 protein activity [31,32]. Also, the expressions of iNOS, COX-2, TNF-α, and IL-6 were downregulated [33,34]. Since naringenin is the major compound in EV extract, this may explain its inhibitory effects on the TLR-4/NF-kB axis and ameliorating impact on AA-induced colitis. It has been revealed that flavonoids, as well as polyphenols, enhanced the therapeutic outcomes in the experimental colitis via decreasing colonic injury and inflammation [27]. All such reports established the anti-inflammatory potential of EV compounds and strengthened their use as an anti-inflammatory candidate in AA-induced UC [34].
In accordance with the preceding investigations, this study revealed increased levels of TNF-α immunostaining after AA-colitis induction and augmented levels of IL-6 [23]. TNF-α, in turn, recruits neutrophils toward the tissues. The MPO level was measured to indicate neutrophil accumulation [35,36]. It was noticed that there was a noteworthy rise in MPO activity after UC induction. Interestingly, the MPO activity was considerably declined by EV, which was demonstrated for the first time and may be partially explained by its constituents, which exhibit marked anti-inflammatory properties and a reduction in TNF-α levels in the colon [22,37]. Additionally, mesalazine resulted in a noteworthy decrease in MPO activity. Such findings are in line with previous studies [38].
As a result of the activity of COX-2, both prostaglandin E2 (PGE2) and thromboxane B2 are produced, which, in turn, induce intestinal hyperemia and edema [39]. Thus, activation of COX-2 could lead to an increase in ROS/RNS and a depletion of the antioxidative system. All of these consequences could lead to the induction of apoptotic injuries in the epithelium of the colon [40,41]. Here, AA-induced UC demonstrates an increase in COX-2 immunostaining. EV treatment significantly decreased the COX-2 immunostaining, and this could be due to inhibition of the ROS/RNS and reestablishment of the antioxidative activity. Our phytochemical investigation was in line with other previous research [37,42,43,44]. All of these phytochemicals revealed pronounced antioxidant and anti-inflammatory potentials, which might explain the beneficial impacts of EV extract.
Previous investigations found that apoptosis is associated with the pathophysiology of IBD [45]. The inflammatory response affects the function of the mucosal barrier and the intestinal integrity, which often leads to apoptosis [46,47,48]. In the current study, caspase-3 immunostaining was expressively expressed. Also, the expression of the tight junction protein, occludin, was reduced in the colon tissues of the AA-induced UC group. EV 100 treatment diminished caspase-3 immunostaining and upregulated occludin gene expression. Thus, the apoptotic injuries were prevented, and the intestinal integrity was maintained, which preserved the functions of the mucosal barriers. Thus, EV produced significant antiapoptotic impacts by diminishing the discharge of ROS and NO.
HO-1 is an antioxidant enzyme present at low levels in different normal tissues. Exposure to oxidants interrupts the transcription of antioxidant proteins like HO-1 and CAT [49,50,51]. HO-1 has immunomodulatory, antiproliferative, anti-inflammatory, and antiapoptotic effects [52]. Therefore, in our study, HO-1 expression levels were markedly suppressed in the AA-induced UC. EV-treated groups significantly replenished HO-1 expression levels, confirming EV extract’s anti-inflammatory, antioxidant, and antiapoptotic properties. These effects may be due to their flavonoids, such as naringenin, daidzein, kaempferol, and quercetin derivatives, as well as phenolic compounds, such as chlorogenic acid and cinnamic acid. All these bioactive molecules were previously reported for their anti-inflammatory, antioxidant, and antiapoptotic impacts [37,53,54,55].
EV ameliorated the morphological mucosal damage recorded by the histological studies, which is consistent with other studies [23,56]. EV lessened the detrimental impact of AA by refining all the studied factors. Our study is the first investigation indicating the possible ameliorative properties of EV on AA-induced UC in rats. All these effects might prove the plausible protective action of Encephalartos villosus on AA-induced UC, which was first demonstrated in this study.
4. Materials and Methods
4.1. Animals
Fifty male Wistar albino rats with weights ranging from 150 to 170 g were obtained from Cairo University (Egypt). Animals were preserved in standard conditions. The experimental techniques were accepted by the research ethical committee (Faculty of Pharmacy, Tanta University, Egypt) (TP/RE/6/23 p-0031).
4.2. Plants, Drugs, and Chemicals
Mesalazine was obtained from EL-Pharonia pharmaceutical company (Cairo, Egypt). All other chemicals were gained from Sigma Aldrich (St. Louis, MO, USA).
4.2.1. Encephalartos villosus Extract Preparation
Encephalartos villosus Lem. leaves were collected from Al Orman Botanical Garden on 15 July 2021. Dr. Esraa Ammar, Plant Ecology, Tanta University, recognized the plant. A voucher sample (PG-G-106-E) was deposited at the Herbarium of the Faculty of Science, Tanta University. The plant was dried at room temperature for ten days, then in an oven at 40 °C for two days, and then ground. The powder (300 g) was extracted by methanol (four L × three times) at two-day intervals using the cold maceration method at room temperature to yield 31.39 g of EV extract [22].
4.2.2. High-Performance Liquid Chromatography (HPLC) Analysis
HPLC analysis of the EV is performed following [57] with some modifications. The SCRE was analyzed using an Agilent 1260 series instrument and Eclipse C18 column (4.6 mm × 250 mm i.d., 5 µm). Separation was performed at a flow rate of 0.9 mL/min. The mobile phase consisted of water (reservoir A) and 0.5% trifluoroacetic acid in acetonitrile (reservoir B) at a concentration of 0.1%. The mobile phase was sequentially programmed with a linear gradient as follows: 0 min (82%A); 0–5 min (80%A); 5–8 min (60%A); 8–12 min (60%A); 12–15 min (82%A); 15–16 min (82%A); and 16–20 (82%A). A multi-wavelength UV detector was used for detection at 280 nm. The injection volume for each sample solution was 5 μL. The column temperature was retained at 40 °C.
4.3. Experimental Procedure
4.3.1. Induction of UC
It was provoked on the 15th day by the intra-rectal delivery of 2 mL of 3% AA via a rubber catheter into rats’ rectum under anesthesia. Then, animals were left in a vertical position for one minute after administration to prevent the leakage of AA [58].
4.3.2. Design
Fifty animals were grouped into five categories (n = 10). Group I (control group) administered 2 mL of saline intra-rectally. The remaining groups administered AA intra-rectally and were subdivided into group II (untreated AA group), group III (mesalazine group) receiving 100 mg/kg of mesalazine [59] orally daily for 14 days, group IV (EV 50 group) receiving 50 mg/kg of EV orally daily for 14 days, and group V (EV 100 group) receiving 100 mg/kg of EV orally daily for 14 days [22]. After the last dose, rats fasted for 12 h before intra-rectal delivery of either AA or saline.
4.3.3. Tissue Collection
Rats were anesthetized, and then blood was collected via cardiac puncture. Blood was then centrifuged and utilized for further investigations. Rats were then euthanized, and colons were isolated, washed, and dried using filter papers. Colons were then weighed and stretched to detect the distance between the colorectal junction and the distal rectum end [60]. The colon weight was divided by its length (gm/cm) to calculate the colon weight/length ratio [61].
The colons were allocated into two sectors. One section was utilized for histopathological and immunohistochemistry studies. The other section was used for the biochemical examination.
4.4. Colorimetric Determination of Colon Catalase and Glutathione Peroxidase
CAT and GSH-Px activities were evaluated in the homogenate of colon tissues using CAT and GSH-Px assay kits Cayman (Ann arbor, MI, USA) and Elabscience (Houston, TX, USA), respectively, according to the instructions of the manufacturers.
4.5. Colorimetric Determination of the Colon Myeloperoxidase Activity
MPO is an indicator of neutrophil infiltration. Thus, it was assessed in the colon tissue homogenate using an MPO assay kit (Abcam, Cambridge, UK).
4.6. ELISA for Interlukin-6 Levels
The IL-6 level was evaluated in the colon tissue homogenates by an ELISA kit (Sun Red Biotechnology Co., Shanghai, China). The intensity of the color was detected at 450 nm by an ELISA reader (Labnics, Glasgow, UK).
4.7. Quantitative Real-Time (qRT-PCR) for TLR-4, Heme Oxygenase-1, and Occludin
The extracted total RNA using TRIzols (Life Technologies, Carlsbad, CA, USA) [62] was reverse-transcribed into cDNA using the QuantiTects reverse transcription kit (Qiagen, Hilden, Germany). The cDNA was amplified using the Maximas SYBR green/fluorescein qPCR master mix using the primers listed in Table S1.
4.8. Histopathology of Colon Sections
Colons were fixed in formalin, and they were embedded in paraffin wax, sliced into very thin sections, and stained by hematoxylin and eosin (H&E) [37]. The stained sections were visualized by a light microscope.
4.9. Immunohistochemical Detection of Caspase-3, NF-kB, TNF-α, and COX-2
The immunohistochemical investigations were carried out using the monoclonal antibodies of caspase-3, NF-kB, TNF-α, and COX-2 (Santa Cruz Biotechnology, Dallas, TX, USA), as previously reported [63,64,65]. Immuno-stained slides were image-analyzed using ImageJ software.
4.10. Statistics
Results are revealed as the mean ± standard error of the mean (SEM). An ANOVA test was utilized to investigate the difference between groups at p < 0.05 using Prism version 6 (San Diego, CA, USA).
5. Conclusions
This investigation revealed that EV successfully replenished the damage in the colonic mucosa via the administration of AA. It exerted potent anti-inflammatory, antioxidant, and antiapoptotic impacts. The studied molecular mechanisms for such effects could be due to suppressing the TLR4/NF-kB signaling pathway owing to its active constituents. Additional studies have to be conducted to endorse the clinical use of Encephalartos villosus in ulcerative colitis patients.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ph16101431/s1, Table S1: Primers used and their sequences. Figure S1: Chromatogram for the standard flavonoids and phenolic compounds.
Author Contributions
Conceptualization, E.E., W.A.N. and A.H.E.-K.; methodology, W.A.N. and A.H.E.-K.; software, A.S.A.; validation, M.M.A. and N.G.M.A.; formal analysis, A.S.A.; investigation, A.H.E.-K.; resources, W.A.N.; data curation, E.E.; writing—original draft preparation, W.A.N. and A.H.E.-K.; writing—review and editing, E.E., W.A.N. and A.H.E.-K.; visualization, A.S.A. and M.M.A. and N.G.M.A.; project administration, A.S.A., N.G.M.A. and M.M.A.; funding acquisition, A.S.A. and M.M.A. All authors have read and agreed to the published version of the manuscript.
Funding
The authors extend their appreciation to the Deputyship for Research and Innovation, Ministry of Education in Saudi Arabia, for funding this research work through the project number RI-44-0911.
Institutional Review Board Statement
The experimental techniques were accepted by the research ethical committee (Faculty of Pharmacy, Tanta University, Egypt) (TP/RE/6/23 p-0031).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data is contained within the article and Supplementary Material.
Acknowledgments
The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research work through the project number RI-44-0911.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Molodecky, N.A.; Soon, S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, G.; Pallone, F.; MacDonald, T.T. Emerging immunological targets in inflammatory bowel disease. Curr. Opin. Pharmacol. 2011, 11, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [PubMed]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef]
- Ke, X.; Zhou, F.; Gao, Y.; Xie, B.; Hu, G.; Fang, W.; Peng, J.; Chen, Y.; Sferra, T.J. Qing Hua Chang Yin exerts therapeutic effects against ulcerative colitis through the inhibition of the TLR4/NF-κB pathway. Int. J. Mol. Med. 2013, 32, 926–930. [Google Scholar] [CrossRef]
- Lin, S.; Yin, Q.; Zhong, Q.; Lv, F.-L.; Zhou, Y.; Li, J.-Q.; Wang, J.-Z.; Su, B.-Y.; Yang, Q.-W. Heme activates TLR4-mediated inflammatory injury via MyD88/TRIF signaling pathway in intracerebral hemorrhage. J. Neuroinflamm. 2012, 9, 46. [Google Scholar] [CrossRef]
- Ahmed, S.; Dewan, M.Z.; Xu, R. Nuclear factor-kappaB in inflammatory bowel disease and colorectal cancer. Am. J. Dig. Dis 2014, 1, 84–96. [Google Scholar]
- Laurindo, L.F.; Santos, A.R.d.O.d.; Carvalho, A.C.A.d.; Bechara, M.D.; Guiguer, E.L.; Goulart, R.d.A.; Vargas Sinatora, R.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and Regulation of NF-kB in Inflammatory Bowel Diseases: An Overview of In Vitro and In Vivo Effects. Metabolites 2023, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Le Loupp, A.-G.; Bach-Ngohou, K.; Bourreille, A.; Boudin, H.; Rolli-Derkinderen, M.; Denis, M.G.; Neunlist, M.; Masson, D. Activation of the prostaglandin D2 metabolic pathway in Crohn’s disease: Involvement of the enteric nervous system. BMC Gastroenterol. 2015, 15, 112. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene 2010, 29, 781–788. [Google Scholar] [CrossRef]
- Piechota-Polanczyk, A.; Fichna, J. The role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Checa, J.; Aran, J.M. Reactive oxygen species: Drivers of physiological and pathological processes. J. Inflamm. Res. 2020, 10, 1057–1073. [Google Scholar] [CrossRef] [PubMed]
- Gautam, M.; Goel, S.; Ghatule, R.; Singh, A.; Nath, G.; Goel, R. Curative effect of Terminalia chebula extract on acetic acid-induced experimental colitis: Role of antioxidants, free radicals and acute inflammatory marker. Inflammopharmacology 2013, 21, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, W.; Liang, S.; Fang, H.; Cao, J. Esculentoside A could attenuate apoptosis and inflammation in TNBS-induced ulcerative colitis via inhibiting the nuclear translocation of NF-κB. Ann. Transl. Med. 2022, 10, 771. [Google Scholar] [CrossRef]
- Raish, M.; Shahid, M.; Bin Jardan, Y.A.; Ansari, M.A.; Alkharfy, K.M.; Ahad, A.; Abdelrahman, I.A.; Ahmad, A.; Al-Jenoobi, F.I. Gastroprotective effect of sinapic acid on ethanol-induced gastric ulcers in rats: Involvement of Nrf2/HO-1 and NF-κB signaling and antiapoptotic role. Front. Pharmacol. 2021, 12, 622815. [Google Scholar] [CrossRef] [PubMed]
- Celiberto, L.S.; Graef, F.A.; Healey, G.R.; Bosman, E.S.; Jacobson, K.; Sly, L.M.; Vallance, B.A. Inflammatory bowel disease and immunonutrition: Novel therapeutic approaches through modulation of diet and the gut microbiome. Immunology 2018, 155, 36–52. [Google Scholar] [CrossRef]
- Temraz, A. Novel illudalane sesquiterpenes from Encephalartos villosus Lehm. antimicrobial activity. Nat. Prod. Res. 2016, 30, 2791–2797. [Google Scholar] [CrossRef]
- Aleisa, A.M.; Al-Rejaie, S.S.; Abuohashish, H.M.; Ola, M.S.; Parmar, M.Y.; Ahmed, M.M. Pretreatment of Gymnema sylvestre revealed the protection against acetic acid-induced ulcerative colitis in rats. BMC Complement. Altern. Med. 2014, 14, 49. [Google Scholar] [CrossRef]
- Chen, M.L.; Sundrud, M.S. Cytokine networks and T-cell subsets in inflammatory bowel diseases. Inflamm. Bowel Dis. 2016, 22, 1157–1167. [Google Scholar] [CrossRef]
- Vishwakarma, N.; Ganeshpurkar, A.; Pandey, V.; Dubey, N.; Bansal, D. Mesalazine–probiotics beads for acetic acid experimental colitis: Formulation and characterization of a promising new therapeutic strategy for ulcerative colitis. Drug Deliv. 2015, 22, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, M.J.; Elekhnawy, E.; Negm, W.A.; Mahgoub, S.; Hussein, I.A. Encephalartos villosus Lem. Displays a strong in vivo and in vitro antifungal potential against Candida glabrata clinical isolates. J. Fungi 2022, 8, 521. [Google Scholar] [CrossRef] [PubMed]
- Dejban, P.; Sahraei, M.; Chamanara, M.; Dehpour, A.; Rashidian, A. Anti-inflammatory effect of amitriptyline in a rat model of acetic acid-induced colitis: The involvement of the TLR4/NF-kB signaling pathway. Fundam. Clin. Pharmacol. 2021, 35, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Hasan Khudhair, D.; Al-Gareeb, A.I.; Al-Kuraishy, H.M.; El-Kadem, A.H.; Elekhnawy, E.; Negm, W.A.; Saber, S.; Cavalu, S.; Tirla, A.; Alotaibi, S.S. Combination of vitamin C and curcumin safeguards against methotrexate-induced acute liver injury in mice by synergistic antioxidant effects. Front. Med. 2022, 9, 866343. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhan, L.; Liao, H.; Chen, L.; Lv, X. Curcumin improves TNBS-induced colitis in rats by inhibiting IL-27 expression via the TLR4/NF-κB signaling pathway. Planta Medica 2013, 29, 102–109. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, B.; Lee, H.-S.; Bae, E.-A.; Lee, H.; Ahn, Y.-T.; Lim, K.-S.; Huh, C.-S.; Kim, D.-H. Lactobacillus suntoryeus inhibits pro-inflammatory cytokine expression and TLR-4-linked NF-κB activation in experimental colitis. Int. J. Color. Dis. 2009, 24, 231–237. [Google Scholar] [CrossRef]
- Momtaz, S.; Navabakhsh, M.; Bakouee, N.; Dehnamaki, M.; Rahimifard, M.; Baeeri, M.; Abdollahi, A.; Abdollahi, M.; Farzaei, M.H.; Abdolghaffari, A.H. Cinnamaldehyde targets TLR-4 and inflammatory mediators in acetic-acid induced ulcerative colitis model. Biologia 2021, 76, 1817–1827. [Google Scholar] [CrossRef]
- Almukainzi, M.; A El-Masry, T.; A Negm, W.; Elekhnawy, E.; Saleh, A.; E Sayed, A.; A Khattab, M.; H Abdelkader, D. Gentiopicroside PLGA nanospheres: Fabrication, in vitro characterization, antimicrobial action, and in vivo effect for enhancing wound healing in diabetic rats. Int. J. Nanomed. 2022, 22, 1203–1225. [Google Scholar] [CrossRef]
- Qu, Y.; Li, X.; Xu, F.; Zhao, S.; Wu, X.; Wang, Y.; Xie, J. Kaempferol alleviates murine experimental colitis by restoring gut microbiota and inhibiting the LPS-TLR4-NF-κB axis. Front. Immunol. 2021, 12, 679897. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.; Silva, A.M. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Chaen, Y.; Yamamoto, Y.; Suzuki, T. Naringenin promotes recovery from colonic damage through suppression of epithelial tumor necrosis factor–α production and induction of M2-type macrophages in colitic mice. Nutr. Res. 2019, 64, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Wu, X.; Guo, H.; Pan, X.; Huang, R.; Wang, G.; Liu, J. Naringin exhibited therapeutic effects against DSS-induced mice ulcerative colitis in intestinal barrier–dependent manner. Molecules 2021, 26, 6604. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.; Zhang, J.; Sun, A.; Zhang, E.; Ding, L.; Mukherjee, S.; Wei, X.; Chou, G.; Wang, Z.-T.; Mani, S. Protective effect of naringenin against experimental colitis via suppression of Toll-like receptor 4/NF-κB signalling. Br. J. Nutr. 2013, 110, 599–608. [Google Scholar] [CrossRef]
- Romier, B.; Schneider, Y.-J.; Larondelle, Y.; During, A. Dietary polyphenols can modulate the intestinal inflammatory response. Nutr. Rev. 2009, 67, 363–378. [Google Scholar] [CrossRef]
- Shi, L.; Dai, Y.; Jia, B.; Han, Y.; Guo, Y.; Xie, T.; Liu, J.; Tan, X.; Ding, P.; Li, J. The inhibitory effects of Qingchang Wenzhong granule on the interactive network of inflammation, oxidative stress, and apoptosis in rats with dextran sulfate sodium-induced colitis. J. Cell. Biochem. 2019, 120, 9979–9991. [Google Scholar] [CrossRef]
- Negm, W.A.; El-Kadem, A.H.; Elekhnawy, E.; Attallah, N.G.; Al-Hamoud, G.A.; El-Masry, T.A.; Zayed, A. Wound-Healing Potential of Rhoifolin-Rich Fraction Isolated from Sanguisorba officinalis Roots Supported by Enhancing Re-Epithelization, Angiogenesis, Anti-Inflammatory, and Antimicrobial Effects. Pharmaceuticals 2022, 15, 178. [Google Scholar] [CrossRef] [PubMed]
- Attallah, N.G.; El-Sherbeni, S.A.; El-Kadem, A.H.; Elekhnawy, E.; El-Masry, T.A.; Elmongy, E.I.; Altwaijry, N.; Negm, W.A. elucidation of the metabolite profile of Yucca gigantea and assessment of its cytotoxic, antimicrobial, and anti-inflammatory activities. Molecules 2022, 27, 1329. [Google Scholar] [CrossRef]
- Ramadan, A.; Afifi, N.; Yassin, N.Z.; Abdel-Rahman, R.F.; Abd El-Rahman, S.S.; Fayed, H.M. Mesalazine, an osteopontin inhibitor: The potential prophylactic and remedial roles in induced liver fibrosis in rats. Chem.-Biol. Interact. 2018, 289, 109–118. [Google Scholar] [CrossRef]
- Attallah, N.G.; El-Kadem, A.H.; Negm, W.A.; Elekhnawy, E.; El-Masry, T.A.; Elmongy, E.I.; Altwaijry, N.; Alanazi, A.S.; Al-Hamoud, G.A.; Ragab, A.E. Promising Antiviral Activity of Agrimonia pilosa Phytochemicals against Severe Acute Respiratory Syndrome Coronavirus 2 Supported with In Vivo Mice Study. Pharmaceuticals 2021, 14, 1313. [Google Scholar] [CrossRef]
- Arab, H.H.; Al-Shorbagy, M.Y.; Abdallah, D.M.; Nassar, N.N. Telmisartan attenuates colon inflammation, oxidative perturbations and apoptosis in a rat model of experimental inflammatory bowel disease. PLoS ONE 2014, 9, e97193. [Google Scholar] [CrossRef]
- Hussein, S.Z.; Mohd Yusoff, K.; Makpol, S.; Mohd Yusof, Y.A. Gelam honey inhibits the production of proinflammatory, mediators NO, PGE2, TNF-α, and IL-6 in carrageenan-induced acute paw edema in rats. Evid. Based Complement. Altern. Med. 2012, 2012, 109636. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Grover, J.; Jachak, S.M. Coumarins as privileged scaffold for anti-inflammatory drug development. RSC Adv. 2015, 5, 38892–38905. [Google Scholar] [CrossRef]
- Raish, M.; Ahmad, A.; Jardan, Y.A.B.; Shahid, M.; Alkharfy, K.M.; Ahad, A.; Ansari, M.A.; Abdelrahman, I.A.; Al-Jenoobi, F.I. Sinapic acid ameliorates cardiac dysfunction and cardiomyopathy by modulating NF-κB and Nrf2/HO-1 signaling pathways in streptozocin induced diabetic rats. Biomed. Pharmacother. 2022, 145, 112412. [Google Scholar] [CrossRef]
- Ali, A.A.; Abd Al Haleem, E.N.; Khaleel, S.A.-H.; Sallam, A.S. Protective effect of cardamonin against acetic acid-induced ulcerative colitis in rats. Pharmacol. Rep. 2017, 69, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Luissint, A.-C.; Parkos, C.A.; Nusrat, A. Inflammation and the intestinal barrier: Leukocyte–epithelial cell interactions, cell junction remodeling, and mucosal repair. Gastroenterology 2016, 151, 616–632. [Google Scholar] [CrossRef] [PubMed]
- Almero, J.M. Influence of Toll-like receptor 2 and interleukin 10 on the intestinal epithelial barrier and their roles in inflammatory bowel disease. Inmunología 2011, 30, 8–16. [Google Scholar] [CrossRef]
- Albert-Garay, J.S.; Riesgo-Escovar, J.R.; Salceda, R. High glucose concentrations induce oxidative stress by inhibiting Nrf2 expression in rat Müller retinal cells in vitro. Sci. Rep. 2022, 12, 1261. [Google Scholar] [CrossRef]
- Wu, M.-L.; Ho, Y.-C.; Lin, C.-Y.; Yet, S.-F. Heme oxygenase-1 in inflammation and cardiovascular disease. Am. J. Cardiovasc. Dis. 2011, 1, 150. [Google Scholar] [PubMed]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.A.; Zhang, M.; Yin, F. Heme oxygenase-1, oxidation, inflammation, and atherosclerosis. Front. Pharmacol. 2012, 3, 119. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-C.; Yeh, C.-H.; Yang, M.-L.; Kuan, Y.-H. Luteolin suppresses inflammatory mediator expression by blocking the Akt/NFκB pathway in acute lung injury induced by lipopolysaccharide in mice. Evid. Based Complement. Altern. Med. 2012, 2012, 383608. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.-H.; Weng, J.-R.; Tsao, L.-T.; Yen, M.-H.; Wang, J.-P.; Lin, C.-N. Anti-inflammatory flavonoids and pterocarpanoid from Crotalaria pallida and C. assamica. Bioorg. Med. Chem. Lett. 2004, 14, 1011–1014. [Google Scholar] [CrossRef]
- Buchmann, D.; Schultze, N.; Borchardt, J.; Böttcher, I.; Schaufler, K.; Guenther, S. Synergistic antimicrobial activities of epigallocatechin gallate, myricetin, daidzein, gallic acid, epicatechin, 3-hydroxy-6-methoxyflavone and genistein combined with antibiotics against ESKAPE pathogens. J. Appl. Microbiol. 2022, 132, 949–963. [Google Scholar] [CrossRef]
- El-Akabawy, G.; El-Sherif, N.M. Zeaxanthin exerts protective effects on acetic acid-induced colitis in rats via modulation of pro-inflammatory cytokines and oxidative stress. Biomed. Pharmacother. 2019, 111, 841–851. [Google Scholar] [CrossRef]
- Attallah, N.G.; Kabbash, A.; Negm, W.A.; Elekhnawy, E.; Binsuwaidan, R.; Al-Fakhrany, O.M.; Shaldam, M.A.; Moglad, E.; Tarek, M.; Samir, N. Protective Potential of Saussurea costus (Falc.) Lipsch. Roots against Cyclophosphamide-Induced Pulmonary Injury in Rats and Its In Vitro Antiviral Effect. Pharmaceuticals 2023, 16, 318. [Google Scholar] [CrossRef]
- Bezerra, G.B.; de Souza, L.d.M.; Dos Santos, A.S.; de Almeida, G.K.M.; Souza, M.T.S.; Santos, S.L.; Camargo, E.A.; dos Santos Lima, B.; de Souza Araújo, A.A.; Cardoso, J.C. Hydroalcoholic extract of Brazilian red propolis exerts protective effects on acetic acid-induced ulcerative colitis in a rodent model. Biomed. Pharmacother. 2017, 85, 687–696. [Google Scholar] [CrossRef]
- Li, J.; Chen, C.; Cao, X.-N.; Wang, G.-H.; Hu, J.-B.; Wang, J. Efficacy of topical versus oral 5-aminosalicylate for treatment of 2, 4, 6-trinitrobenzene sulfonic acid-induced ulcerative colitis in rats. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 59–65. [Google Scholar] [CrossRef]
- Abdin, A.A. Targeting sphingosine kinase 1 (SphK1) and apoptosis by colon-specific delivery formula of resveratrol in treatment of experimental ulcerative colitis in rats. Eur. J. Pharmacol. 2013, 718, 145–153. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Jeon, H.; Bae, C.H.; Lee, Y.; Kim, H.; Kim, S. Rumex japonicus Houtt. alleviates dextran sulfate sodium-induced colitis by protecting tight junctions in mice. Integr. Med. Res. 2020, 9, 100398. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, B.; El-Masry, T.A.; Elekhnawy, E.; El-Kadem, A.H.; Saleh, A.; Negm, W.A.; Abdelkader, D.H. Aqueous core epigallocatechin gallate PLGA nanocapsules: Characterization, antibacterial activity against uropathogens, and in vivo reno-protective effect in cisplatin induced nephrotoxicity. Drug Deliv. 2022, 29, 1848–1862. [Google Scholar] [CrossRef] [PubMed]
- Elekhnawy, E.; Negm, W.A. The potential application of probiotics for the prevention and treatment of COVID-19. Egypt. J. Med. Hum. Genet. 2022, 23, 36. [Google Scholar] [CrossRef]
- Sivakumar, M.; Yoithapprabhunath, T.R.; Nirmal, R.M.; Veeravarmal, V.; Dineshshankar, J.; Amsaveni, R. Immunohistochemical analysis of Nuclear Factor-kappa B (NF-κB) between follicular and plexiform ameloblastomas: A pilot study. J. Oral Maxillofac. Pathol. JOMFP 2020, 24, 466. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).