
Eugenol Inhibits Neutrophils Myeloperoxidase In Vitro and Attenuates LPS-Induced Lung Inflammation in Mice
 and
and 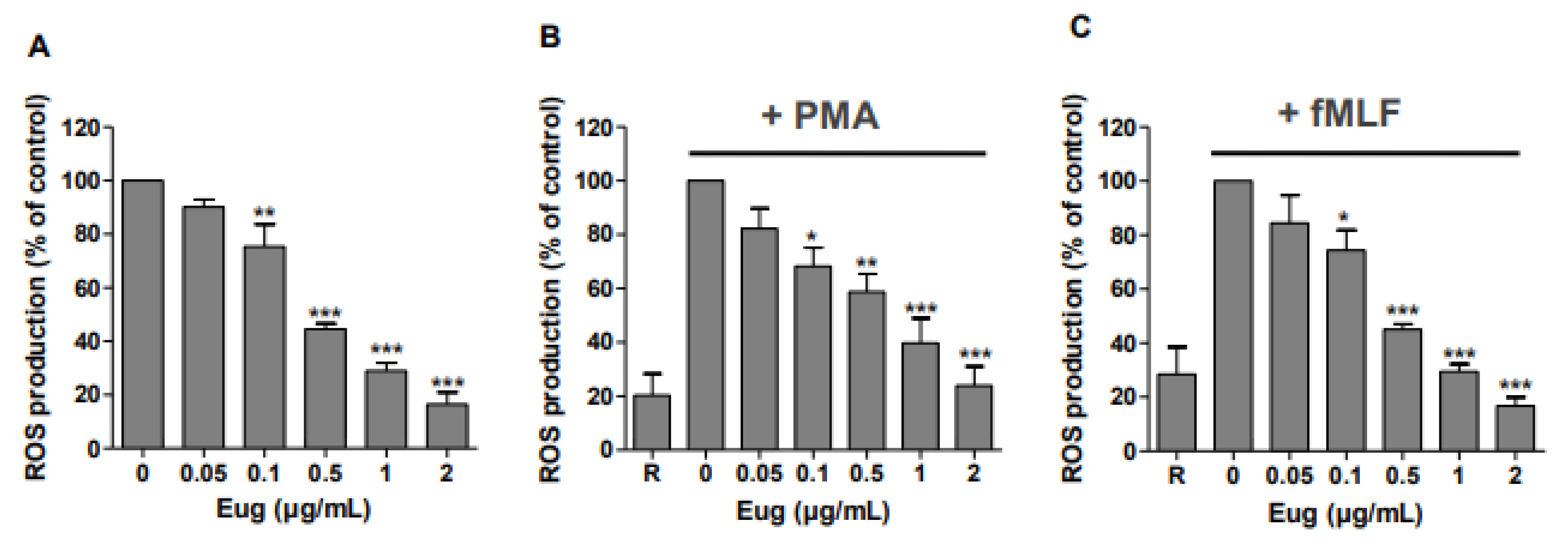{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Eugenol Significantly Decreases ROS Production in Human Neutrophils
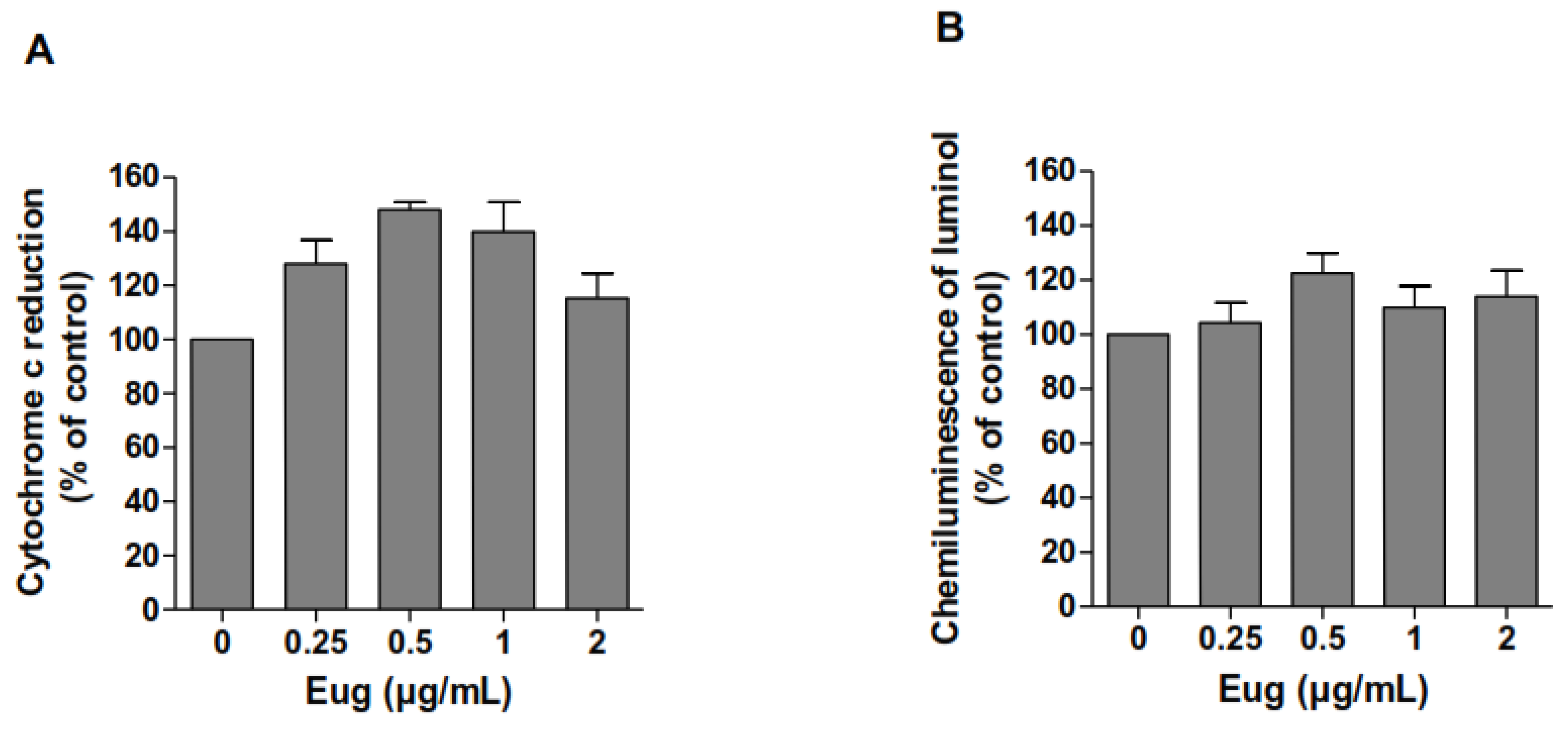
2.2. Eugenol Does Not Scavenge Superoxide Anions nor Hydrogen Peroxide
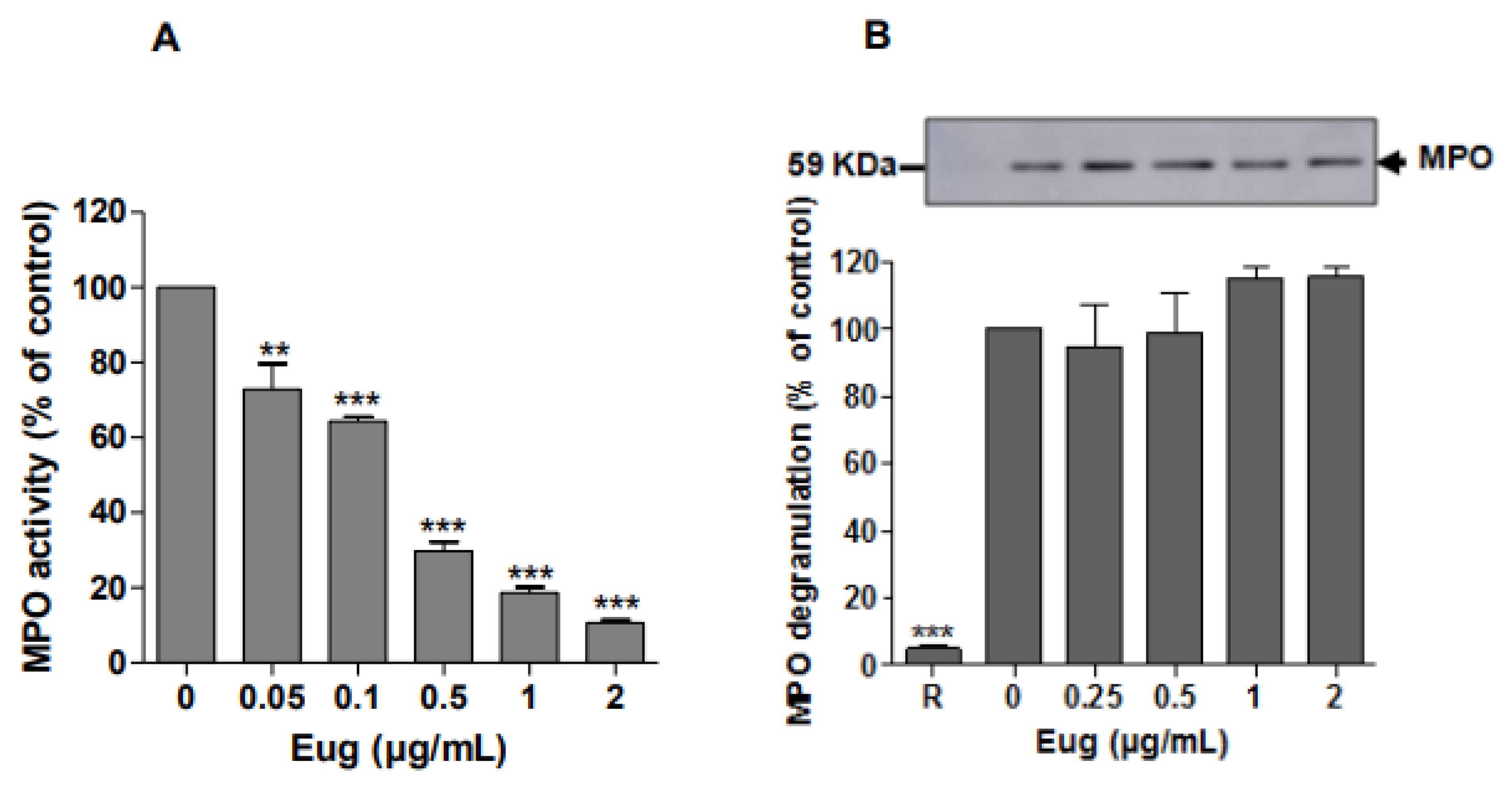
2.3. Eugenol Inhibits Strongly Myeloperoxidase Activity
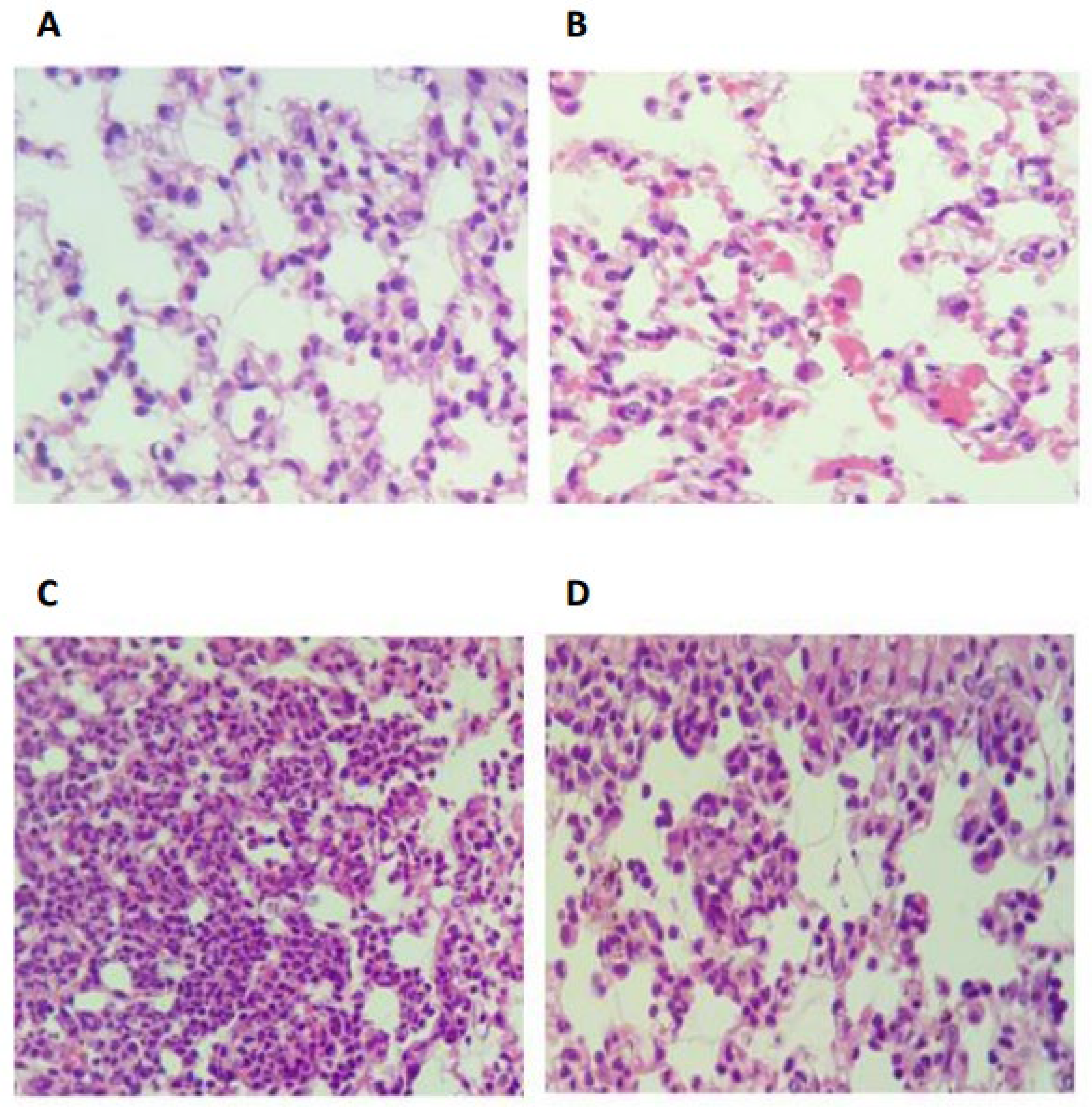
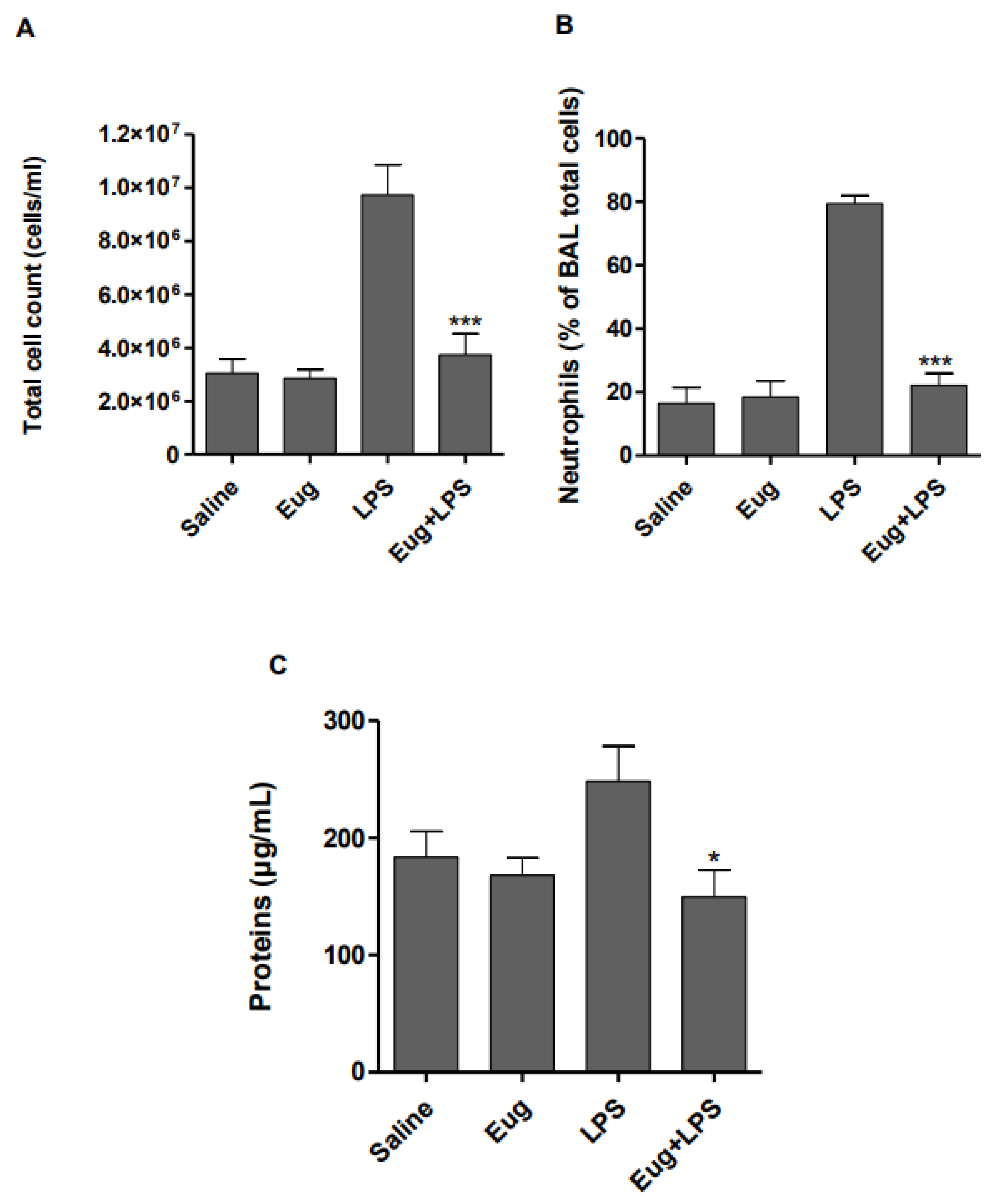
2.4. Eugenol Attenuates the LPS-Induced Inflammatory Response in Mice
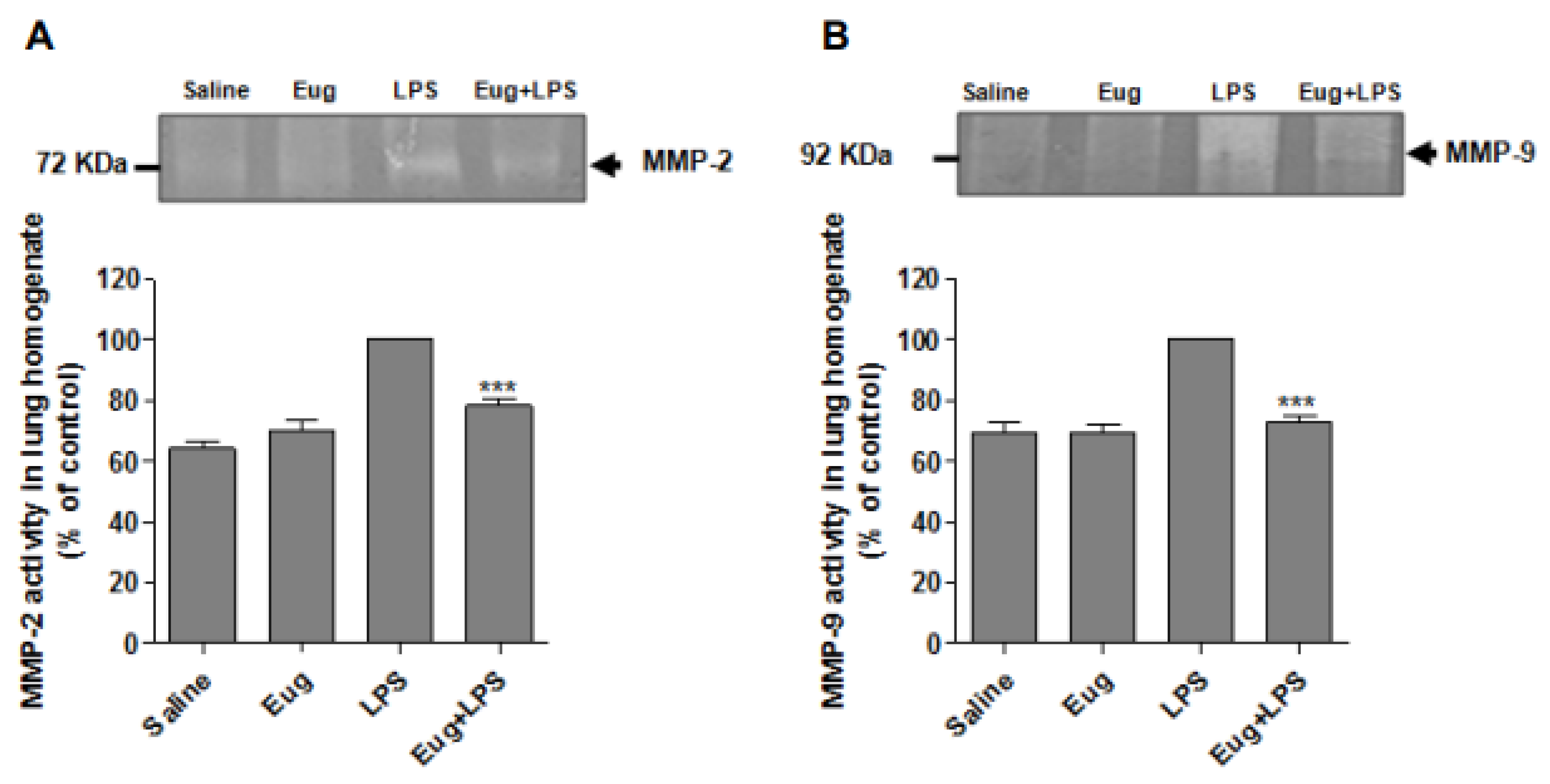
2.5. Eugenol Reduces MMP-2 and MMP-9 Activity in Lung Homogenates
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Eugenol
4.3. Human Neutrophils Isolation
4.4. Luminol-Amplified Chemiluminescence by Human Neutrophils
4.5. Xanthine/Xanthine Oxidase Activity Assay
4.6. Measurement of H2O2 Scavenging Effect
4.7. Measurement of Neutrophils MPO Activity
4.8. Neutrophils Degranulation
4.9. Animals
4.10. Model of Lung Inflammation
4.11. BALF Collection and Cell Counting
4.12. Histological Study
4.13. Gelatin Zymography
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Srivastava, K.K.; Kumar, R. Stress, oxidative injury and disease. Indian J. Clin. Biochem. 2015, 30, 3–10. [Google Scholar] [CrossRef]
- MacNee, W. Pathogenesis of chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2005, 2, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef]
- Lazarevic-Pasti, T.; Leskovac, A.; Vasic, V. Myeloperoxidase inhibitors as potential drugs. Curr. Drug Metab. 2015, 16, 168–190. [Google Scholar] [CrossRef]
- Quaderi, S.A.; Hurst, J.R. The unmet global burden of COPD. Glob. Health Epidemiol. Genom. 2018, 3, e4. [Google Scholar] [CrossRef] [PubMed]
- Balkissoon, R. Stem Cell Therapy for COPD: Where are we? Chronic Obstructive Pulmonary Diseases. J. COPD Found. 2018, 5, 148. [Google Scholar]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Rollinger, J.M. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef]
- Teplova, V.V.; Isakova, E.P.; Klein, O.I.; Dergachova, D.I.; Gessler, N.N.; Deryabina, Y.I. Natural polyphenols: Biological activity, pharmacological potential, means of metabolic engineering. Appl. Biochem. Microbiol. 2018, 54, 221–237. [Google Scholar] [CrossRef]
- Kamatou, G.P.; Vermaak, I.; Viljoen, A.M. Eugenol—From the remote Maluku Islands to the international market place: A review of a remarkable and versatile molecule. Molecules 2012, 17, 6953–6981. [Google Scholar] [CrossRef]
- Nejad, S.M.; Ozgunes, H.; Basaran, N. Pharmacological and toxicological properties of eugenol. Turk. J. Pharm. Sci. 2017, 14, 201–206. [Google Scholar] [CrossRef]
- Chniguir, A.; Pintard, C.; Liu, D.; Dang, P.M.C.; El-Benna, J.; Bachoual, R. Eugenol prevents fMLF-induced superoxide anion production in human neutrophils by inhibiting ERK1/2 signaling pathway and p47phox phosphorylation. Sci. Rep. 2019, 9, 18540. [Google Scholar] [CrossRef] [PubMed]
- Pramod, K.; Ansari, S.H.; Ali, J. Eugenol: A natural compound with versatile pharmacological actions. Nat. Prod. Commun. 2010, 5, 1999–2006. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J.; Kettle, A.J.; Rosen, H.; Winterbourn, C.C.; Nauseef, W.M. Myeloperoxidase: A front-line defender against phagocytosed microorganisms. J. Leukoc. Biol. 2013, 93, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Alsahli, M.A.; Rahmani, A.H. Myeloperoxidase as an active disease biomarker: Recent biochemical and pathological perspectives. Med. Sci. 2018, 6, 33. [Google Scholar] [CrossRef]
- Kim, H.P.; Lim, H.; Kwon, Y.S. Therapeutic potential of medicinal plants and their constituents on lung inflammatory disorders. Biomol. Ther. 2017, 25, 91. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M.C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Glennon-Alty, L.; Hackett, A.P.; Chapman, E.A.; Wright, H.L. Neutrophils and redox stress in the pathogenesis of autoimmune disease. Free Radic. Biol. Med. 2018, 125, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Chen, W.S.; Chueng, A.L.; Dunne, A.A.; Seredenina, T.; Filippova, A.; Allan, C. Discovery of GSK2795039, a novel small molecule NADPH oxidase 2 inhibitor. Antioxid. Redox Signal. 2015, 23, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Bréchard, S.; Tschirhart, E.J. Regulation of superoxide production in neutrophils: Role of calcium influx. J. Leukoc. Biol. 2008, 84, 1223–1237. [Google Scholar] [CrossRef]
- Bedouhène, S.; Moulti-Mati, F.; Hurtado-Nedelec, M.; Dang, P.M.C.; El-Benna, J. Luminol-amplified chemiluminescence detects mainly superoxide anion produced by human neutrophils. Am. J. Blood Res. 2017, 7, 41. [Google Scholar]
- Al-Shehri, S.S. Reactive oxygen and nitrogen species and innate immune response. Biochimie 2021, 181, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Frangie, C.; Daher, J. Role of myeloperoxidase in inflammation and atherosclerosis. Biomed. Rep. 2022, 16, 53. [Google Scholar] [CrossRef] [PubMed]
- Antonelou, M.; Michaëlsson, E.; Wang, C.J.; Walker, L.; Unwin, R.; Salama, A. Therapeutic myeloperoxidase inhibition attenuates neutrophil activation, ANCA-mediated endothelial damage and crescentic glomrullonephritis. Nephrol. Dial. Transplant. 2019, 34 (Suppl. S1), gfz106.FP192. [Google Scholar] [CrossRef]
- Pérez-Rosés, R.; Risco, E.; Vila, R.; Peñalver, P.; Cañigueral, S. Effect of essential oils on the activity of human neutrophil myeloperoxidase in vitro. Planta Medica 2012, 60, 70. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Frevert, C.W.; Martin, T.R. Animal models of acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, 379–399. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, H.M. Dynamic lipopolysaccharide transfer cascade to TLR4/MD2 complex via LBP and CD14. BMB Rep. 2017, 50, 55. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Q.; Mo, Z.Z.; Liao, J.B.; Feng, X.X.; Liang, Y.Z.; Zhang, X.; Lai, X.P. Usnic acid protects LPS-induced acute lung injury in mice through attenuating inflammatory responses and oxidative stress. Int. Immunopharmacol. 2014, 22, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Su, S.C.; Hua, K.F.; Lee, H.; Chao, L.K.; Tan, S.K.; Yang, S.F.; Hsu, H.Y. LTA and LPS mediated activation of protein kinases in the regulation of inflammatory cytokines expression in macrophages. Clin. Chim. Acta 2006, 374, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Tazzyman, S.; Barry, S.T.; Ashton, S.; Wood, P.; Blakey, D.; Lewis, C.E.; Murdoch, C. Inhibition of neutrophil infiltration into A549 lung tumors in vitro and in vivo using a CXCR2-specific antagonist is associated with reduced tumor growth. Int. J. Cancer 2011, 129, 847–858. [Google Scholar] [CrossRef]
- Folkesson, H.G.; Matthay, M.A.; Hebert, C.A.; Broaddus, V.C. Acid aspiration-induced lung injury in rabbits is mediated by interleukin-8-dependent mechanisms. J. Clin. Investig. 1995, 96, 107–116. [Google Scholar] [CrossRef]
- Johnson, J.L. Matrix metalloproteinases and their inhibitors in cardiovascular pathologies: Current knowledge and clinical potential. Met. Med. 2014, 1, 21–36. [Google Scholar]
- Klein, T.; Bischoff, R. Physiology and pathophysiology of matrix metalloproteases. Amino Acids 2011, 41, 271–290. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Fanjul-Fernández, M.; Folgueras, A.R.; Cabrera, S.; López-Otín, C. Matrix metalloproteinases: Evolution, gene regulation and functional analysis in mouse models. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2010, 1803, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Mohan, V.; Talmi-Frank, D.; Arkadash, V.; Papo, N.; Sagi, I. Matrix metalloproteinase protein inhibitors: Highlighting a new beginning for metalloproteinases in medicine. Met. Med. 2016, 3, 31. [Google Scholar]
- Nam, H.; Kim, M.M. Eugenol with antioxidant activity inhibits MMP-9 related to metastasis in human fibrosarcoma cells. Food Chem. Toxicol. 2013, 55, 106–112. [Google Scholar] [CrossRef]
- Abdullah, M.L.; Hafez, M.M.; Al-Hoshani, A.; Al-Shabanah, O. Anti-metastatic and anti-proliferative activity of eugenol against triple negative and HER2 positive breast cancer cells. BMC Complement. Altern. Med. 2018, 18, 321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Reddy, N. Bioactive molecules from medicinal herbs for life threatening diseases. J. Mol. Sciences. 2018, 2, 4. [Google Scholar]
- Yoon, J.H.; Baek, S.J. Molecular targets of dietary polyphenols with anti-inflammatory properties. Yonsei Med. J. 2005, 46, 585–596. [Google Scholar] [CrossRef]
- Mileo, A.M.; Miccadei, S. Polyphenols as modulator of oxidative stress in cancer disease: New therapeutic strategies. Oxid. Med. Cell. Longev. 2016, 2016, 6475624. [Google Scholar] [CrossRef]
- Daniel, A.N.; Sartoretto, S.M.; Schmidt, G.; Caparroz-Assef, S.M.; Bersani-Amado, C.A.; Cuman, R.K.N. Anti-inflammatory and antinociceptive activities A of eugenol essential oil in experimental animal models. Rev. Bras. Farmacogn. 2009, 19, 212–217. [Google Scholar] [CrossRef]
- Fangjun, L.; Zhijia, Y. Tumor suppressive roles of eugenol in human lung cancer cells. Thorac. Cancer 2018, 9, 25–29. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Dang, P.M.C. Analysis of protein phosphorylation in human neutrophils. Neutrophil Methods Protoc. 2007, 412, 85–96. [Google Scholar]
- Udby, L.; Borregaard, N. Subcellular fractionation of human neutrophils and analysis of subcellular markers. Neutrophil Methods Protoc. 2007, 412, 35–56. [Google Scholar]
- Ciccocioppo, R.; Di Sabatino, A.; Bauer, M.; Della Riccia, D.N.; Bizzini, F.; Biagi, F.; Schuppan, D. Matrix metalloproteinase pattern in celiac duodenal mucosa. Lab. Investig. 2005, 85, 397–407. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chniguir, A.; Saguem, M.H.; Dang, P.M.-C.; El-Benna, J.; Bachoual, R. Eugenol Inhibits Neutrophils Myeloperoxidase In Vitro and Attenuates LPS-Induced Lung Inflammation in Mice. Pharmaceuticals 2024, 17, 504. https://doi.org/10.3390/ph17040504
Chniguir A, Saguem MH, Dang PM-C, El-Benna J, Bachoual R. Eugenol Inhibits Neutrophils Myeloperoxidase In Vitro and Attenuates LPS-Induced Lung Inflammation in Mice. Pharmaceuticals. 2024; 17(4):504. https://doi.org/10.3390/ph17040504
Chicago/Turabian StyleChniguir, Amina, Mohamed Hedi Saguem, Pham My-Chan Dang, Jamel El-Benna, and Rafik Bachoual. 2024. "Eugenol Inhibits Neutrophils Myeloperoxidase In Vitro and Attenuates LPS-Induced Lung Inflammation in Mice" Pharmaceuticals 17, no. 4: 504. https://doi.org/10.3390/ph17040504