Abstract
Background: The balance between antioxidants and pro-oxidants plays a significant role in the context of oxidative stress, influenced by both physiological and non-physiological factors. Objectives: In this study, 18 prescribed antibiotics (including doxycycline hydrochloride, tigecycline, rifampicin, tebipenem, cefuroxime, cefixime, potassium clavulanate, colistin, ampicillin, amoxicillin, amikacin, nalidixic acid, azithromycin, pipemidic acid trihydrate, pivmecillinam, aztreonam, fosfomycin sodium, and ciprofloxacin) were subjected to simultaneous determination of antioxidant and pro-oxidant potential to assess if pro-oxidant activity is a dominant co-mechanism of antibacterial activity or if any antibiotic exhibits a balanced effect. Methods: This study presents a recently developed approach for the simultaneous assessment of antioxidant and pro-oxidant potential on a single microplate in situ, applied to prescribed antibiotics. Results: Ten antibiotics from eighteen showed lower antioxidant or pro-oxidant potential, while five exhibited only mild potential with DPPH50 values over 0.5 mM. The pro-oxidant antioxidant balance index (PABI) was also calculated to determine whether antioxidant or pro-oxidant activity was dominant for each antibiotic. Surprisingly, three antibiotics—doxycycline hydrochloride, tigecycline, and rifampicin—showed significant measures of both antioxidant and pro-oxidant activities. Especially notable was tebipenem, a broad-spectrum, orally administered carbapenem, showed a positive PABI index ratio, indicating a dominant antioxidant over pro-oxidant effect. Conclusions: These findings could be significant for both therapy, where the antibacterial effect is enhanced by radical scavenging activity, and biotechnology, where substantial pro-oxidant activity might limit microbial viability in cultures and consequently affect yield.
1. Introduction
The anatomical therapeutic chemical (ATC) classification system currently records more than 200 antibiotics approved for clinical or veterinary use [1]. This relatively large number and diversity arise from the need for antibiotic therapy for different bacterial infections. The situation is further complicated by increasing bacterial resistance to available antibiotics [2,3]. Such infections were significant during the COVID-19 pandemic, where cytokine inflammatory storms [4] and secondary bacterial infections [5] significantly contributed to patient mortality. Managing these superinfections often involves combination antibiotic therapy [6]. The rise in bacterial resistance poses a significant threat, potentially leading to a shortage of effective antibiotics for future pandemics.
Antibiotics are widely recognized for their role in inhibiting bacterial growth by targeting vital cellular processes such as protein synthesis, nucleic acid replication, and cell wall biosynthesis [7]. Antibiotics can be classified as either bacteriostatic, halting bacterial growth and reproduction, or bactericidal, inducing oxidative stress within bacterial cells and leading to cell death [8]. Recent research has highlighted additional roles antibiotics may play in influencing the redox environment within bacterial and host cells. Specifically, antibiotics may exert dual pro-oxidant and antioxidant effects, contributing to their therapeutic efficacy or adverse effects. Pro-oxidant properties, often associated with oxidative stress generation, contribute to the bactericidal activity by inducing reactive oxygen species (ROS), reactive nitrogen species (RNS), or reactive carbonyl species, which damage bacterial DNA, proteins, and lipids, ultimately leading to cell death [9,10,11,12,13]. Several mechanisms have been described for ROS overproduction, such as the Krebs cycle and electron transport chain activation, leading to the formation of peroxide radicals [14]. Heleen van Acker and Tom Coenye proposed a mechanism for bacterial cell death due to hydroxyl radical production, which can be promoted by Fenton reactions involving transition metals like iron and copper [10]. Oxidative burst is a physiological mechanism by which macrophages eliminate bacterial cells [15]. The consequences of oxidative stress in bacterial cells include DNA breaks, lipid peroxidation, protein carbonylation, and other irreversible changes. ROS-induced cell death is primarily linked to the guanine nucleotide pool’s oxidative damage (the formation of 8-oxo-deoxyguanosine lesions), leading to oxidized nucleotides’ incorporation into RNA and DNA [12]. This ROS-mediated mechanism is often utilized by bactericidal antibiotics such as beta-lactams, fluoroquinolones, and aminoglycosides. Conversely, some antibiotics like minocycline, azithromycin, and doxycycline demonstrate antioxidant properties, which can mitigate unwanted oxidative damage in host tissues [16]. This balance between pro-oxidant and antioxidant effects may influence not only bacterial survival but also tissue recovery during infection. The dual properties of antibiotics—pro-oxidant and antioxidant—are of growing interest, as they may play a crucial role in therapeutic outcomes, resistance mechanisms, and even in biotechnological processes where antibiotics are used in microbial cultures. Antibiotics are primarily produced through the fermentation of soil microorganisms, particularly actinomycetes, micromycetes, and other groups of soil microbes. The predominant pro-oxidant effect of antibiotics can significantly negatively impact their biotechnological production. Penicillins, beta-lactam antibiotics, are originally derived from Penicillium fungi, such as P. chrysogenum and P. rubens, and are typically synthesized fermentatively [17]. Semisynthetic penicillins are derived from 6-aminopenicillinic acid (6-APA), which contains the beta-lactam core of Penicillin G but with modified side chains [18,19]. Broad-spectrum antibiotics (ampicillin and amoxicillin) are effective against a wide range of Gram-negative bacteria, including Escherichia coli and Salmonella typhi, and are further divided into carboxypenicillins and ureidopenicillins. Penicillins, for example, are beta-lactam antibiotics derived from Penicillium molds and are typically synthesized fermentatively. Tetracyclines and macrolides, such as erythromycin, are also produced fermentatively. The complex media used for these cultures contain nutrients and elements that can facilitate pro-oxidant effects through Fenton-like reactions. Similarly, cephalosporins are derived from 7-aminocephalosporanic acid (7-ACA), a core structure for synthesizing beta-lactam antibiotics originally isolated from Cephalosporium acremonium [20,21]. Fermentative production of tetracyclines served as a template for semisynthetic derivatives [22]. Chlortetracycline was first isolated in 1945 [23], followed by the structural identification of oxytetracycline from Streptomyces rimosus [24]. The first macrolide, erythromycin, was isolated in 1952 from Saccharopolyspora erythraea and is listed among the World Health Organization’s essential medicines. Antibiotics’ toxic effects on eukaryotic cells and their metabolic burden on the liver are significant [25]. Culture media are complex, containing essential nutrients and macro- and microelements. Numerous studies highlight the significant pro-oxidant effects of compounds with heteroatoms, particularly those with multiple bonds. These structural conformations can form coordination complexes with transition metals, contacting with hydrogen peroxide participating in Fenton and Fenton-like reactions to produce ROS and RNS [26,27,28].
Currently, various methods exist for measuring antioxidant and pro-oxidant potential, although there is no universally accepted definition or standard protocol for either. Common colorimetric assays for antioxidant activity determination include DPPH (2,2-diphenyl-1-picrylhydrazyl), TRAP (total radical-trapping antioxidant parameter), ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)), TAR (total antioxidant response), ORAC (oxygen radical absorbance capacity), CUPRAC (cupric reducing antioxidant capacity), and FRAP (ferric reducing antioxidant power). Frequently used pro-oxidant activity measurement methods involve FRAP, Fenton reaction-based assays, TBARS (thiobarbituric acid reactive substances), and metal-catalyzed oxidation assays. The FRAP assay is particularly versatile, as it can be used to quantify both antioxidant and pro-oxidant activities by measuring substances that react with divalent or trivalent iron. In the context of pro-oxidant activity, the FRAP assay assesses the induction of lipid peroxidation. Another widely used method, the DPPH assay, measures antiradical activity but is limited to water-soluble antioxidants. These methods have inherent limitations and conventions, such as varying levels of specificity, sensitivity, and relevance, depending on the biological system and antioxidant or pro-oxidant properties being studied [29].
Recent advancements have allowed for the simultaneous determination of antioxidant and pro-oxidant potential using a single microplate to evaluate the pro-oxidant antioxidant balance index (PABI) [29]. Addressing the therapeutic aspects and biotechnological production of antibiotics is essential. Understanding the balance between antioxidant and pro-oxidant properties can enhance therapeutic efficacy by potentially improving infection outcomes and aiding tissue healing. Additionally, insights into pro-oxidant effects are crucial for optimizing biotechnological production processes, ensuring microbial viability and maximizing yield. Furthermore, this study could contribute to solving the problem of bacterial resistance by identifying antibiotic properties that enhance antibacterial efficacy through oxidative mechanisms, potentially leading to the development of more effective treatment strategies. Given the significance of oxidative stress in bacterial and host cell interactions, this study aims to further explore the antioxidant and pro-oxidant potential of 18 commonly prescribed antibiotics in relation to their physiological effects and the limitations of the biotechnological production of their precursors. Using the DPPH and FRAP assays, we investigate whether specific antibiotics favor pro-oxidant or antioxidant mechanisms under different conditions.
2. Results
We evaluated 18 selected antibiotics, each representing different mechanisms of action: doxycycline hydrochloride and tigecycline (tetracyclines) [30,31]; amikacin (aminoglycoside) [32] and azithromycin (macrolide) [33], which inhibit protein synthesis (PSI); beta-lactams, including penicillins (ampicillin, amoxicillin, and pivmecillinam) [34,35,36]; carbapenem (tebipenem) [37]; cephalosporins (cefuroxime and cefixime) [38,39]; monobactam (aztreonam) [40] and a beta-lactamase inhibitor (potassium clavulanate) [41], along with colistin (polymyxin E) [42] and fosfomycin sodium [43], which target the bacterial cell membrane and peptidoglycan in the cell wall (BCED); and rifampicin [44], nalidixic acid [45], pipemidic acid trihydrate [46], and ciprofloxacin [47], which act on bacterial DNA replication and transcription processes (NAB). The structures and CAS numbers of evaluated antibiotics presents Table S1 in supplementary file.
Antioxidant potential was determined using the DPPH method, and pro-oxidant potential was determined using the FRAP method, with results expressed as DPPH50 and FRAP50 values, respectively. The obtained results, along with antibiotic classification and antibacterial mechanisms, are presented in Table 1, ordered by descending antioxidant potential.

Table 1.
Antioxidant and pro-oxidant potential of selected antibiotics with their classification and antibacterial mechanisms.
Only six antibiotics—doxycycline hydrochloride, tigecycline, rifampicin, tebipenem, cefuroxime, and cefixime—demonstrated detectable antioxidant potential. Potassium clavulanate exhibited nonsignificant pro-oxidant effect. Doxycycline hydrochloride (PABI index 0.27) and rifampicin (PABI index 0.26) showed significant pro-oxidant potential compared to their antioxidant properties, approximately fourfold, suggesting a link to their antibacterial mechanisms. In contrast, tebipenem, a carboxypenicillin, displayed an unexpectedly high antioxidant potential (PABI index = 6.12), indicating a strong radical scavenging ability that may contribute to the healing process in inflamed tissues near infection sites. All other antibiotics did not exhibit any detectable antioxidant or pro-oxidant potential within the tested concentration range up to 4096 μM, indicating that their antibacterial effects are likely based on mechanisms other than redox modulation.
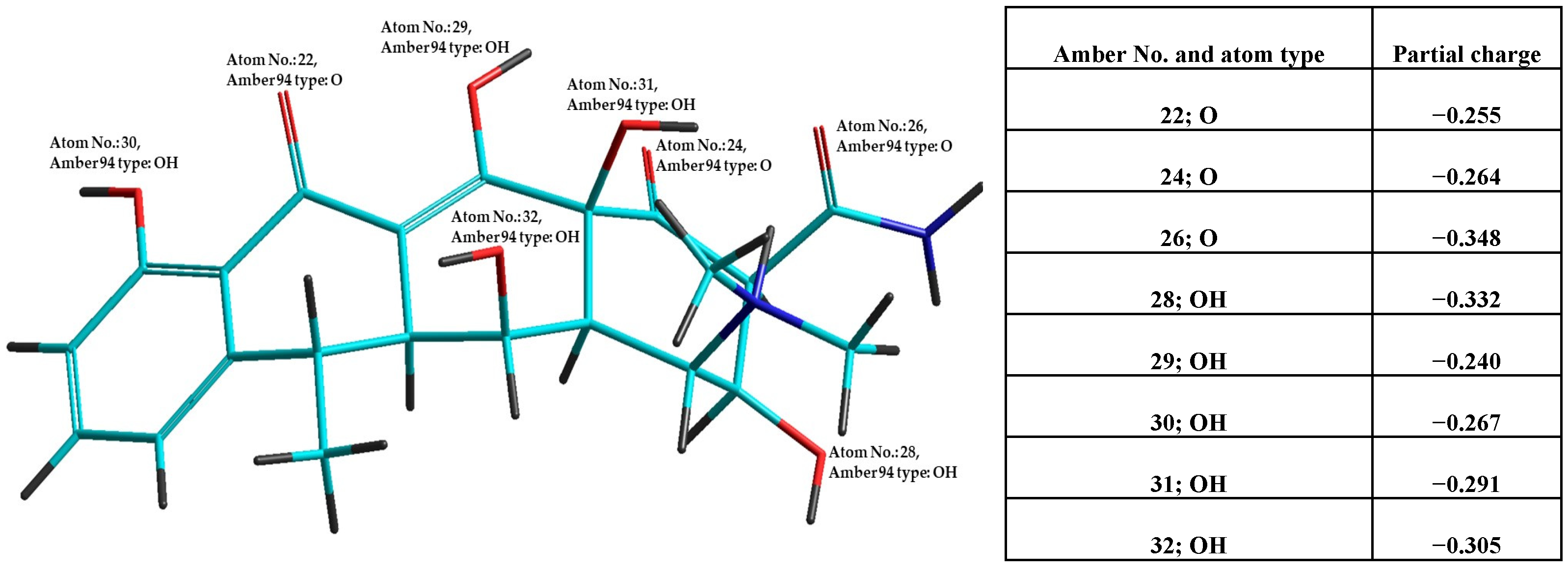
To elucidate the observed antioxidant and pro-oxidant potential, in silico calculations were conducted using the Molinspiration program/Molinspiration Cheminformatics 2024/, (Table 2). For all 18 selected antibiotics, parameters based on Lipinski’s rule of five were calculated, including the number of violations of these criteria and the bioactivity predictions across six different mechanisms. These in silico predictions are complemented by results from calculations using Hyperchem 7.5. For demonstration, Figure 1 presents the optimized structure of the antibiotic doxycycline hydrochloride after combined molecular mechanics (semiempirical quantum mechanics /MM-QM/ calculation, with oxygen atoms labeled and partial charges indicated).
Table 2.
The calculations conducted by Molinspiration/Molinspiration Cheminformatics 2024/ and Hyperchem 7.5 software in silico.

Figure 1.
Structure of the antibiotic doxycycline quaternary cation after MM-QM optimization. Oxygen atoms are labeled with their atom number and type, and partial charges are provided in the accompanying table.
Bioactivity predictions, expressed as dimensionless scores, are significant for positive values or those nearing the maximum of 0.5. While the primary mechanisms of action for antibiotics are well-established, such as the inhibition of penicillin-binding proteins (PBPs) by beta-lactams (penicillins, cephalosporins, carbapenems, and monobactams), alternative mechanisms also exist. Molinspiration’s predictions indicate various potential mechanisms. Six antibiotics, including ampicillin, amoxicillin, amikacin, pivmecillinam, and aztreonam, may act as GPCR ligands or inhibitors of proteases and enzymes. Cefuroxime and cefixime (cephalosporins) demonstrate potential as protease and enzyme inhibitors, as do tetracycline, doxycycline hydrochloride, and potassium clavulanate. Ciprofloxacin (a fluoroquinolone) also shows potential as a GPCR ligand and protease inhibitor.
Using Hyperchem software, we analyzed the oxygen atoms in antibiotic molecules. As heteroatoms, oxygen atoms are significant hydrogen bond acceptors and, as hydroxyl groups, also hydrogen bond donors. Hydroxyl groups on sp2 hybridized carbon atoms can participate in hydrogen atom transfer (HAT) mechanisms, with hydrogen bond formation influenced by the OH group’s hybridization and the partial charge on oxygen, affecting bond polarity. The data in Table 2 show that the number and partial charge of oxygen atoms are unique structural characteristics for each antibiotic. The antibiotics in both tables are ranked by descending antioxidant activity, correlated with the number of OH groups on sp2 hybridized carbon atoms (aromatic OH groups). These oxygen atoms exhibit lower partial charges, as reflected in the last two columns of the table. This is due to the +M effect of conjugating OH groups with aromatic rings, enhancing conditions for hydrogen bond donation via the HAT mechanism. The primary impact of antibiotics may be augmented by pro-oxidant effects and the formation of ROS/RNS in bacterial cells, as indicated by FRAP50 values. This is particularly evident for tetracyclines and Rifamycin, which exhibit antioxidant potential. Among tetracyclines, tigecycline shows a balanced antioxidant and pro-oxidant effect. Tebipenem, a carbapenem, demonstrates predominantly antioxidant potential, which, although not high, may contribute to the healing process of post-infection clearance.
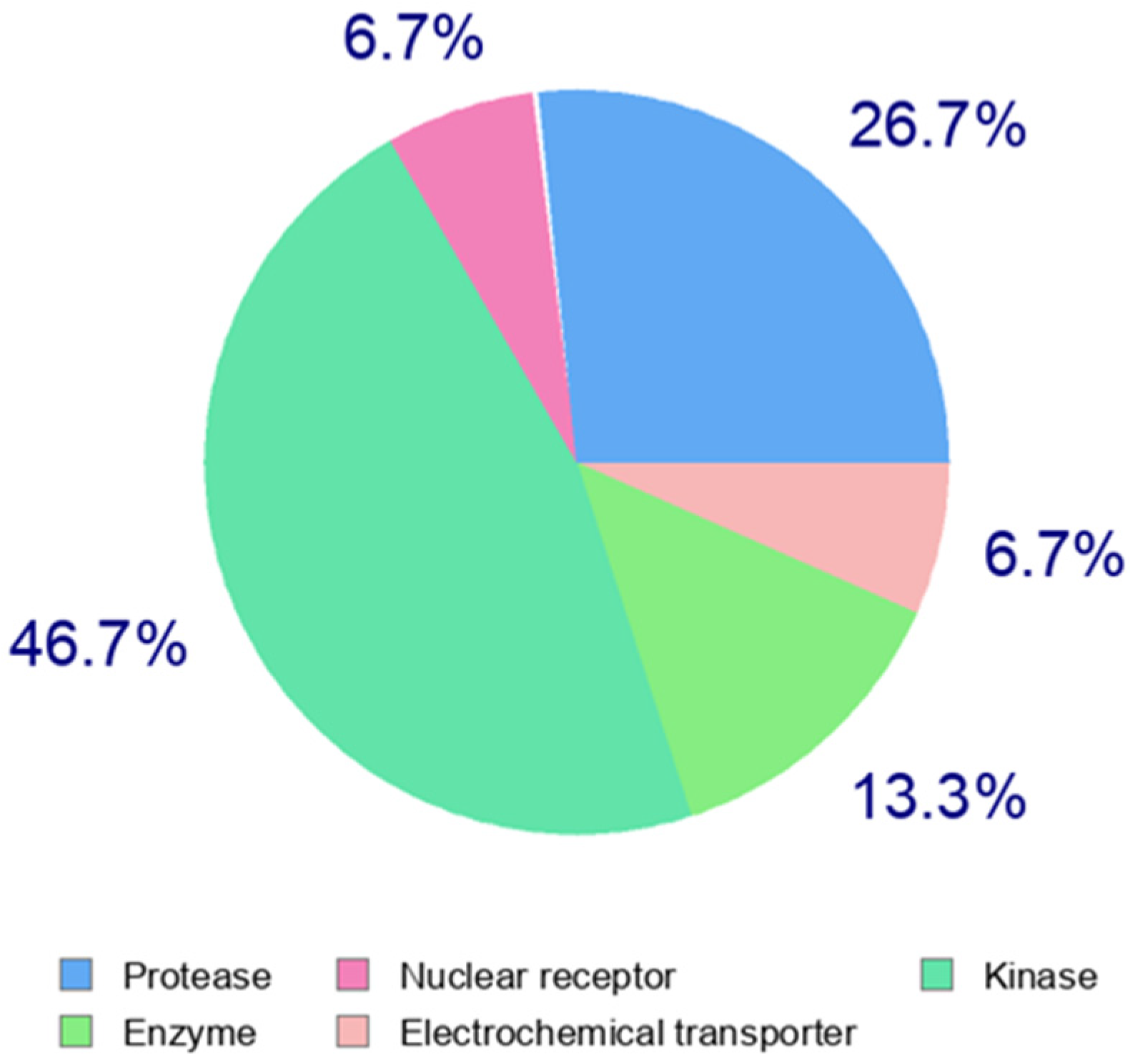
The effects of antibiotics may not be strictly monofactorial. The complete impact on the biochemical pathways of prokaryotic and eukaryotic cells is likely not fully understood. The diversity and variability of possible antibiotic effects, even among structurally similar compounds within the same group, are shown in Table 3. This table outlines the estimated potential targets (target categories) in human cells and systems based on the structure of the antibiotic. The analysis was processed using the Swiss target prediction web application, listing the 2–3 target categories with the highest probability of interaction, and specifying the top-predicted target in the last column. For illustration, Figure 2 presents a pie chart for the tetracycline doxycycline hydrochloride, demonstrating the high structural variability of antibiotics and the corresponding range of potential targets.
Table 3.
Predicted human targets for selected antibiotics based on structural analysis.

Figure 2.
Target prediction analysis of possible human targets for the tetracycline doxycycline hydrochloride.
The differences are also evident when comparing pairs of similar antibiotics, such as doxycycline hydrochloride vs. tigecycline or ampicillin vs. amoxicillin. Generally, antibiotics target various biochemical pathways, primarily inhibiting kinases, lyases, oxidoreductases, phosphatases, and enzymes in general. They also engage G protein-coupled receptors (families A and E) and nuclear receptors. To a lesser extent, they affect electrochemical transporters, cytosolic proteins, and other targets. This multifactorial impact can be approximated to predict potential targets in prokaryotic cells as well. Additionally, this target prediction analysis is valuable for estimating the potential toxicity of antibiotics in the human body.
3. Discussion
Our study investigated the antioxidant and pro-oxidant properties of 18 clinically applied antibiotics, aiming to understand their dual roles in therapeutic efficacy and potential biotechnological implications. We tested 2 out of 18 tetracycline antibiotics, 2 out of 50 cephalosporin antibiotics, and 1 out of 23 macrolide antibiotics. The study revealed that several clinically relevant antibiotics exhibit both antioxidant and pro-oxidant potential, which may contribute to their therapeutic mechanisms and influence host responses during bacterial infections. The balance between these properties is crucial in determining whether an antibiotic primarily acts through oxidative stress induction or provides protective antioxidant effects to host tissues. These results highlight the complexity of antibiotic actions beyond their primary antibacterial mechanisms.
Our comprehensive in silico study (unpublished data) involving 214 clinically approved antibiotics confirmed a significant dominance of hydrogen bond acceptors over hydrogen bond donors. This finding suggests a prevailing pro-oxidant effect over the antioxidant effect. The data for a typical antibiotic show compounds of borderline size with marked polarity, as indicated by the following parameters: molecular weight (MW) = 464.93 g/mol, milogP = −0.217, number of atoms = 32, number of hydrogen bond acceptors = 10, number of hydrogen bond donors = 4, number of rotatable bonds = 6, and positive bioactivity predictions only in two cases: protease inhibitor (0.112) and general enzyme inhibitor (0.017).
Clinically approved antibiotics do not necessarily comply with all Lipinski criteria. For instance, tigecycline, rifamycin, colistin, and amikacin violate three out of five Lipinski criteria, often due to their molecular size, which affects molecular weight and the number of hydrogen bond donors and acceptors. Despite these violations, these antibiotics are approved, indicating that not all clinically approved drugs must strictly adhere to these criteria.
The observed antioxidant and pro-oxidant potential provide insights into the potential mechanisms through which antibiotics exert their therapeutic effects. The generation of ROS and subsequent oxidative damage are essential components of the bactericidal activity of antibiotics. Antibiotics that induce oxidative stress have been shown to cause significant physiological alterations in bacterial cells, contributing to their lethality [48,49]. Among the antibiotics tested, doxycycline hydrochloride and rifampicin exhibited significant pro-oxidant properties, suggesting that their antibacterial mechanisms likely involve oxidative stress induction. Both antibiotics displayed high ROS generation, likely contributing to bacterial DNA damage and cellular death. Kohanski et al. [50] demonstrated that bactericidal antibiotics, such as beta-lactams, quinolones, and aminoglycosides, stimulate the production of harmful hydroxyl radicals through an oxidative damage pathway. Interestingly, the antioxidant potential of these antibiotics was relatively low, indicating that their primary mode of action is through ROS-mediated mechanisms.
On the other hand, tebipenem showed a distinct antioxidant profile, with significantly higher antioxidant potential compared to its pro-oxidant effect. This finding aligns with previous reports of tebipenem’s role in reducing oxidative stress in host tissues, which may facilitate tissue recovery post-infection. The strong radical scavenging ability of Tebipenem could play a supportive role in clinical settings, particularly in inflammatory conditions where excessive ROS could exacerbate tissue damage. However, it is important to consider the risk of antioxidant stress, where an overabundance of antioxidants may suppress essential ROS signaling and disrupt redox balance. This phenomenon may also create an environment that is harmful to micro-organisms, as seen in studies where antiviral drugs with strong antioxidant properties effectively counteract viral oxidative stress. While a pro-oxidant effect is desirable for its bactericidal action, antioxidant properties, like those of tebipenem and tetracyclines, may not diminish antibacterial efficacy. Instead, they may help in post-infection tissue repair, but caution should be taken regarding the balance of redox regulation. This unique antioxidant property contrasts with the ROS generation typically observed in other beta-lactams and highlights the complexity of redox balance in antibiotic therapy. Furthermore, the data from the DPPH and FRAP assays revealed that the majority of antibiotics tested did not exhibit significant redox-modulating effects at the concentrations studied. This suggests that the primary mechanisms of action for most antibiotics are likely unrelated to redox balance and are instead driven by direct inhibition of bacterial cellular processes. However, understanding the redox properties of antibiotics like tebipenem and rifampicin could inform future therapeutic strategies, particularly in cases where managing oxidative stress is crucial to patient recovery.
This mechanism, in which oxidative stress and resulting genomic instability play a central role in the effectiveness of antibacterial agents, highlights the complexity of antibiotic action [51]. The published pro-oxidant effects of nitrofurantoin [52] and polymyxin B [53], although not included in our test collection, further support this mechanism. Our study did not observe significant oxidative or pro-oxidant effects for colistin (polymyxin E). Hoeksema et al. [54] confirmed that ROS production is an additional mechanism of action for beta-lactam antibiotics, quinolones, and aminoglycosides, with interesting implications for de novo-acquired resistance. Under aerobic conditions, beta-lactam antibiotics at higher concentrations trigger significant ROS production in Enterococcus faecalis, whereas this is not observed under anaerobic conditions, suggesting an interaction mechanism with the respiratory chain [55].
Besides antibiotics, ROS generation is typical for metallic nanoparticles as green synthesis products [56]. They generate high amounts of ROS which cause damage to bacterial cells. Nanoparticles made of silver, silver oxide, titanium dioxide, silicon, copper oxide, zinc oxide, gold, calcium oxide, and magnesium oxide have been studied and found to be effective against both Gram-positive and Gram-negative bacteria [57].
It is essential to distinguish between direct ROS production, observed in the presence of transition metals by the antibiotics identified in our study, and indirect ROS production as a terminal state of blocked key metabolic pathways in bacterial cells. In this context, attention should be given to antibiotics that inhibit oxidoreductases or electrochemical transporters, as indicated in Table 3.
The findings of this study have significant implications for the biotechnological production of antibiotics. Understanding the balance between antioxidant and pro-oxidant properties can aid in optimizing fermentation processes and improving the yield and stability of antibiotic production. Antibiotics that exhibit high pro-oxidant activity may pose challenges during production, as oxidative stress can negatively affect the microbial strains used in fermentation. Conversely, antibiotics with notable antioxidant properties, such as tebipenem, may enhance microbial resilience and improve overall production efficiency. Future biotechnological strategies could involve modifying culture conditions to mitigate oxidative stress or engineering microbial strains with enhanced antioxidant defenses to boost production yields [58]. By considering the redox properties of antibiotics, biotechnological processes can be tailored to maximize productivity while ensuring the quality and efficacy of the final product.
Limitations
The limitations of the DPPH and FRAP assays have been well documented in the literature. Both methods utilize synthetic reagents that do not fully mimic physiological conditions, which restricts their ability to represent in vivo antioxidant or pro-oxidant dynamics accurately. The DPPH assay, which measures the ability of compounds to scavenge the stable DPPH radical, primarily reflects the interaction with a model radical that does not exist in biological systems. This reduces the physiological relevance of the assay when interpreting how antioxidants behave in complex biological environments.
4. Materials and Methods
4.1. Materials and Chemicals
Chemicals and solvents for assays, such as 2,2-diphenyl-1-picrylhydrazyl radical (DPPH●), 2,2-diphenyl-1-picrylhydrazine (DPPH), 2,4,6-tris(2-pyridyl)-s-triazine (TPTZ), FeCl2·4H2O, FeCl3·6H2O, TROLOX, sodium acetate, ethanol, and acetic acid were purchased from Merck/Sigma (St. Louis, MO, USA). Similarly, the following antibiotics were purchased as standards from Merck/Sigma (USA): doxycycline hydrochloride, tigecycline, rifampicin, tebipenem, cefuroxime, cefixime, potassium clavulanate, colistin, ampicillin, amoxicillin, amikacin, nalidixic acid, azithromycin, pipemidic acid trihydrate, pivmecillinam, aztreonam, fosfomycin sodium, and ciprofloxacin. Standard 96-microwell plates (F-type, SPL Life Sciences, Pocheon-si, Republic of Korea) were used.
4.2. Measurement of Antioxidant and Pro-Oxidant Properties Using the Modified DPPH/FRAP Method
The modified DPPH/FRAP method was used to simultaneously measure the antioxidant and pro-oxidant properties of compounds, as described by Maliar et al. [29]. The DPPH method was used to measure antioxidant activity, and the FRAP method was used to measure both antioxidant and pro-oxidant activities. Modified microplate assays ensured equal concentrations of key reagents (DPPH, TPTZ, FeCl3) at 0.4 mM. Conversion standards were set with DPPH and FeCl2·4H2O for 100% conversion and DPPH● and FeCl3·6H2O for 0% conversion.
The antibiotic solutions were diluted to achieve final concentrations ranging from 4096 μM to 8 μM. The microplate preparation involved applying conversion standards and an FeCl3 solution. Assays started with adding 0.4 mM DPPH and FRAP reagents, achieving a final concentration of 0.3 mM. The microplate was incubated for 10 min for DPPH and 1 h for FRAP, followed by measurements at 520 nm and 630 nm for DPPH and FRAP, respectively.
DPPH50 and FRAP50 values were calculated, with the pro-oxidant antioxidant balance index (PABI), determined as FRAP50/DPPH50 [28]. Each experiment was repeated three times with eight replicates, with results presented as mean ± SD. Statistical significance was determined using the Spearman method (* p < 0.1).
4.3. Molinspiration Calculation
The Molinspiration/Molinspiration Cheminformatics 2024/ calculations include Lipinski’s “drug-likeness” parameters (rule of five), based on the following:
- Octanol/water partition coefficient (LogP): calculated as a sum of fragment-based contributions and correction factors;
- Topological polar surface area (TPSA): calculated based on methodology of (https://molinspiration.com/ accessed on 17 September 2024) with summing fragment contributions fitted to the 3D volume of a training set of about 12,000, mostly drug-like, molecules. These geometries were optimized using the semi-empirical AM1 method;
- Rule of five: Most “drug-like” molecules have logP ≤ 5, molecular weight ≤ 500, ≤10 hydrogen bond acceptors, and ≤5 hydrogen bond donors;
- Number of rotatable bonds (nrotb): measures molecular flexibility and is a good descriptor of oral bioavailability and defined as any single non-ring bond bound to a non-terminal heavy atom, excluding amide C-N bonds due to their high rotational energy barrier.
The number of violations of Lipinski’s “drug-likeness” parameters, specifying which parameters are outside the criteria, was recorded. In the second step, the bioactivity score predictions were calculated using the Molinspiration virtual screening toolkit (miscreen) for GPCR ligands (GPCRL), ion channel modulators (ICHM), kinase inhibitors (KI), nuclear receptor ligands (NRL), protease inhibitors (PI), and other enzyme inhibitors (EI). Finally, the average bioactivity scores across these six different mechanisms were calculated, as the exact mechanism of action in bacterial cells may not be fully known or described.
4.4. Swiss Target Prediction Calculation
SwissTargetPrediction (http://swisstargetprediction.ch/index.php, accessed on 17 September 2024) predicts bioactive molecule targets using 2D and 3D similarity measures with known ligands. Predictions are available for five organisms, with homology mapping for close paralogs and orthologs. The prediction utilizes a library of 370,000 known actives on over 3000 proteins across 3 species, sourced from the ChEMBL database. Similarity thresholds are 0.85 for ES5D (shape) and 0.65 for FP2 (2D). Below these thresholds, the known actives are not considered similar enough to be displayed. More details can be found at SwissTargetPrediction [29].
In this study, we applied the prediction of human targets. The structures of antibiotics were drawn by specifying SMILES in the designated text box.
4.5. Optimization of Antibiotics Structure in Vacuum for Parameters Calculation
The partial charges of key atoms (oxygens) in all evaluated antibiotic structures were calculated by Hyperchem 8.52 using a combination of molecular mechanics and semiempirical methods, shortly MM-QM optimization, (forcefield AMBER and AM1) under defined terminal conditions in Hyperchem 7.5 (Hypercube, St Gainesville, USA). The number of aromatic and non-aromatic oxygen atoms with their partial charges was recorded.
4.6. Statistical Evaluation
Each experiment was repeated three times with eight replicates. The results were presented as the mean ± standard deviation (SD). The correlation coefficient was calculated using the Spearman method. The statistical significance between groups was evaluated using the Student’s t-test, with a difference considered statistically significant when p < 0.1. Each experiment was repeated three times with eight replicates.
5. Conclusions
Among the 18 clinically applied antibiotics studied, only six demonstrated detectable antioxidant potential. Doxycycline hydrochloride and rifampicin exhibited significantly higher pro-oxidant effects compared to their antioxidant effects, suggesting a contribution to their antibacterial mechanisms. Tebipenem showed notable antioxidant properties, potentially aiding in tissue healing. Most antibiotics did not display significant redox potential, indicating that their antibacterial effects are likely based on mechanisms other than redox modulation. In silico analysis revealed that not all antibiotics adhere to Lipinski’s rule of five, highlighting exceptions in drug design. The position of oxygen atoms on the antibiotic molecule, along with their partial charges, significantly influences the molecule’s antioxidant and pro-oxidant properties. Understanding these mechanisms not only underscores the complexity of antibiotic action but also highlights potential targets for enhancing antibacterial efficacy and addressing bacterial resistance. This study’s findings provide valuable insights into the dual antioxidant and pro-oxidant properties of antibiotics, with implications for therapy and biotechnology. Future research should expand the collection of tested antibiotics for both antioxidant and pro-oxidant effects to proportionally represent each antibiotic group, aiming to achieve a better understanding of these properties and their impact on antibacterial action. Additionally, we plan to study radical production mechanisms in bacterial cultures exposed to homogeneous parameters or using EPR methods.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ph17101257/s1, Table S1: The structures and CAS numbers of evaluated antibiotics.
Author Contributions
Conceptualization, T.M. and M.B.; methodology, T.M. and J.V.; software, T.M. and J.P.; validation, M.B and E.Ü.; formal analysis, M.B., J.P. and E.Ü.; investigation, T.M., M.B. and J.P.; resources, T.M. and J.V.; data curation, M.B., M.M. and E.Ü.; writing—original draft preparation, T.M., M.B. and J.V.; writing—review and editing, J.P., M.M. and J.V.; visualization, T.M. and M.B.; supervision, T.M. and J.P.; project administration, T.M. and E.Ü., funding acquisition, J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Ministry of Education, Research, Development and Youth of the Slovak Republic, grant number: VEGA 1/0230/24.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
Author Jaroslav Polák was employed by the company Helgeheim, Inc. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- WHO Collaborating Centre for Drug Statistics Methodology. Purpose of the ATC/DDD System. Archived from the Original on 14 January 2010. Available online: https://atcddd.fhi.no/atc_ddd_methodology/purpose_of_the_atc_ddd_system/ (accessed on 6 July 2021).
- Reffat, N.; Schwei, R.J.; Griffin, M.; Pop-Vicas, A.; Schulz, L.T.; Michael, S.; Pulia, M.S. A Scoping Review of Bacterial Resistance Among Inpatients Amidst the COVID-19 Pandemic. J. Glob. Antimicrob. Resist. 2024, 38, 49–65. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Nunez, C.; Cesaro, A.; Hancock, R.E.W. Antibiotic failure: Beyond antimicrobial resistance. Drug Resist. Updat. 2023, 71, 101012. [Google Scholar] [CrossRef] [PubMed]
- Prakash, C.; Long, J.; Edelkamp, P.; Knafl, M.; Ahmed, S.; Khawaja, F.; Nastoupil, L.J.; Manzano, J.G.; Mulanovich, V.; Prabhakaran, S.; et al. Prediction of Mortality Following Cytokine Storm in Patients with Hematological Malignancies and COVID-19 Infections. Blood 2022, 140, 10907–10908. [Google Scholar] [CrossRef]
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Westwood, D.; MacFadden, D.R.; Soucy, J.-P.R.; Nick Daneman, N. Bacterial co-infection and secondary infection in patients with COVID-19: A living rapid review and meta-analysis. Clin. Microbiol. Infect. 2020, 26, 1622–1629. [Google Scholar] [CrossRef]
- Lin, J.Y.; Zhu, Z.C.; Zhu, J.; Chen, L.; Hong Du, H. Antibiotic heteroresistance in Klebsiella pneumoniae: Definition, detection methods, mechanisms, and combination therapy. Microbiol. Res. 2024, 283, 127701. [Google Scholar] [CrossRef]
- Orthobullets. Available online: https://www.orthobullets.com/basic-science/9059/antibiotic-classification-and-mechanism (accessed on 15 June 2024).
- Saleem, N.; Ryckaert, F.; Snow, T.A.C.; Satta, G.; Singer, M.; Arulkumaran, N. Mortality and clinical cure rates for pneumonia: A systematic review, meta-analysis, and trial sequential analysis of randomized control trials comparing bactericidal and bacteriostatic antibiotic treatments. Clin. Microbiol. Infect. 2022, 28, 936–945. [Google Scholar] [CrossRef]
- Guillouzo, A.; Guguen-Guillouzo, C. Antibiotics-induced oxidative stress. Curr. Opin. Toxicol. 2020, 20–21, 23–28. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Wierzbowski, J.; Cottarel, G.; Collins, J.J. Mistranslation of membrane proteins and two-component system activation trigger antibiotic-mediated cell death. Cell 2008, 135, 679–690. [Google Scholar] [CrossRef]
- Foti, J.J.; Devadoss, B.; Winkler, J.A.; Collins, J.J.; Walker, G.C. Oxidation of the guanine nucleotide pool underlies cell death by bactericidal antibiotics. Science 2012, 336, 315–319. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X. Contribution of oxidative damage to antimicrobial lethality. Antimicrob. Agents Chemother. 2009, 53, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Van Acker, H.; Coenye, T. The Role of Reactive Oxygen Species in Antibiotic—Mediated Killing of Bacteria. Trends Microbiol. 2017, 25, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Slauch, J.M. How does the oxidative burst of macrophages kill bacteria? Still an open question. Mol. Microbiol. 2011, 80, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Delghandi, P.S.; Soleimani, V.; Bazzaz, B.S.F.; Hosseinzadeh, H. A review on oxidant and antioxidant effects of antibacterial agents: Impacts on bacterial cell death and division and therapeutic effects or adverse reactions in humans. Naunyn Schmiedebergs Arch. Pharmacol. 2023, 396, 2667–2686. [Google Scholar] [CrossRef]
- Barreiro, C.; García-Estrada, C. Proteomics and Penicillium chrysogenum: Unveiling the secrets behind penicillin production. J. Proteom. 2019, 198, 119–131. [Google Scholar] [CrossRef]
- Pathak, A.; Nowell, R.W.; Wilson, C.G.; Ryan, M.J.; Barraclough, T.G. Comparative genomics of Alexander Fleming’s original Penicillium isolate (IMI 15378) reveals sequence divergence of penicillin synthesis genes. Sci. Rep. 2020, 10, 15705. [Google Scholar] [CrossRef]
- Pandey, N.; Cascella, M. Beta Lactam Antibiotics. In Stat Pearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Lin, X.; Kuck, U. Cephalosporins as key lead generation beta-lactam antibiotics. Appl. Microbiol. Biotechnol. 2022, 106, 8007–8020. [Google Scholar] [CrossRef]
- Tan, Q.; Qiu, J.; Luo, X.; Zhang, Y.; Liu, Y.; Chen, Y.; Yuan, J.; Liao, W. Progress in one-pot bioconversion of cephalosporin C to 7-Aminocephalosporanic acid. Curr. Pharm. Biotechnol. 2018, 19, 30–42. [Google Scholar] [CrossRef]
- Pickens, L.B.; Tang, Y. Oxytetracycline biosynthesis. J. Biol. Chem. 2010, 285, 27509–27515. [Google Scholar] [CrossRef]
- Jukes, T.H. Some historical notes on chlortetracycline. Rev. Infect. Dis. 1985, 7, 702–707. [Google Scholar] [CrossRef]
- Ni, H.; Mohsin, A.; Guo, M.; Chu, J.; Zhuang, Y. Two-component system AfrQ1Q2 involved in oxytetracycline biosynthesis of Streptomyces rimosus M4018 in a medium-dependent manner. J. Biosci. Bioeng. 2020, 129, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What we learn from cells culture and in vivo studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Macáková, K.; Mladenka, P.; Filipsky, T.; Říha, M.; Jahodár, L.; Trejtnar, F.; Bovicelli, P.; Silvestri, I.P.; Hrdina, R.; Saso, L. Iron reduction potentiates hydroxyl radical formation only in flavonols. Food Chem. 2012, 135, 2584–2592. [Google Scholar] [CrossRef] [PubMed]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Jukić, D.P.; Rotim, C.; Trebše, P.; Starc, A. Prooxidant activities of antioxidants and their impact on health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef]
- Kutscher, A.H.; Lane, S.L. The clinical toxicity of antibacterial drugs: A comparative review of the literature based on 96,075 cases treated with the sulfonamides and antibiotics. Oral Surg. Oral Med. Oral Path. 1952, 5, 347–352. [Google Scholar] [CrossRef]
- Maliar, T.; Maliarová, M.; Blažková, M.; Kunštek, M.; Uváčková, Ľ.; Viskupičová, J.; Purdešová, A.; Beňovič, P. Simultaneously Determined Antioxidant and Pro-Oxidant Activity of Randomly Selected Plant Secondary Metabolites and Plant Extracts. Molecules 2023, 28, 6890. [Google Scholar] [CrossRef]
- Singh, S.; Khanna, D.; Kalra, S. Minocycline and Doxycycline: More Than Antibiotics. Curr. Mol. Pharmacol. 2021, 14, 1046–1065. [Google Scholar] [CrossRef]
- Pournaras, S.; Koumaki, V.; Spanakis, N.; Gennimata, V.; Tsakris, A. Current perspectives on tigecycline resistance in Enterobacteriaceae: Susceptibility testing issues and mechanisms of resistance. Int. J. Antimicrob. Agents 2016, 48, 11–18. [Google Scholar] [CrossRef]
- Vairo, C.; Vidal, M.V.; Hernandez, R.M.; Igartua, M.; Villullas, S. Colistin- and amikacin-loaded lipid-based drug delivery systems for resistant gram-negative lung and wound bacterial infections. Int. J. Pharm. 2023, 635, 122739. [Google Scholar] [CrossRef]
- Yang, M.; Yuan, F.; Guo, Y.; Wang, S. Efficacy of adding azithromycin to antibiotic prophylaxis in caesarean delivery: A meta-analysis and systematic review. Int. J. Antimicrob. Agents. 2022, 59, 106533. [Google Scholar] [CrossRef]
- Thottathil, S.; Puttaiahgowda, M.Y.; Kanth, S. Advancement and future perspectives on ampicillin-loaded antimicrobial polymers—A review. J. Drug Del. Scien. Technol. 2023, 81, 104227. [Google Scholar] [CrossRef]
- Sodhi, K.K.; Kumar, M.; Singh, D.K. Insight into the amoxicillin resistance, ecotoxicity, and remediation strategies. J. Water Proc, Eng. 2021, 39, 101858. [Google Scholar] [CrossRef]
- Pinart, M.; Kranz, J.; Jensen, K.; Proctor, T.; Naber, K.; Kunath, F.; Wagenlehner, F.; Schmidt, S. Optimal dosage and duration of pivmecillinam treatment for uncomplicated lower urinary tract infections: A systematic review and meta-analysis. Int. J. Infect. Dis. 2017, 58, 96–109. [Google Scholar] [CrossRef] [PubMed]
- El-Gamal, M.I.; Brahim, I.; Hisham, N.; Aladdin, R.; Mohammed, H.; Bahaaeldin, A. Recent updates of carbapenem antibiotics. Eur. J. Med. Chem. 2017, 131, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, D.E.; Tomas, M.; Miller, D.; Tomcsanyi, L.; Signorella, C.; Montepara, C.A.; Covvey, J.R.; Guarascio, A.J. Cephalosporins for the treatment of uncomplicated pyelonephritis: A systematic review. J. Am. Pharm. Assoc. 2023, 63, 1461–1471. [Google Scholar] [CrossRef]
- Verhoef, J.; Gillissen, A. Resistant Haemophilus influenzae in community-acquired respiratory tract infections: A role for cefixime. Int. J. Antimicrob. Agents 2003, 21, 501–509. [Google Scholar] [CrossRef]
- Falagas, M.E.; Trigkidis, K.K.; Vardakas, K.Z. Inhaled antibiotics beyond aminoglycosides, polymyxins and aztreonam: A systematic review. Int. J. Antimicrob. Agents 2015, 45, 221–233. [Google Scholar] [CrossRef]
- Huttner, A.; Bielicki, J.; Clements, M.N.; Frimodt-Møller, N.; Muller, A.E.; Paccaud, J.P.; Mouton, J.W. Oral amoxicillin and amoxicillin–clavulanic acid: Properties, indications and usage. Clin. Microbiol. Infect. 2020, 26, 871–879. [Google Scholar] [CrossRef]
- Ahmed, F.; Mahmud, M.T.; Naher, S.; Rana, M.J.; Ara, R.; Ur-Rahman, K.M.S. Effectiveness of Colistin in carbapenem resistant Acinetobacter baumannii—A systematic review. Health Sci. Rev. 2023, 8, 100113. [Google Scholar] [CrossRef]
- Tajik, S.; Shokri, F.; Rostamnezhad, M.; Khoshnood, S.; Mortazavi, S.M.; Mohammad Sholeh, M.; Kouhsari, M. Fosfomycin: A look at its various aspects. Gene Rep. 2020, 19, 100640. [Google Scholar] [CrossRef]
- Bobba, S.; Khader, S.A. Rifampicin drug resistance and host immunity in tuberculosis: More than meets the eye. Trends Imunnol. 2023, 44, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Zadeh, F.A.; Ebrahimzadeh, F.; Jafari-Sales, A.; Gholami, S. Globally Vibrio cholera antibiotics resistance to RNA and DNA effective antibiotics: A systematic review and meta-analysis. Microb. Pathog. 2022, 172, 105514. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.K.; Singh, D.K. Insight into the fluoroquinolone resistance, sources, ecotoxicity, and degradation with special emphasis on ciprofloxacin. J. Water Proc. Eng. 2021, 43, 102218. [Google Scholar] [CrossRef]
- Dwyer, P.C.; Hogan, M.J.; Stewart, I. An integrated critical thinking framework for the 21st century. Think. Skills Creat. 2014, 12, 43–52. [Google Scholar] [CrossRef]
- Vaishampayan, A.; Grohmann, E. Antimicrobials Functioning through ROS-Mediated Mechanisms: Current Insights. Microorganisms 2021, 10, 61. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Foti, J.R.; Bray, C.B.; Thompson, J.N.; Allgood, F.S. Know thy self, know thy leader: Contributions of a pattern-oriented approach to examining leader perceptions. Leadership Q. 2012, 23, 702–717. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, C.; Jang, H.J.; Kim, B.O.; Bae, H.W.; Chung, I.Y.; Kim, E.S.; Cho, Y.H. Antibacterial strategies inspired by the oxidative stress and response networks. J. Microbiol. 2019, 57, 203–212. [Google Scholar] [CrossRef]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of Acinetobacter baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents Chemother. 2012, 56, 5642–5649. [Google Scholar] [CrossRef]
- Hoeksema, M.; Brul, S.; Ter Kuile, B.H. Influence of Reactive Oxygen Species on De Novo Acquisition of Resistance to Bactericidal Antibiotics. Antimicrob. Agents Chemother. 2018, 25, e02354-17. [Google Scholar] [CrossRef]
- Léger, L.; Budin-Verneuil, A.; Cacaci, M.; Benachour, A.; Hartke, A.; Verneuil, N. β-Lactam Exposure Triggers Reactive Oxygen Species Formation in Enterococcus faecalis via the Respiratory Chain Component DMK. Cell Rep. 2019, 29, 2184–2191.e3. [Google Scholar] [CrossRef] [PubMed]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.M.; Choi, H.Y.; Cho, S.G. The Role of Reactive Oxygen Species (ROS) in the Biological Activities of Metallic Nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Memar, M.Y.; Ghotaslou, R.; Samiei, M.; Adibkia, K. Antimicrobial use of reactive oxygen therapy: Current insights. Infect. Drug Resis. 2018, 11, 567–576. [Google Scholar] [CrossRef]
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).