
Chemical Composition, Antioxidant, and Cytotoxic Effects of Senna rugosa Leaf and Root Extracts on Human Leukemia Cell Lines

 ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of ELSR
2.2. Antioxidant Activity Assays
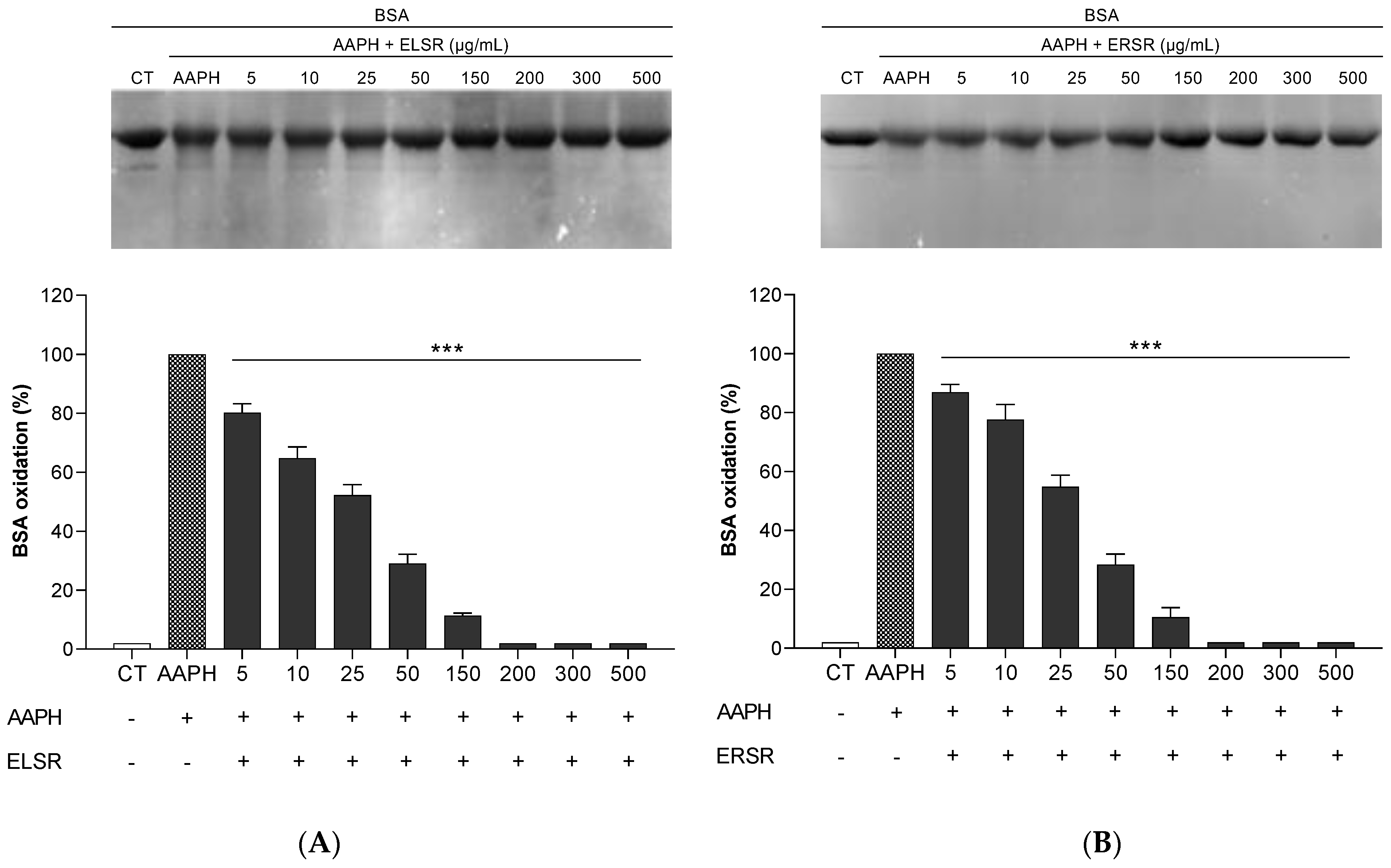
2.3. Protection of ELSR and ERSR against Oxidative Damage in Macromolecules
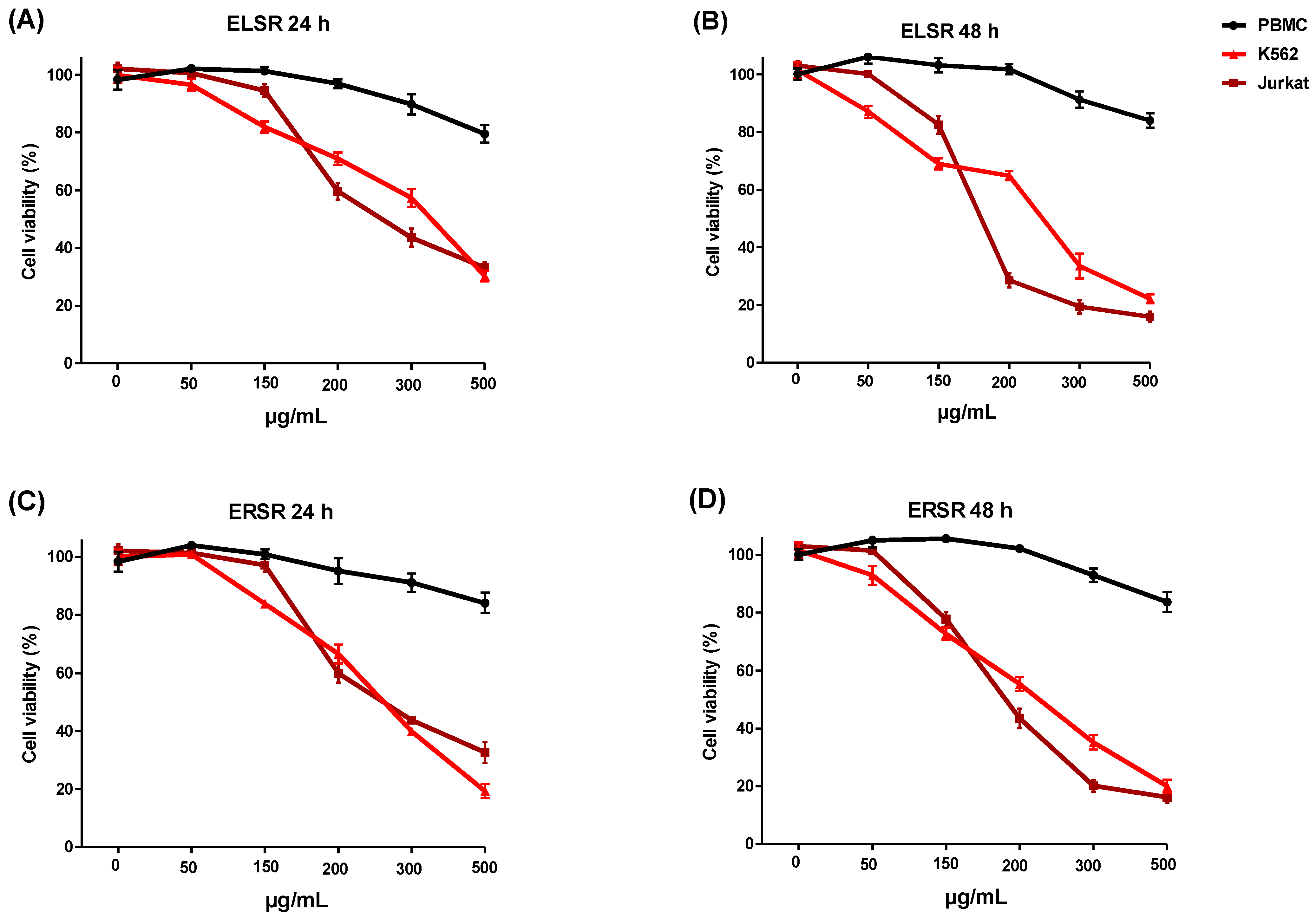
2.4. Cytotoxicity against Leukemic Strains
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
Phytochemical Analyses
4.3. Antioxidant Activity Assays
4.3.1. Direct ABTS•+ Radical Scavenging Assay
4.3.2. Direct DPPH• Radical Scavenging Assay
4.4. Protection against Oxidative Damage to the Macromolecules by ELSRs and ERSRs
4.4.1. Azo Initiator 2,2′-Azobis-(2-amidinopropane) dihydrochloride AAPH-Induced Oxidation of Proteins
4.4.2. Assay on Protection against DNA Damages (H2O2)
4.5. Cell-Based Assays
4.5.1. Cell Culture
4.5.2. Isolation of Mononuclear Cells from Human Peripheral Blood
4.5.3. Evaluation of Cytotoxicity of ELSR and ERSR
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nogueira, V.; Hay, N. Molecular Pathways: Reactive Oxygen Species Homeostasis in Cancer Cells and Implications for Cancer Therapy. Clin. Cancer Res. 2013, 19, 4309. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: Antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch. Toxicol. 2024, 98, 1323–1367. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.E.; Rivera-Del Valle, N.; Chandra, J. Redox Control of Leukemia: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2013, 18, 1349. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Ma, W.; Liu, P.; Zhou, F. Overview of thioredoxin system and targeted therapies for acute leukemia. Mitochondrion 2019, 47, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Badana, A.K.; Murali Mohan, G.; Shailender, G.; Malla, R.R. Reactive Oxygen Species: A Key Constituent in Cancer Survival. Biomark. Insights 2018, 13, 1177271918755391. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, A.; Sahu, R.P. Potential Contributions of Antioxidants to Cancer Therapy: Immunomodulation and Radiosensitization. Integr. Cancer Ther. 2018, 17, 210. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Az-Zahra, F.; Wongso, H.; Setyawati, L.U.; Novitasari, D.; Ikram, E.H.K. Molecular Mechanism of Natural Food Antioxidants to Regulate ROS in Treating Cancer: A Review. Antioxidants 2024, 13, 207. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Qureshi, A.; Hall, G. Leukaemias: A review. Paediatr. Child Health 2017, 27, 489–494. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Ma, S. Racial Differences in Four Leukemia Subtypes: Comprehensive Descriptive Epidemiology. Sci. Rep. 2018, 8, 548. [Google Scholar] [CrossRef] [PubMed]
- Mwirigi, A.; Dillon, R.; Raj, K. Acute leukaemia. Medicine 2017, 45, 280–286. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Schwartz, D.M.; Villarino, A.V.; Gadina, M.; McInnes, I.B.; Laurence, A. The JAK-STAT Pathway: Impact on Human Disease and Therapeutic Intervention. Annu. Rev. Med. 2015, 66, 311. [Google Scholar] [CrossRef] [PubMed]
- Gioia, L.; Siddique, A.; Head, S.R.; Salomon, D.R.; Su, A.I. A genome-wide survey of mutations in the Jurkat cell line. BMC Genom. 2018, 19, 334. [Google Scholar] [CrossRef] [PubMed]
- Rose-Inman, H.; Kuehl, D. Acute Leukemia. Hematol. Oncol. Clin. N. Am. 2017, 31, 1011–1028. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Estimated Number of New Cases in 2020, Worldwide, Both Sexes, All Ages. Available online: https://gco.iarc.fr/today/online-analysis-table?v=2020&mode=cancer&mode_population=countries&population=900&populations=900&key=asr&sex=0&cancer=39&type=0&statistic=5&prevalence=0&population_group=0&ages_group%5B%5D=0&ages_group%5B%5D=17&group_cancer=1&in (accessed on 10 June 2022).
- De Giffoni De Carvalho, J.T.; Da Silva Baldivia, D.; Leite, D.F.; De Araújo, L.C.A.; De Toledo Espindola, P.P.; Antunes, K.A.; Rocha, P.S.; De Picoli Souza, K.; Dos Santos, E.L. Medicinal Plants from Brazilian Cerrado: Antioxidant and Anticancer Potential and Protection against Chemotherapy Toxicity. Oxidative Med. Cell. Longev. 2019, 2019, 3685264. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.T.H.; Campos, J.F.; Damião, M.J.; Torquato, H.F.V.; Paredes-Gamero, E.J.; Carollo, C.A.; Rodrigues, E.G.; de Picoli Souza, K.; dos Santos, E.L. Ethanolic Extract of Senna velutina Roots: Chemical Composition, In Vitro and In Vivo Antitumor Effects, and B16F10-Nex2 Melanoma Cell Death Mechanisms. Oxidative Med. Cell. Longev. 2019, 2019, 5719483. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.T.H.; Leite, D.F.; da Silva Baldivia, D.; dos Santos, H.F.; Balogun, S.O.; da Silva, D.B.; Carollo, C.A.; de Picoli Souza, K.; dos Santos, E.L. Structural Characterization and Anticancer Activity of a New Anthraquinone from Senna velutina (Fabaceae). Pharmaceuticals 2023, 16, 951. [Google Scholar] [CrossRef] [PubMed]
- Oladeji, O.S.; Adelowo, F.E.; Oluyori, A.P. The genus Senna (Fabaceae): A review on its traditional uses, botany, phytochemistry, pharmacology and toxicology. S. Afr. J. Bot. 2021, 138, 1–32. [Google Scholar] [CrossRef]
- Baldivia, D.d.S.; Leite, D.F.; de Castro, D.T.H.; Campos, J.F.; Dos Santos, U.P.; Paredes-Gamero, E.J.; Carollo, C.A.; Silva, D.B.; Souza, K.d.P.; Dos Santos, E.L. Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens. Int. J. Mol. Sci. 2018, 19, 2432. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.F.; De Castro, D.T.H.; Damiaõ, M.J.; Vieira Torquato, H.F.; Paredes-Gamero, E.J.; Carollo, C.A.; Estevinho, L.M.; De Picoli Souza, K.; Santos, E.L. Dos The Chemical Profile of Senna velutina Leaves and Their Antioxidant and Cytotoxic Effects. Oxidative Med. Cell. Longev. 2016, 2016, 8405957. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.F.; Espindola, P.P.d.T.; Torquato, H.F.V.; Vital, W.D.; Justo, G.Z.; Silva, D.B.; Carollo, C.A.; de Picoli Souza, K.; Paredes-Gamero, E.J.; dos Santos, E.L. Leaf and Root Extracts from Campomanesia adamantium (Myrtaceae) Promote Apoptotic Death of Leukemic Cells via Activation of Intracellular Calcium and Caspase-3. Front. Pharmacol. 2017, 8, 466. [Google Scholar] [CrossRef]
- Casagrande, J.C.; Macorini, L.F.B.; Antunes, K.A.; Dos Santos, U.P.; Campos, J.F.; Dias-Júnior, N.M.; Sangalli, A.; Cardoso, C.A.L.; Do Carmo Vieira, M.; Rabelo, L.A.; et al. Antioxidant and cytotoxic activity of hydroethanolic extract from Jacaranda decurrens leaves. PLoS ONE 2014, 9, e112748. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H. Cancer Preventive Activities of Tea Catechins. Molecules 2016, 21, 1679. [Google Scholar] [CrossRef]
- Farias, D.F.; Cavalheiro, M.G.; Viana, M.P.; Queiroz, V.A.; Rocha-Bezerra, L.C.B.; Vasconcelos, I.M.; Morais, S.M.; Carvalho, A.F.U. Water extracts of Brazilian leguminous seeds as rich sources of larvicidal compounds against Aedes aegypti L. An. Acad. Bras. Cienc. 2010, 82, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, L.M.; De Paula-Souza, J.; Andrade, A.; Brandão, M.G.L. Plants from the Brazilian Traditional Medicine: Species from the books of the Polish physician Piotr Czerniewicz (Pedro Luiz Napoleão Chernoviz, 1812–1881). Rev. Bras. Farmacogn. 2017, 27, 388–400. [Google Scholar] [CrossRef]
- Rodrigues, V.E.G.; Carvalho, D.A. de Levantamento etnobotânico de plantas medicinais no domínio do Cerrado na região do Alto Rio Grande—Minas Gerais. Ciência Agrotecnol. 2001, 25, 102–123. [Google Scholar]
- Cunha, L.F.; Costa, C.M.; Barroso, P.R.; Kato, K.C.; Oliveira, F.D.; Victor, C.; Filho, M.; Fernanda, C.; Grael, F.; Gregório, L.E.; et al. Pharmacognosy Phytochemical screening and biological assays of ethanolic leaf extract of Senna rugosa widely used in the popular medicine although not yet adequately investigated as to its phytoconstituents and Abstract Resumo Senna rugosa (Fabaceae) é. Rodriguésia 2020, 7, 2–15. [Google Scholar]
- Silva, J.G.A.; Silva, A.A.; Coutinho, I.D.; Pessoa, C.O.; Cavalheiro, A.J.; Silva, M.G.V. Chemical Profile and Cytotoxic Activity of Leaf Extracts from Senna spp. from Northeast of Brazil. J. Braz. Chem. Soc. 2016, 27, 1872–1880. [Google Scholar]
- Maia, I.R.D.O.; Trevisan, M.T.S.; Silva, M.G.D.V.; Breuer, A.; Owen, R.W. Characterization and quantitation of polyphenolic compounds in senna macranthera var pudibunda from the Northeast of Brazil. Nat. Prod. Commun. 2019, 14, 1934578X19851704. [Google Scholar] [CrossRef]
- Wang, S.; Feng, K.; Han, L.; Fang, X.; Zhang, Y.; Yu, H.; Pang, X. Glycosidic compounds from Cassia obtusifolia seeds and their inhibitory effects on OATs, OCTs and OATPs. Phytochem. Lett. 2019, 32, 105–109. [Google Scholar] [CrossRef]
- Negri, A.; Naponelli, V.; Rizzi, F.; Bettuzzi, S. Molecular targets of epigallocatechin—Gallate (EGCG): A special focus on signal transduction and cancer. Nutrients 2018, 10, 1936. [Google Scholar] [CrossRef] [PubMed]
- Almatrood, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydh, F.A.; Alsahl, M.A.; Rahmani, A.H. Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 2020, 25, 3146. [Google Scholar] [CrossRef] [PubMed]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C.E.; Farzaei, M.H.; Bishayee, A. Targeting Multiple Signaling Pathways in Cancer: The Rutin Therapeutic Approach. Cancers 2020, 12, 2276. [Google Scholar] [CrossRef]
- Yadav, P.; Parshad, B.; Manchanda, P.; Sharma, S.K. Chromones and their derivatives as radical scavengers: A remedy for cell impairment. Curr. Top. Med. Chem. 2014, 14, 2552–2575. [Google Scholar] [CrossRef]
- Duan, Y.; Jiang, Y.Y.; Guo, F.X.; Chen, L.X.; Xu, L.L.; Zhang, W.; Liu, B. The antitumor activity of naturally occurring chromones: A review. Fitoterapia 2019, 135, 114–129. [Google Scholar] [CrossRef]
- Amen, Y.; Elsbaey, M.; Othman, A.; Sallam, M.; Shimizu, K. Naturally occurring chromone glycosides: Sources, bioactivities, and spectroscopic features. Molecules 2021, 26, 7646. [Google Scholar] [CrossRef]
- Qiu, H.; Wang, P.; Lin, H.; Tang, C.; Zhu, H.; Yang, Y. Naphthoquinones: A continuing source for discovery of therapeutic antineoplastic agents. Chem. Biol. Drug Des. 2018, 91, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Wellington, K.W. Understanding cancer and the anticancer activities of naphthoquinones—A review. RSC Adv. 2015, 5, 20309–20338. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F.; Farhan, M.; Jafri, A. Elucidation of Rutin’s Role in Inducing Caspase-Dependent Apoptosis via HPV-E6 and E7 down-Regulation in Cervical Cancer HeLa Cells. Biosci. Rep. 2021, 41, BSR20210670. [Google Scholar] [CrossRef] [PubMed]
- Raina, R.; Pramodh, S.; Rais, N.; Haque, S.; Shafarin, J.; Bajbouj, K.; Hamad, M.; Hussain, A. Luteolin Inhibits Proliferation, Triggers Apoptosis and Modulates Akt/MTOR and MAP Kinase Pathways in HeLa Cells. Oncol. Lett. 2021, 21, 192. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.C.; Chang, C.H.; Liao, H.R.; Cheng, M.J.; Wu, M.D.; Fu, S.L.; Chen, J.J. Rare Chromone Derivatives from the Marine-Derived Penicillium citrinum with Anti-Cancer and Anti-Inflammatory Activities. Mar. Drugs 2021, 19, 25. [Google Scholar] [CrossRef]
- Kaur, S.; Kumar, A.; Thakur, S.; Kumar, K.; Sharma, R.; Sharma, A.; Singh, P.; Sharma, U.; Kumar, S.; Landi, M.; et al. Antioxidant, Antiproliferative and Apoptosis-Inducing Efficacy of Fractions from Cassia fistula L. Leaves. Antioxidants 2020, 9, 173. [Google Scholar] [CrossRef]
- Minker, C.; Duban, L.; Karas, D.; Järvinen, P.; Lobstein, A.; Muller, C.D. Impact of Procyanidins from Different Berries on Caspase 8 Activation in Colon Cancer. Oxidative Med. Cell. Longev. 2015, 2015, 154164. [Google Scholar] [CrossRef]
- Klaunig, J.E. Oxidative Stress and Cancer. Curr. Pharm. Des. 2019, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Novel aspects of oxidative stress-associated carcinogenesis. Antioxid. Redox Signal. 2006, 8, 1373–1377. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- George, S.; Abrahamse, H. Redox potential of antioxidants in cancer progression and prevention. Antioxidants 2020, 9, 1156. [Google Scholar] [CrossRef]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K. Reactive oxygen species (ROS) and cancer: Role of antioxidative nutraceuticals. Cancer Lett. 2017, 387, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Kaurinovic, B.; Vastag, D. Flavonoids and Phenolic Acids as Potential Natural Antioxidants; IntechOpen: London, UK, 2019. [Google Scholar]
- Nijveldt, R.J.; Van Nood, E.; Van Hoorn, D.E.C.; Boelens, P.G.; Van Norren, K.; Van Leeuwen, P.A.M. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Farha, A.K.; Gan, R.Y.; Li, H.B.; Wu, D.T.; Atanasov, A.G.; Gul, K.; Zhang, J.R.; Yang, Q.Q.; Corke, H. The anticancer potential of the dietary polyphenol rutin: Current status, challenges, and perspectives. Crit. Rev. Food Sci. Nutr. 2022, 62, 832–859. [Google Scholar] [CrossRef] [PubMed]
- Lucas, D.M.; Still, P.C.; Pérez, L.B.; Grever, M.R.; Kinghorn, A.D. Potential of Plant-Derived Natural Products in the Treatment of Leukemia and Lymphoma. Curr. Drug Targets 2010, 11, 812. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, C.M.; Cruz-Rodriguez, N.; Quijano, S.; Fiorentino, S. Plant-derived extracts and metabolic modulation in leukemia: A promising approach to overcome treatment resistance. Front. Mol. Biosci. 2023, 10, 1229760. [Google Scholar] [CrossRef] [PubMed]
- Senapati, S.; Mahanta, A.K.; Kumar, S.; Maiti, P. Controlled drug delivery vehicles for cancer treatment and their performance. Signal Transduct. Target. Ther. 2018, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Mócsai, A.; Ruland, J.; Tybulewicz, V.L.J. The SYK tyrosine kinase: A crucial player in diverse biological functions. Nat. Rev. Immunol. 2010, 10, 387. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Gupta, D.; Gupta, R.K. Bioprotective properties of Dragon’s blood resin: In vitro evaluation of antioxidant activity and antimicrobial activity. BMC Complement. Altern. Med. 2011, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Lopez-Burillo, S.; Reiter, R.J. Oxidative damage to catalase induced by peroxyl radicals: Functional protection by melatonin and other antioxidants. Free Radic. Res. 2003, 37, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chattopadhyay, S. DNA damage protecting activity and antioxidant potential of pudina extract. Food Chem. 2007, 100, 1377–1384. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ELSR | |||||||
|---|---|---|---|---|---|---|---|
| Compound | Classes | MS/MS | [M + H]+ | Formula | UV | Time (min) | Peak |
| Catechin | Flavan-3-ol | - | 291.087 | C15H14O6 | 280 | 10.6 | L-1 |
| Rutin | Flavonol | 465 (C21H21O12), 449 (C21H21O11), 303 (C15H11O7) | 611.1599 | C27H30O16 | 256/353 | 19.4 | L-2 |
| 3,3′,5,5′-Tetrahydroxy-4-methoxystilbene | Stilbene | 225 (C14H9O3), 213 (C13H9O3), 197 (C13H9O2), 185 (C12H9O2), 157 (C11H9O) | 275.0914 | C15H14O5 | 313 | 19.8 | L-3 |
| Epiafzelechin-epiafzelechin derivative | Proanthocyanidin | 393 (C22H17O7), 285 (C16H13O5), 271 (C15H11O5), 259 (C14H11O5), 241 (C14H9O4), 147 (C9H7O2) | 547.158 | C30H26O10 | 280 | 20.1 | L-4 |
| Epiafzelechin-epiafzelechin derivative | Proanthocyanidin | 393 (C22H17O7), 271 (C15H11O5), 241 (C14H9O4) | 547.1582 | C30H26O10 | 280 | 20.3 | L-5 |
| Epiafzelechin-epiafzelechin derivative | Proanthocyanidin | 409 (C22H17O8), 269 (C16H13O4), 243 (C14H11O4), 163 (C9H7O3) | 547.1589 | C30H26O10 | 280 | 20.7 | L-6 |
| Kaempferol 3-O-rutinoside | Flavonol | 449 (C21H21O11), 287 (C15H11O6) | 595.1651 | C27H30O15 | 265/347 | 21.1 | L-7 |
| Trihydroxyflavan-epiafzelechin derivative | Proanthocyanidin | 393 (C22H17O7), 269 (C16H13O4) | 531.1644 | C30H26O9 | 280 | 22.6 | L-8 |
| Trihydroxyflavan-epiafzelechin derivative | Proanthocyanidin | 393 (C22H17O7), 269 (C16H13O4) | 531.1648 | C30H26O9 | 280 | 24.9 | L-9 |
| Trihydroxyflavan-epiafzelechin derivative | Proanthocyanidin | 393 (C22H17O7), 269 (C16H13O4), 243 (C14H11O4), 207 (C11H11O4) | 531.1648 | C30H26O9 | 280 | 25.6 | L-10 |
| Luteolin | Flavone | 241 (C14H9O4), 153 (C7H5O4) | 287.0546 | C15H10O6 | 277/345 | 25.9 | L-11 |
| Trihydroxyflavan-epiafzelechin derivative | Proanthocyanidin | - | 531.1651 | C30H26O9 | 280 | 26.3 | L-12 |
| 3-metoxyluteolin | - | 317.0662 | C16H12O7 | 279/350 | 27.2 | L-13 | |
| Trimeric procyanidins | Proanthocyanidin | - | 787.236 | C45H38O13 | 280 | 29.2–30.1 | L-14 |
| Trimeric procyanidins | Proanthocyanidin | - | 771.2412 | C45H38O12 | 280 | 30.2–30.9 | L-15 |
| Dimeric tetrahydroanthracene derivative | Tetrahydroanthracene | 533 (C32H37O7), 461 (C29H33O5) | 593.275 | C34H40O9 | 406 | 43.2 | L-16 |
| ERSR | |||||||
| Compound | Classes | MS/MS | [M + H]+ | Formula | UV | Time (min) | Peak |
| unknown | - | 489 (C24H25O11), 423 (C20H23O10), 345 (C18H17O7), 291 (C15H15O6) | 561.1791 | C24H32O15 | 253/298 | 9.8 | R-1 |
| Obtusichromoneside derivative | Chromone | 323 (C16H19O7), 293 (C15H17O6), 235 (C12H11O5), 205 (C11H9O4) | 443.1542 | C20H26O11 | 254/299 | 12.7 | R-2 |
| Obtusichromoneside derivative | Chromone | 371 (C20H19O7), 259 (C14H11O5), 235 (C12H11O5), 205 (C11H9O4) | 443.1548 | C20H26O11 | 252/295 | 13 | R-3 |
| 2-Methoxystypandrone | Naphthoquinone | 197 (C13H9O2), 169 (C12H9O) | 261.0757 | C14H12O5 | 320 | 14.9 | R-4 |
| Tetrahydroxy-methoxy stilbene derivative | Stilbene | 213 (C13H9O3), 197 (C13H9O2), 169 (C12H9O) | 275.091 | C15H14O5 | 313 | 16.2 | R-5 |
| 3,3′,5,5′-Tetrahydroxy-4-methoxystilbene | Stilbene | 225 (C14H9O3), 213 (C13H9O3), 197 (C13H9O2), 185 (C12H9O2), 157 (C11H9O) | 275.0914 | C15H14O5 | 313 | 19.8 | R-6 |
| chromone deriative | Chromone | 271 (C15H11O5), 243 (C14H11O4), 203 (C11H7O4) | 303.0867 | C16H14O6 | 280/334 | 22 | R-7 |
| Putative norrubrofusarin gentiobioside | Naphthopyrone | 259 (C14H11O5) | 553.1554 | C25H28O14 | 278/326/400 | 22.3 | R-8 |
| Putative hexahydroxy Flavanonol pentosyl-hexosyl | Flavanone | 273 (C15H13O5) | 567.1699 | C26H30O14 | 279/312/366 | 22.9 | R-9 |
| Rubrofusarin gentiobioside | Naphthopyrone | 273 (C15H13O5) | 597.18 | C27H32O15 | 277/324/399 | 24.2 | R-10 |
| Putative hexahydroxy Flavanonol sulfate | Flavanone | 273 (C15H13O5), 230 (C13H10O4) | 353.0314 | C15H12O8S | 276/310/369 | 25.1 | R-11 |
| Cassiaside B | Naphthopyrone | 273 (C15H13O5) | 567.1709 | C26H30O14 | 277/324/395 | 26.4 | R-12 |
| Putative hexahydroxy-methoxy Flavanonol pentosyl-hexosyl | Flavanone | 449 (C22H25O10), 287 (C16H15O5) | 581.1864 | C27H32O14 | 285/320/379 | 26.7 | R-13 |
| Rubrofusarin-O-glucopyranoside | Naphthopyrone | 273 (C15H13O5) | 435.1286 | C21H22O10 | 277/324/401 | 27.5 | R-14 |
| unknown | - | 449 (C22H25O10), 287 (C16H15O5) | 611.1963 | C28H34O15 | 279/330/413 | 28.6 | R-15 |
| Putative hexahydroxy-methoxy Flavanonol sulfate | Flavanone | 287 (C16H15O5) | 367.0472 | C16H14O8S | 285/321/377 | 28.9 | R-16 |
| unknown | - | 449 (C22H25O10), 419 (C21H23O9), 287 (C16H15O5) | 581.1861 | C27H32O14 | 258/278/331/411 | 30.1 | R-17 |
| unknown | - | 287 (C16H15O5) | 449.1433 | C22H24O10 | 279/330/411 | 30.8 | R-18 |
| Rubrofusarin | Naphthopyrone | 230 (C13H10O4) | 273.0758 | C15H12O5 | 277/325/402 | 35.7 | R-19 |
| unknown | - | 272 (C15H12O5), 254 (C15H10O4), 244 (C14H12O4), 226 (C15H10O3), 198 (C13H10O2) | 287.0917 | C16H15O5 | 281/337/425 | 37.7 | R-20 |
| unknown | - | 561 (C34H41O7) | 621.308 | C36H44O9 | 410 | 43.4 | R-21 |
| unknown | - | 561 (C34H41O7) | 621.3077 | C36H44O9 | 410 | 44.5 | R-22 |
| Samples | ABTS•+ | DPPH• | ||||
|---|---|---|---|---|---|---|
| IC50 (µg/mL) | Maximum Activity (%) | (µg/mL) | IC50 (µg/mL) | Maximum Activity (%) | (µg/mL) | |
| AA | 1.32 ± 0.04 | 99.28 ± 0.16 | 5 | 2.35 ± 0.34 | 94.88 ± 0.16 | 10 |
| BHT | 6.08 ± 0.77 | 98.98 ± 0.24 | 50 | 71.86 ± 2.32 | 93.16 ± 0.23 | 500 |
| ELSR | 4.86 ± 0.51 | 98.15 ± 0.53 | 50 | 19.98 ± 1.96 | 95.24 ± 0.20 | 100 |
| ERSR | 8.33 ± 0.90 | 98.40 ± 0.33 | 25 | 13.37 ± 1.05 | 94.98 ± 0.27 | 100 |
| Cell Line | ELSR IC50 (μg/mL) | ERSR IC50 (μg/mL) | ||
|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | |
| PBMC | ND | ND | ND | ND |
| K562 | 345.01 ± 2.53 | 242.54 ± 2.38 | 257.49 ± 2.41 | 223.00 ± 2.34 |
| Jurkat | 255.33 ± 2.40 | 171.45 ± 2.25 | 256.65 ± 2.40 | 189.30 ± 2.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, C.M.; Baldivia, D.d.S.; de Castro, D.T.H.; Carvalho, J.T.d.G.; Oliveira, A.S.; da Rocha, P.d.S.; Campos, J.F.; Balogun, S.O.; de Oliveira, C.F.R.; da Silva, D.B.; et al. Chemical Composition, Antioxidant, and Cytotoxic Effects of Senna rugosa Leaf and Root Extracts on Human Leukemia Cell Lines. Pharmaceuticals 2024, 17, 974. https://doi.org/10.3390/ph17080974
dos Santos CM, Baldivia DdS, de Castro DTH, Carvalho JTdG, Oliveira AS, da Rocha PdS, Campos JF, Balogun SO, de Oliveira CFR, da Silva DB, et al. Chemical Composition, Antioxidant, and Cytotoxic Effects of Senna rugosa Leaf and Root Extracts on Human Leukemia Cell Lines. Pharmaceuticals. 2024; 17(8):974. https://doi.org/10.3390/ph17080974
Chicago/Turabian Styledos Santos, Cintia Miranda, Debora da Silva Baldivia, David Tsuyoshi Hiramatsu de Castro, José Tarciso de Giffoni Carvalho, Alex Santos Oliveira, Paola dos Santos da Rocha, Jaqueline Ferreira Campos, Sikiru Olaitan Balogun, Caio Fernando Ramalho de Oliveira, Denise Brentan da Silva, and et al. 2024. "Chemical Composition, Antioxidant, and Cytotoxic Effects of Senna rugosa Leaf and Root Extracts on Human Leukemia Cell Lines" Pharmaceuticals 17, no. 8: 974. https://doi.org/10.3390/ph17080974