
R-Baclofen Treatment Corrects Autistic-like Behavioral Deficits in the RjIbm(m):FH Fawn-Hooded Rat Strain
 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
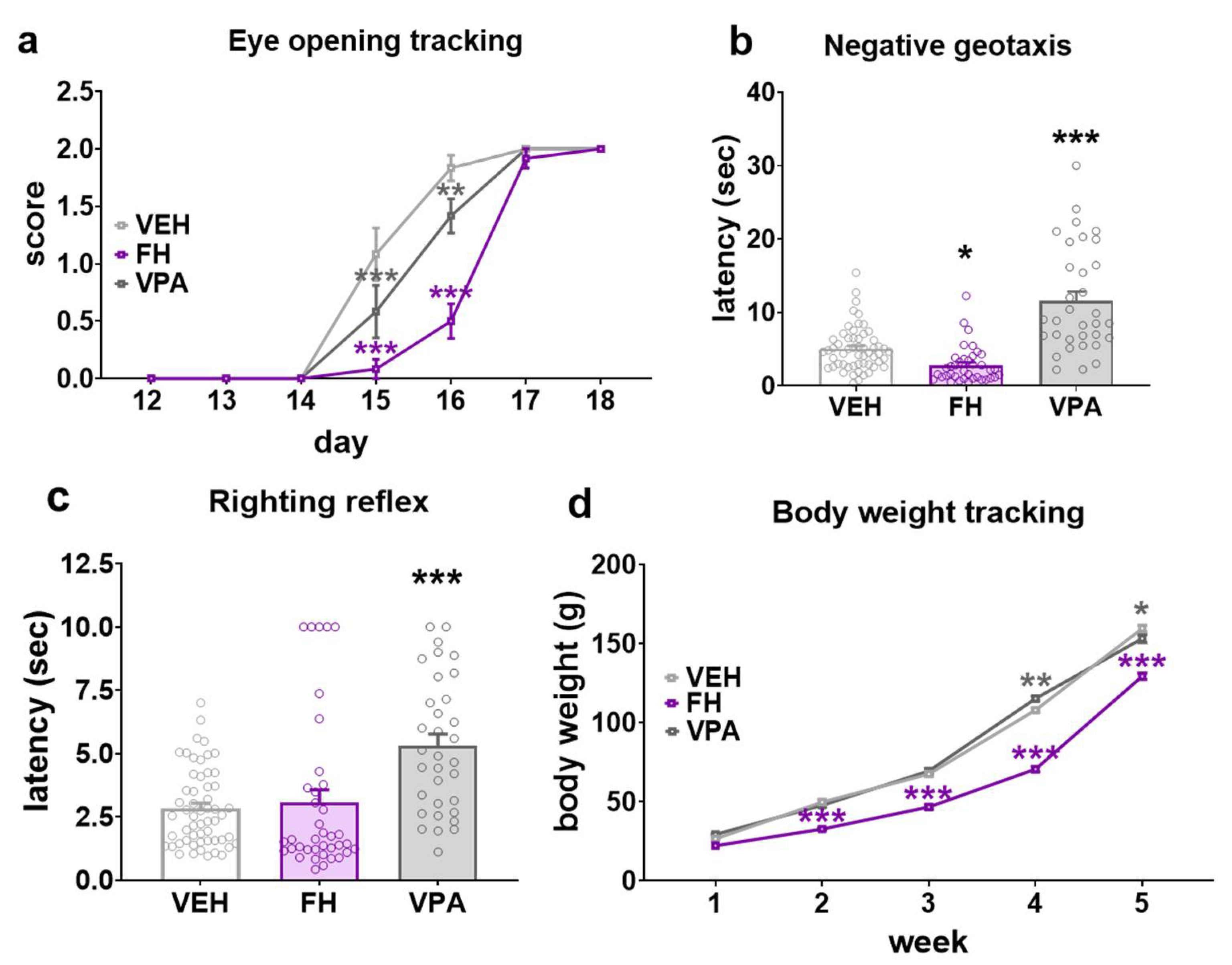
2.1. Early Development
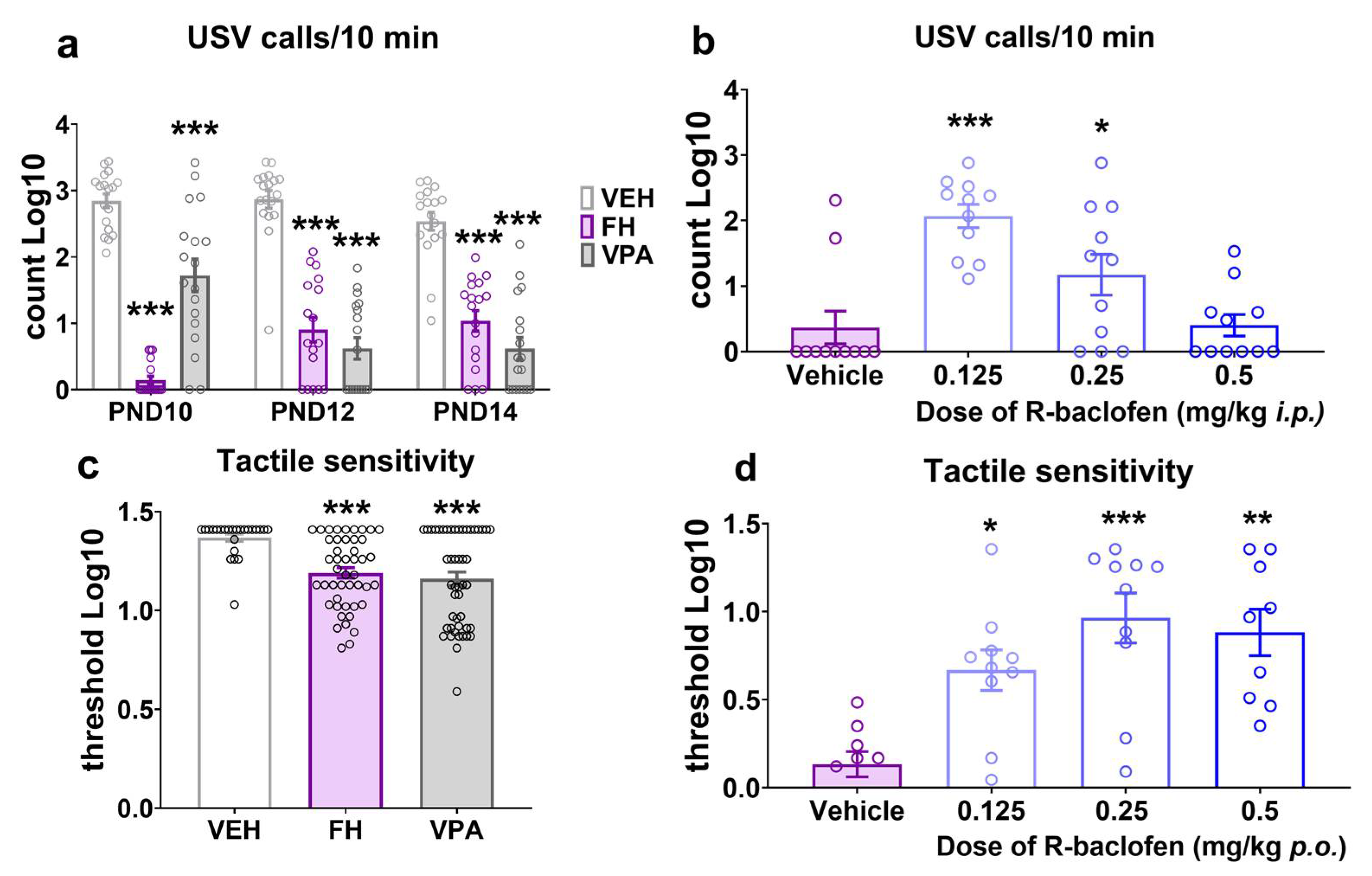
2.2. Ultrasonic Vocalization in Rat Pups
2.3. Tactile Sensitivity
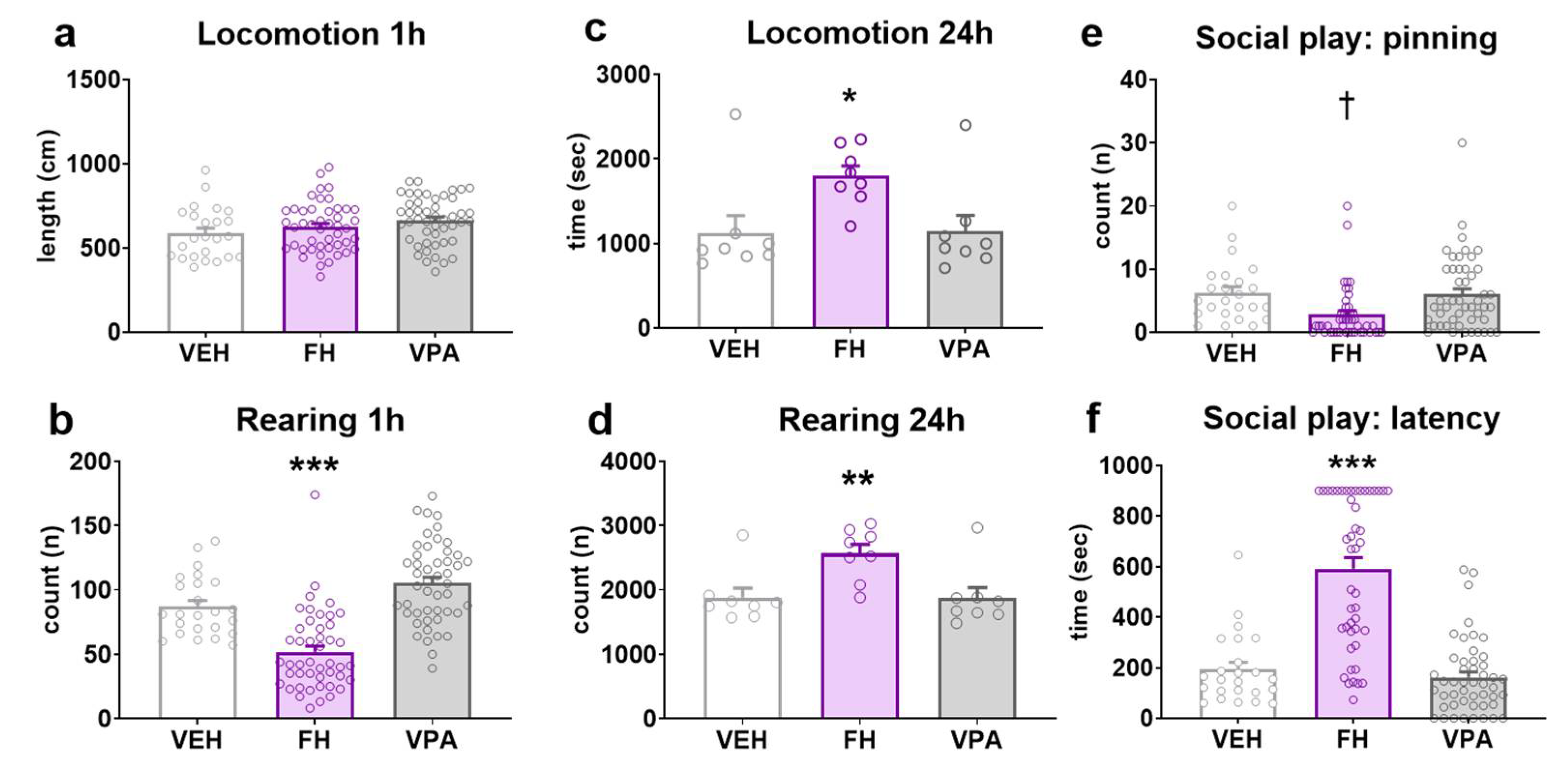
2.4. Spontaneous Motor Activity
2.5. General Activity in the Home Cage
2.6. Juvenile Social Play Behavior
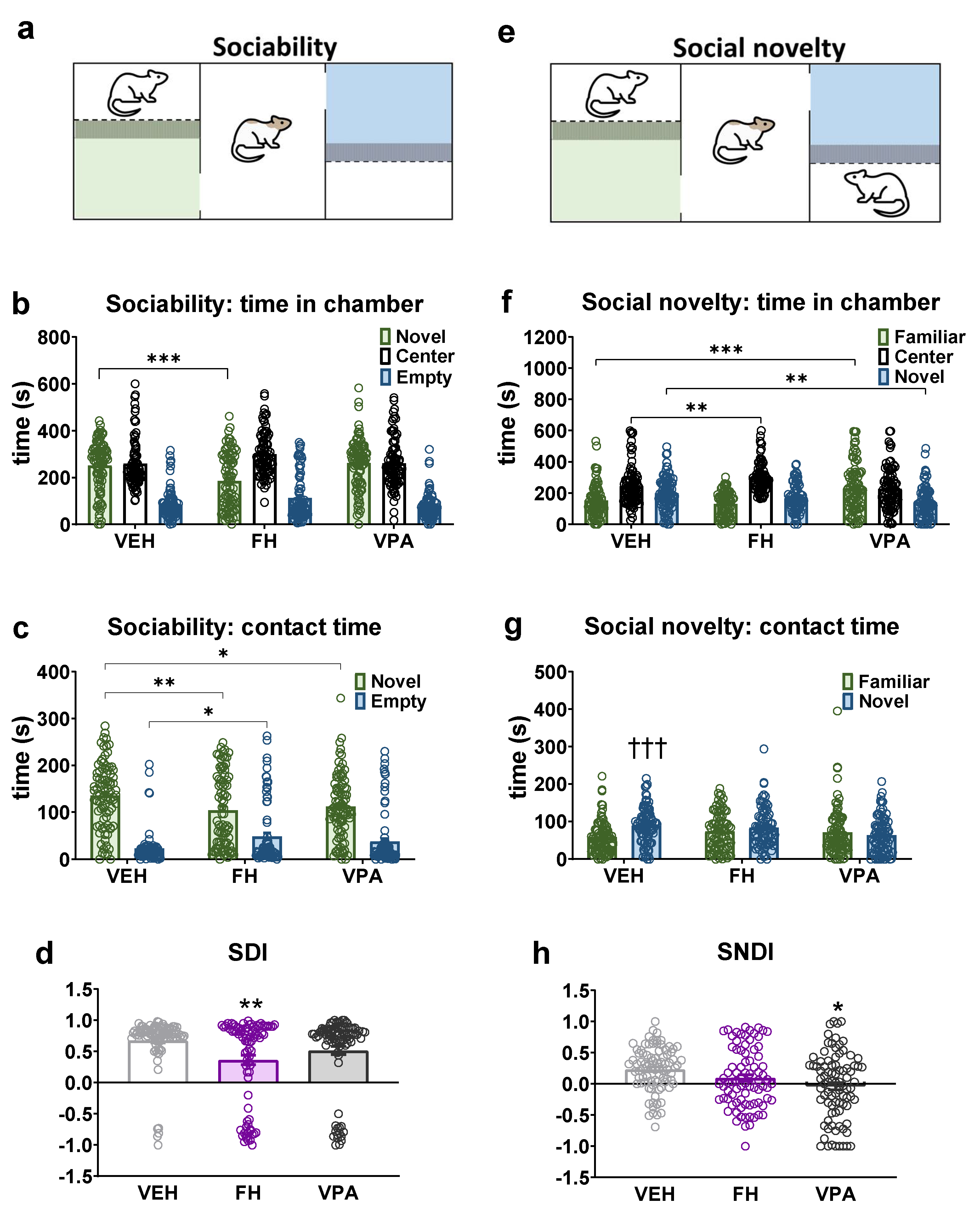
2.7. Sociability and Social Novelty Test
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Husbandry
4.2. Drug Administrations
4.3. Developmental Observations
4.4. Maternal Deprivation-Induced Rat Pup USV
4.5. Spontaneous Locomotor Activity
4.6. General Home Cage Activity
4.7. Juvenile Social Play
4.8. Tactile Sensitivity
4.9. Three-Chamber Sociability and Social Novelty Test
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aulakh, C.S.; Hill, J.L.; Murphy, D.L. Attenuation of hypercortisolemia in fawn-hooded rats by antidepressant drugs. Eur. J. Pharmacol. 1993, 240, 85–88. [Google Scholar] [CrossRef] [PubMed]
- van Dokkum, R.P.; Sun, C.W.; Provoost, A.P.; Jacob, H.J.; Roman, R.J. Altered renal hemodynamics and impaired myogenic responses in the fawn-hooded rat. Am. J. Physiol. 1999, 276, R855–R863. [Google Scholar] [CrossRef] [PubMed]
- Schaap, M.W.; van Oostrom, H.; Arndt, S.S.; Hellebrekers, L.J. Bleeding Diathesis in Fawn Hooded Rats-Possible Implications for Invasive Procedures and Refinement Strategies. Animals 2015, 5, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Aulakh, C.S.; Hill, J.L.; Murphy, D.L. Fawn hooded rats are subsensitive to the food intake suppressant effects of 5-HT agonists. Psychopharmacology 1988, 94, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Aulakh, C.S.; Tolliver, T.; Wozniak, K.M.; Hill, J.L.; Murphy, D.L. Functional and biochemical evidence for altered serotonergic function in the fawn-hooded rat strain. Pharmacol. Biochem. Behav. 1994, 49, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Azechi, H.; Hakamada, K.; Yamamoto, T. A new inbred strain of Fawn-Hooded rats demonstrates mania-like behavioural and monoaminergic abnormalities. IBRO Rep. 2019, 7, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.S.; Huang, S.; Fong, G. Effects of isolation-rearing on acoustic startle and pre-pulse inhibition in Wistar and fawn hooded rats. Ann. N. Y. Acad. Sci. 1997, 821, 542–544. [Google Scholar] [CrossRef] [PubMed]
- Calcagnetti, D.J.; Schechter, M.D. Deficits in shock-induced freezing and naltrexone enhancement of freezing in fawn hooded rats. Brain Res. Bull. 1994, 35, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, S.; Oliva, J.M.; Pérez-Rial, S.; Palomo, T.; Manzanares, J. Differences in basal cannabinoid CB1 receptor function in selective brain areas and vulnerability to voluntary alcohol consumption in Fawn Hooded and Wistar rats. Alcohol. Alcohol. 2004, 39, 297–302. [Google Scholar] [CrossRef]
- Tizabi, Y.; Getachew, B.; Rezvani, A.H.; Hauser, S.R.; Overstreet, D.H. Antidepressant-like effects of nicotine and reduced nicotinic receptor binding in the Fawn-Hooded rat, an animal model of co-morbid depression and alcoholism. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 398–402. [Google Scholar] [CrossRef]
- Hu, B.; Das, P.; Lv, X.; Shi, M.; Aa, J.; Wang, K.; Duan, L.; Gilbert, J.A.; Nie, Y.; Wu, X.L. Effects of ‘Healthy’ Fecal Microbiota Transplantation against the Deterioration of Depression in Fawn-Hooded Rats. mSystems 2022, 7, e0021822. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, D.H.; Rezvani, A.H.; Parsian, A. Behavioural features of alcohol-preferring rats: Focus on inbred strains. Alcohol. Alcohol. 1999, 34, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Knapp, D.J.; Harper, K.M.; Melton, J.; Breese, G. Comparative effects of stressors on behavioral and neuroimmune responses of fawn-hooded (FH/Wjd) and Wistar rats: Implications for models of depression. J. Neuroimmunol. 2018, 322, 74–80. [Google Scholar] [CrossRef]
- Ryan, J.; Bloch, K.; Archer, S.L. Rodent models of pulmonary hypertension: Harmonisation with the world health organisation’s categorisation of human PH. Int. J. Clin. Pract. Suppl. 2011, 172, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, A.H.; Parsian, A.; Overstreet, D.H. The Fawn-Hooded (FH/Wjd) rat: A genetic animal model of comorbid depression and alcoholism. Psychiatr. Genet. 2002, 12, 1–16. [Google Scholar] [CrossRef]
- Altemus, M.; Smith, M.A.; Diep, V.; Aulakh, C.S.; Murphy, D.L. Increased mRNA for corticotrophin releasing hormone in the amygdala of fawn-hooded rats: A potential animal model of anxiety. Anxiety 1994, 1, 251–257. [Google Scholar] [CrossRef]
- Overstreet, D.H.; Rezvani, A.H.; Djouma, E.; Parsian, A.; Lawrence, A.J. Depressive-like behavior and high alcohol drinking co-occur in the FH/WJD rat but appear to be under independent genetic control. Neurosci. Biobehav. Rev. 2007, 31, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Lahmame, A.; Gomez, F.; Armario, A. Fawn-hooded rats show enhanced active behaviour in the forced swimming test, with no evidence for pituitary-adrenal axis hyperactivity. Psychopharmacology 1996, 125, 74–78. [Google Scholar] [CrossRef]
- Penner, J.D.; Prieur, D.J. Fibroblasts of fawn-hooded rats. Vet. Pathol. 1988, 25, 169–171. [Google Scholar] [CrossRef]
- Overstreet, D.H.; Rezvani, A.H. Behavioral differences between two inbred strains of Fawn-Hooded rat: A model of serotonin dysfunction. Psychopharmacology 1996, 128, 328–330. [Google Scholar] [CrossRef]
- Kantor, S.; Anheuer, Z.E.; Bagdy, G. High social anxiety and low aggression in Fawn-Hooded rats. Physiol. Behav. 2000, 71, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Kantor, S.; Graf, M.; Anheuer, Z.E.; Bagdy, G. Rapid desensitization of 5-HT(1A) receptors in Fawn-Hooded rats after chronic fluoxetine treatment. Eur. Neuropsychopharmacol. 2001, 11, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Moy, S.S.; Nadler, J.J.; Perez, A.; Barbaro, R.P.; Johns, J.M.; Magnuson, T.R.; Piven, J.; Crawley, J.N. Sociability and preference for social novelty in five inbred strains: An approach to assess autistic-like behavior in mice. Genes Brain Behav. 2004, 3, 287–302. [Google Scholar] [CrossRef]
- Moy, S.S.; Nadler, J.J.; Young, N.B.; Perez, A.; Holloway, L.P.; Barbaro, R.P.; Barbaro, J.R.; Wilson, L.M.; Threadgill, D.W.; Lauder, J.M.; et al. Mouse behavioral tasks relevant to autism: Phenotypes of 10 inbred strains. Behav. Brain Res. 2007, 176, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Berg, E.L.; Copping, N.A.; Rivera, J.K.; Pride, M.C.; Careaga, M.; Bauman, M.D.; Berman, R.F.; Lein, P.J.; Harony-Nicolas, H.; Buxbaum, J.D.; et al. Developmental social communication deficits in the Shank3 rat model of phelan-mcdermid syndrome and autism spectrum disorder. Autism Res. 2018, 11, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.M.; Green, J.R.; Veeraragavan, S.; Yuva, L.; McCoy, A.; Wu, Y.; Warren, J.; Little, L.; Ji, D.; Cui, X.; et al. Fmr1 and Nlgn3 knockout rats: Novel tools for investigating autism spectrum disorders. Behav. Neurosci. 2014, 128, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Martin Lorenzo, S.; Muniz Moreno, M.D.M.; Atas, H.; Pellen, M.; Nalesso, V.; Raffelsberger, W.; Prevost, G.; Lindner, L.; Birling, M.C.; Menoret, S.; et al. Changes in social behavior with MAPK2 and KCTD13/CUL3 pathways alterations in two new outbred rat models for the 16p11.2 syndromes with autism spectrum disorders. Front. Neurosci. 2023, 17, 1148683. [Google Scholar] [CrossRef]
- Saxena, K.; Webster, J.; Hallas-Potts, A.; Mackenzie, R.; Spooner, P.A.; Thomson, D.; Kind, P.; Chattarji, S.; Morris, R.G.M. Experiential contributions to social dominance in a rat model of fragile-X syndrome. Proc. Biol. Sci. 2018, 285, 20180294. [Google Scholar]
- Schneider, M.; de Vries, P.J.; Schönig, K.; Rößner, V.; Waltereit, R. mTOR inhibitor reverses autistic-like social deficit behaviours in adult rats with both Tsc2 haploinsufficiency and developmental status epilepticus. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Ku, K.M.; Weir, R.K.; Silverman, J.L.; Berman, R.F.; Bauman, M.D. Behavioral Phenotyping of Juvenile Long-Evans and Sprague-Dawley Rats: Implications for Preclinical Models of Autism Spectrum Disorders. PLoS ONE 2016, 11, e0158150. [Google Scholar] [CrossRef]
- Brust, V.; Schindler, P.M.; Lewejohann, L. Lifetime development of behavioural phenotype in the house mouse (Mus musculus). Front. Zool. 2015, 12, S17. [Google Scholar] [CrossRef] [PubMed]
- Homberg, J.R.; Wöhr, M.; Alenina, N. Comeback of the Rat in Biomedical Research. ACS Chem. Neurosci. 2017, 8, 900–903. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Health Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; pp. 50–59. [Google Scholar]
- Baska, F.; Bozó, É.; Szeleczky, Z.; Szántó, G.; Vukics, K.; Szakács, Z.; Domány-Kovács, K.; Kurkó, D.; Vass, E.; Thán, M.; et al. Discovery and Characterization of RGH-122, a Potent, Selective, and Orally Bioavailable V1a Receptor Antagonist. J. Med. Chem. 2024, 67, 643–673. [Google Scholar] [CrossRef] [PubMed]
- Mueggler, T.; D’Souza, D.D.; Biemans, B.; Bruns, A.; Künnecke, B.; Schnider, P.; Grundschober, C.; von Kienlin, M. V1a antagonism normalizes a social brain network in valproate rat model of autism revealed by functional MRI. Proc. Intl. Soc. Mag. Reson. Med. 2014, 22, 0223. [Google Scholar]
- Mabunga, D.F.; Gonzales, E.L.; Kim, J.W.; Kim, K.C.; Shin, C.Y. Exploring the Validity of Valproic Acid Animal Model of Autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Nicolini, C.; Fahnestock, M. The valproic acid-induced rodent model of autism. Exp. Neurol. 2018, 299, 217–227. [Google Scholar] [CrossRef]
- Frye, R.E. Clinical potential, safety, and tolerability of arbaclofen in the treatment of autism spectrum disorder. Drug Healthc. Patient Saf. 2014, 6, 69–76. [Google Scholar] [CrossRef]
- Huang, Q.; Pereira, A.C.; Velthuis, H.; Wong, N.M.L.; Ellis, C.L.; Ponteduro, F.M.; Dimitrov, M.; Kowalewski, L.; Lythgoe, D.J.; Rotaru, D.; et al. GABAB receptor modulation of visual sensory processing in adults with and without autism spectrum disorder. Sci. Transl. Med. 2022, 14, eabg7859. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Velthuis, H.; Pereira, A.C.; Ahmad, J.; Cooke, S.F.; Ellis, C.L.; Ponteduro, F.M.; Puts, N.A.J.; Dimitrov, M.; Batalle, D.; et al. Exploratory evidence for differences in GABAergic regulation of auditory processing in autism spectrum disorder. Transl. Psychiatry 2023, 13, 320. [Google Scholar]
- Kordás, K.; Kis-Varga, Á.; Varga, A.; Eldering, H.; Bulthuis, R.; Lendvai, B.; Lévay, G.; Román, V. Measuring sociability of mice using a novel three-chamber apparatus and algorithm of the LABORAS™ system. J. Neurosci. Methods 2020, 343, 108841. [Google Scholar]
- Möhrle, D.; Wang, W.; Whitehead, S.N.; Schmid, S. GABAB Receptor Agonist R-Baclofen Reverses Altered Auditory Reactivity and Filtering in the Cntnap2 Knock-Out Rat. Front. Integr. Neurosci. 2021, 15, 710593. [Google Scholar] [CrossRef]
- Qin, M.; Huang, T.; Kader, M.; Krych, L.; Xia, Z.; Burlin, T.; Zeidler, Z.; Zhao, T.; Smith, C.B. R-Baclofen Reverses a Social Behavior Deficit and Elevated Protein Synthesis in a Mouse Model of Fragile X Syndrome. Int. J. Neuropsychopharmacol. 2015, 18, pyv034. [Google Scholar] [CrossRef] [PubMed]
- Sharghi, S.; Flunkert, S.; Daurer, M.; Rabl, R.; Chagnaud, B.P.; Leopoldo, M.; Lacivita, E.; Hutter-Paier, B.; Prokesch, M. Evaluating the effect of R-Baclofen and LP-211 on autistic behavior of the BTBR and Fmr1-KO mouse models. Front. Neurosci. 2023, 17, 1087788. [Google Scholar] [CrossRef]
- Silverman, J.L.; Pride, M.C.; Hayes, J.E.; Puhger, K.R.; Butler-Struben, H.M.; Baker, S.; Crawley, J.N. GABAB Receptor Agonist R-Baclofen Reverses Social Deficits and Reduces Repetitive Behavior in Two Mouse Models of Autism. Neuropsychopharmacology 2015, 40, 2228–2239. [Google Scholar] [CrossRef]
- Trezza, V.; Baarendse, P.J.; Vanderschuren, L.J. The pleasures of play: Pharmacological insights into social reward mechanisms. Trends Pharmacol. Sci. 2010, 31, 463–469. [Google Scholar] [CrossRef]
- Schneider, T.; Przewłocki, R. Behavioral alterations in rats prenatally exposed to valproic acid: Animal model of autism. Neuropsychopharmacology 2005, 30, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Modi, M.E.; Brooks, J.M.; Guilmette, E.R.; Beyna, M.; Graf, R.; Reim, D.; Schmeisser, M.J.; Boeckers, T.M.; O’Donnell, P.; Buhl, D.L. Hyperactivity and Hypermotivation Associated with Increased Striatal mGluR1 Signaling in a Shank2 Rat Model of Autism. Front. Mol. Neurosci. 2018, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Vitor-Vieira, F.; Vilela, F.C.; Giusti-Paiva, A. Hyperactivation of the amygdala correlates with impaired social play behavior of prepubertal male rats in a maternal immune activation model. Behav. Brain Res. 2021, 414, 113503. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N. Mouse behavioral assays relevant to the symptoms of autism. Brain Pathol. 2007, 17, 448–459. [Google Scholar] [CrossRef]
- Bambini-Junior, V.; Rodrigues, L.; Behr, G.A.; Moreira, J.C.; Riesgo, R.; Gottfried, C. Animal model of autism induced by prenatal exposure to valproate: Behavioral changes and liver parameters. Brain Res. 2011, 1408, 8–16. [Google Scholar] [CrossRef]
- Li, X.; Nie, Y.; Qiu, Z.; Wang, S. Human MECP2 transgenic rats show increased anxiety, severe social deficits, and abnormal prefrontal neural oscillation stability. Biochem. Biophys. Res. Commun. 2023, 648, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Qi, S.; Calhoun, V.; Dai, J.; Yu, B.; Zhang, K.; Pei, M.; Li, C.; Wei, Y.; Jiang, R.; et al. Aberrant brain functional and structural developments in MECP2 duplication rats. Neurobiol. Dis. 2022, 173, 105838. [Google Scholar] [CrossRef] [PubMed]
- Lever, C.; Burton, S.; O’Keefe, J. Rearing on hind legs, environmental novelty, and the hippocampal formation. Rev. Neurosci. 2006, 17, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Walsh, R.N.; Cummins, R.A. The open-field test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, I.; Hatem, S.M.; Montoya, P. Abnormal Pressure Pain, Touch Sensitivity, Proprioception, and Manual Dexterity in Children with Autism Spectrum Disorders. Neural Plast. 2016, 2016, 1723401. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hao, S.; Zou, S.; Tu, X.; Kong, W.; Jiang, T.; Chen, J.G. Cortex-restricted deletion of Foxp1 impairs barrel formation and induces aberrant tactile responses in a mouse model of autism. Mol. Autism. 2023, 14, 34. [Google Scholar] [CrossRef]
- Auerbach, B.D.; Manohar, S.; Radziwon, K.; Salvi, R. Auditory hypersensitivity and processing deficits in a rat model of fragile X syndrome. Neurobiol. Dis. 2021, 161, 105541. [Google Scholar] [CrossRef] [PubMed]
- Deemyad, T.; Puig, S.; Papale, A.E.; Qi, H.; LaRocca, G.M.; Aravind, D.; LaNoce, E.; Urban, N.N. Lateralized Decrease of Parvalbumin+ Cells in the Somatosensory Cortex of ASD Models Is Correlated with Unilateral Tactile Hypersensitivity. Cereb. Cortex. 2022, 32, 554–568. [Google Scholar] [CrossRef]
- Uzunova, G.; Pallanti, S.; Hollander, E. Excitatory/inhibitory imbalance in autism spectrum disorders: Implications for interventions and therapeutics. World. J. Biol. Psychiatry 2016, 17, 174–186. [Google Scholar] [CrossRef]
- Lin, H.C.; Gean, P.W.; Wang, C.C.; Chan, Y.H.; Chen, P.S. The amygdala excitatory/inhibitory balance in a valproate-induced rat autism model. PLoS ONE 2013, 8, e55248. [Google Scholar] [CrossRef]
- Kelley, A.E. Locomotor activity and exploration. Tech. Behav. Neural Sci. 1993, 10, 499–518. [Google Scholar]
- Dixon, W.J. Efficient analysis of experimental observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–462. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Thomas, A.M.; Schwartz, M.D.; Saxe, M.D.; Kilduff, T.S. Sleep/Wake Physiology and Quantitative Electroencephalogram Analysis of the Neuroligin-3 Knockout Rat Model of Autism Spectrum Disorder. Sleep 2017, 40, 10. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, A.; Kedves, R.; Sághy, K.; Garab, D.; Zádor, F.; Lendvai, B.; Lévay, G.; Román, V. R-Baclofen Treatment Corrects Autistic-like Behavioral Deficits in the RjIbm(m):FH Fawn-Hooded Rat Strain. Pharmaceuticals 2024, 17, 939. https://doi.org/10.3390/ph17070939
Varga A, Kedves R, Sághy K, Garab D, Zádor F, Lendvai B, Lévay G, Román V. R-Baclofen Treatment Corrects Autistic-like Behavioral Deficits in the RjIbm(m):FH Fawn-Hooded Rat Strain. Pharmaceuticals. 2024; 17(7):939. https://doi.org/10.3390/ph17070939
Chicago/Turabian StyleVarga, Anita, Rita Kedves, Katalin Sághy, Dénes Garab, Ferenc Zádor, Balázs Lendvai, György Lévay, and Viktor Román. 2024. "R-Baclofen Treatment Corrects Autistic-like Behavioral Deficits in the RjIbm(m):FH Fawn-Hooded Rat Strain" Pharmaceuticals 17, no. 7: 939. https://doi.org/10.3390/ph17070939