Abstract
Background: Gentamicin (GM) administration is associated with decreased metabolism, increased oxidative stress, and induction of nephrotoxicity. Sambucus nigra L., containing flavonoids, anthocyanins, and phytosterols, possesses antioxidant and anti-inflammatory potential. Objectives: The present study aimed to investigate the nephroprotective and anti-inflammatory potential of lyophilized Sambucus nigra fruit extract (S. nigra extract) to reduce acute oxidative stress and residual toxicity of GM in a 7-day experimental model in Balb/c rodents. Methods: The S. nigra extract was lyophilized (300 rpm; 10 min; −45 °C) to improve pharmacological properties. Balb/c mice were divided into four (n = 6) groups: controls; S. nigra extract per os (120 mg kg−1 day−1 bw); GM (200 mg kg−1 day−1 bw) (4); and GM + S. nigra therapy. The activities of antioxidant and renal enzymes, cytokines, and levels of oxidative stress biomarkers—Hydroxiproline, CysC, GST, KIM-1, PGC-1α, MDA, GSPx—were analyzed by ELISA tests. The ROS and RNS levels, as well as 5-MSL-protein oxidation, were measured by EPR spectroscopy. Results: The antioxidant-protective effect of S. nigra extract (120 mg kg−1) was demonstrated by reduced MDA, ROS, and RNS and increased activation of endogenous enzymes. Furthermore, S. nigra extract significantly reduced the expression of IL-1β, IL-6, IL-10, TNF-α, IFN-γ, and KIM-1 and regulated collagen/protein (PGC-1α and albumin) deposition in renal tissues. Conclusions: Histological evaluation confirmed that S. nigra (120 mg kg−1) attenuated renal dysfunction and structural damage by modulating oxidative stress and acute inflammation and could be used as an anti-fibrotic alternative in GM nephrotoxicity.
1. Introduction
Gentamicin (GM), as a broad-spectrum aminoglycoside antibiotic, has marked antibiotic activity against aerobic/Gram-negative bacterial infections. Clinically, GM is used in the systemic treatment of septicemia, nosocomial infections of the respiratory tract, urinary tract, and intra-abdominal infections [1]. Despite its application, GM is toxic due to reduced metabolism and difficulty in excretion with normal renal function [1,2]. The cellular toxicity pathways by which GM treatment leads to cellular destruction is concentration in the Golgi complex, followed by concentrations in the endosomal and lysosomal vacuoles of the proximal tubular cells of the kidney, leading to inflammatory and vascular responses, and acute tubular necrosis [1,2]. Different hypotheses indicate that such toxicity appears to be induced by abnormal vesicle fusion, reduced protein synthesis, and increased mitochondrial toxicity, followed by an increased imbalance of aerobic oxidative stress (OS) [3]. Directly or indirectly, the cytoplasmic GM accumulation leads to OS induction, followed by mitochondrial activity reduction and apoptotic activation [3]. GM therapy causes OS by increasing the production of reactive oxygen and nitrogen species (ROS/RNS), inflammation, and fibrosis [3,4]. The short-lived ROS (superoxide ion (•O2−), hydrogen peroxide (H2O2)), RNS (nitric oxide (•NO), peroxynitrite (ONOO−)), and the oxidized lipids as redox signaling agents generated under the cytokines and enzymes control (nitric oxide synthase, lipoxygenases and cyclooxygenases) [4] are directly involved in the metabolic regulation and adaptation to xenobiotic stress [5]. Afterward, in healthy cells, the redox balance is rapidly reduced by enzymatic and non-enzymatic systems (glutathione (GSH), melatonin, uric acid, bilirubin, and vitamin C) by natural ferro-statins, such as vitamin E, and by plant polyphenols [6,7,8]. The inability of the intracellular antioxidants to deal with oxidative disturbances leads to free radical’s induction and oxidative cascade signaling [7,8] and is involved in renal, cardiovascular, hematological, and inflammatory pathogenesis [8,9,10,11,12]. Moreover, GM therapy reduces enzyme efficacy, impairs lipid peroxidation [10], and induces renal genotoxicity by incensement of ROS/RNS [10].
Overall, plant compounds with antioxidant, anti-inflammatory, and radical-scavenging properties play a significant protective role in direct renal protection and the ability to ameliorate GM-induced nephrotoxicity [13,14,15]. Recently, there has been much interest in investigating the effects of plants containing polyphenols and quercetin-like flavonoids as protectors of inflammatory activation. The polyphenols and flavonoids reduce ROS and RNS production as controlled oxidative stress changes in acute kidney injury [16,17,18,19].
Sambucus nigra (S. nigra, black elderberry) is an active plant with antioxidant, antibacterial, and antitumor properties [20,21,22]. Traditionally, in the Balkans, the most popular is the usage of S. nigra fruits, with high content of flavonoids (rutin, quercetin-3-O-rutinoside, kaempferol-3-O-rutinoside), anthocyanins, phytosterols, triterpenes, tannins, glycosides, p-coumaric acid, and lectins, which determine the anti-inflammatory and immunomodulatory promoting activities [23,24,25,26]. The antioxidant properties of S. nigra extract have been verified to primarily depend on the presence of rutin, quercetin-3-O-rutinoside, anthocyanins, and other phenolic compounds. Furthermore, the presence of flavonoids and phenolic compounds in S. nigra helps inhibit acute phase inflammation, modulating renal toxicity through increased enzymatic protection and alleviating ROS and RNS accumulation [23,24,25,26]. In addition, it has been proven that ROS regulation by S. nigra is linked to the prevention of GM-induced renal intoxication and necrosis [25,26,27,28]. Previous research has shown that rutin and epigallocatechin contending S. nigra extracts reduced lipid peroxidation [10,11,26]. Interestingly, it has been demonstrated that rutin contending S. nigra extracts ameliorate nitric oxide (NO2) and nitric radicals (•NO) and diminish pro-inflammatory cytokine signaling as tumor necrosis factor (TNF) and interleukins IL-1β and IL-6 [29]. The experimental research on cell cultures and animal models shows that S. nigra fruit extracts promoted the increased ferocytosis of apoptotic neutrophils by increasing IL-10, reduced TNF and heme oxygenase-1, i.e., modulated redox balance by stimulating the antioxidant and enzymatic production [26,29]. Recently, it has been shown that Sambucus sp. reduced OS and inflammatory stimulated preadipocytes and macrophages by cyto-protective [26] and geno-protective properties.
Numerous components of S. nigra fruits have been identified, but their biological activity is still not fully understood. The use of dietary supplements and preparations of S. nigra fruits is accompanied by a lack of complete information on the composition, dosage, anti-inflammatory and protective activity, and compatibility with other preparations. Extracts of S. nigra fruits have not been extensively studied for their radical-modulating and anti-inflammatory effects in vivo in animal models.
Therefore, we investigated the cytoprotective and antioxidant therapeutic effects of a lyophilized S. nigra fruit extract on GM-induced nephrotoxicity in a 7-day experimental rodent model. Furthermore, we focused on the possible oxidative mechanisms of action of S. nigra-lyophilized extract against acute renal toxicity. We suggest that the ameliorating effect of the lyophilized extract of S. nigra is focused on protective action and direct regulation of ROS, NO• and •O2− concentrations, redox–homeostatic albumin imbalance, and anti-inflammatory activity in acute GM-nephrotoxicity.
2. Results
2.1. Phenolics Quantitative Evaluation and Antioxidant Capacity in S. nigra-Lyophilized Extract
The HPLC–DAD analysis showed that rutin (782.6 ± 0.68 µg g−1) was the dominant phenolic component in S. nigra-lyophilized extract, followed by epigallocatechin (314.15 ± 0.59 µg g−1), myricetin (136.06 ± 0.84 µg g−1), quercetin (37.9 ± 1.27 µg g−1), and other compounds in low concentrations (Table 1).

Table 1.
Bioactive phenolic constituents identified in lyophilized extract of S. nigra by high-performance liquid chromatography with diode array detector (HPLC–DAD; G1315B).
In addition to Table 1, Figure 1a,b presents the HPLC–DAD chromatograms of the analyzed S. nigra extract, compared to standards.
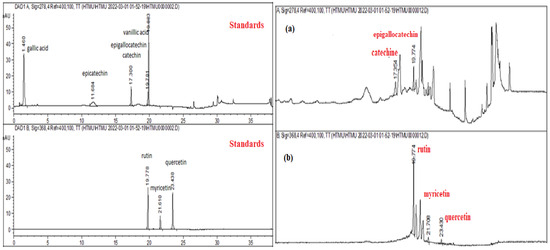
Figure 1.
HPLC–DAD chromatograms of the S. nigra extract. The reference standards ((+)-epicatechin gallate, (-)-catechin, (-) epigallocatechin, rutin, quercetin, myricetin, kaempferol, gallic, and vanillic acids) and HPLC chromatogram, where (a) catechin/epigallocatechin and (b) rutin, myricetin, and quercetin.
2.2. S. nigra-Lyophilized Extract Increased Antioxidant, Anticlastogenic, and Cytoprotective Effects
In order to assess the DPPH/R scavenging, the elevated 0.0691 µg mL−1 (87.59 ± 0.037%) of S. nigra extract confirms increased antioxidant activity, compared to other published results: 0.049 µg mL−1 (62.56 ± 1.12%) scavenging activity of S. nigra fruits methanolic extract, and 0.051 µg mL−1 (65.965 ± 0.003%) for analyzed fruit samples. Our lyophilized S. nigra extract showed a ~23% increase in scavenging potential (Figure 2).
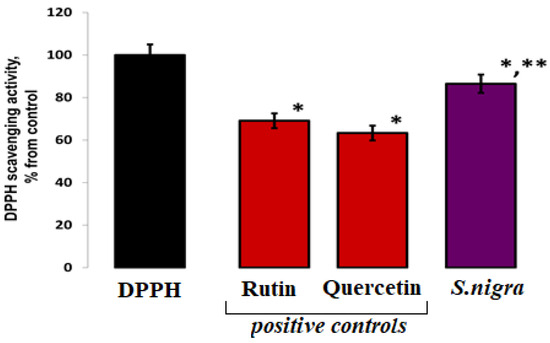
Figure 2.
DPPH (80 µM) radical scavenging capacities of S. nigra-lyophilized extract and positive controls—rutin and quercetin. DPPH-H/R generation started immediately at 23 °C, and the EPR processing was performed using WIN-EPR SimFonia 1.2/6130860 software. One-way ANOVA with Student’s t-test was used to determine statistically significant differences, at p < 0.002, between groups. The results were present in arbitrary units and recalculated as µg/DPPH radicals possible to be neutralized by 1 mL extract; * vs. DPPH; ** vs. 782.60 ppm rutin.
The chromosomal aberrations frequency shows the cytoprotective (anticlastogenic) effect of S. nigra-lyophilized extract in lymphocyte cultures (Figure 3). In the lymphocytes treated with S. nigra extract (n = 300), the aberrant cell percentage was two-fold reduced (0.7%) compared to untreated cells (2.0%) (Cramer’s V = 0.058; p < 0.2). A non-aberrant lymphocyte cultures frequency treated with S. nigra was 99.3% vs. untreated 98.0% lymphocytes. Despite the weak relationship between the factor “type of lymphocyte treatment” and aberrant cells between the two studied groups, no statistically significant difference was found (p = 0.155; p > 0.05). It should be noted that the 4 µL mL−1 S. nigra administration does not display clastogenic activity and protect against oxidative stress (Figure 3).
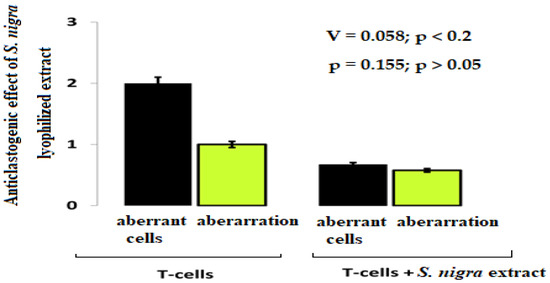
Figure 3.
The cytoprotective effect of S. nigra-lyophilized extract tracked against aberrant cells and aberrations. Lymphocyte cultures were prepared from peripheral vein blood in heparin of healthy, nonsmoking, non-drinking (female), and drug-free donors after signing written informed consent forms. The aberrance and aberrant cell degrees were processed by one-way ANOVA followed by χ2-Cramer’s V test (GraphPadPrism 6/Windows; GraphPad Software, Inc., San Diego, CA, USA). Data are presented as mean ± SD; p < 0.05 was considered statistically significant.
2.3. S. nigra Extract Attenuated GM-Induced Kidney Hypertrophy
Figure 4 shows the S. nigra effect on general physical condition, weight gain, and relative kidney weight in BALB/c mice after a 10-day GM induction. Daily GM intoxication resulted in a significant reduction in body weight (~24.63%, p < 0.05, t-test) and characteristic renal hypertrophy, as assessed by a two-fold increase in kidney weight (0.49 ± 0.02 g vs. 0.25 ± 0.01 g vs. controls). Interestingly, GM-treated mice receiving a daily protective dose of 120 mg kg−1 S. nigra showed a significant increase in body weight. S. nigra extract stimulation improved renal hypertrophy and restored kidney function compared to GM administration, observable by the kidney weight decrease (0.31 ± 0.03 g, p < 0.05).
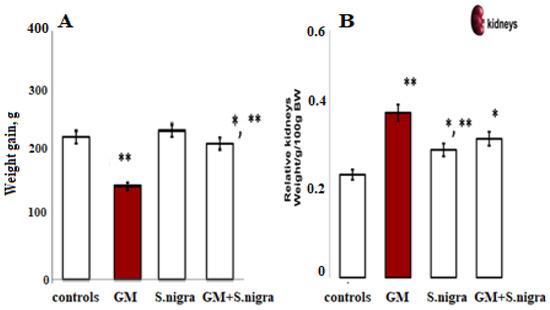
Figure 4.
Body weight increment (BW, (A)) and relative kidney weight (KW, (B)) were measured on the 10th day of this experiment; controls (standard diet), GM only, S. nigra only, GM + S. nigra combined therapy. The quantitative data were expressed as the means ± SD (n = 6). * p < 0.001 vs. controls; ** p < 0.05 vs. GM group.
2.4. S. nigra Extract Ameliorated Renal Histopathological Changes
Significantly pronounced degenerative, hypermeritic, inflammatory, and vascular changes in renal tissue were registered in the GM group. Pathomorphological renal changes were not observed in the controls, S. nigra alone, GM + S. nigra combined therapy (Table 2).
Table 2.
Protective S. nigra effect on acute GM-induced alterations in renal structures.
2.5. S. nigra Extract Ameliorated Renal Fibrosis and Reducde MCs Density and CFT
The beneficial S. nigra role in GM-induced renal fibrosis reduction by influencing MCs density was present. The kidney sections from controls, GM, and GM + S. nigra-treated mice by both toluidine-blue and Azan staining techniques were examined to determine whether GM-induced kidney fibrosis was reduced after S. nigra treatment (Table 3). Significant differences in CFT vs. controls and GM + S. nigra mice were not detected.
Table 3.
Protective S. nigra effect on GM-induced fibrosis on renal structures in controls, GM only, and GM + S. nigra combined therapy. MCs number per a microscopic field and CFT are given as mean ± SD (n = 6) in renal cortex, upper, and inner medulla of the controls, GM only, and GM + S. nigra-treated mice. Collagen fibers and bundles were thicker in GM only compared to GM + S. nigra and controls (in cortex p < 0.0001; in upper medulla p < 0.01).
Metachromatic MC localization was found in the renal corpuscles and between the proximal and distal convoluted tubules on nephron in the renal cortex. Single MCs were estimated in glomeruli and between nephron tubules in controls (Figure 5’).
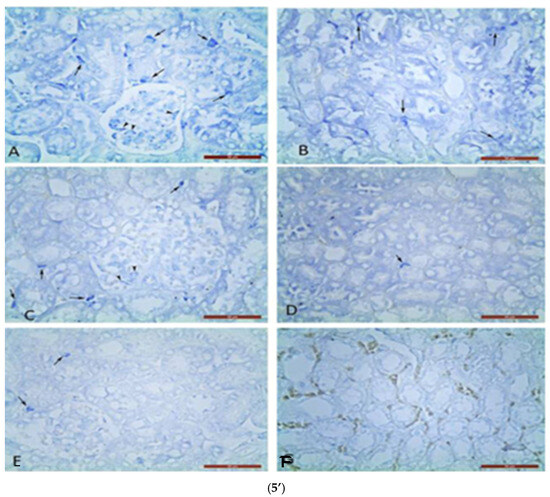
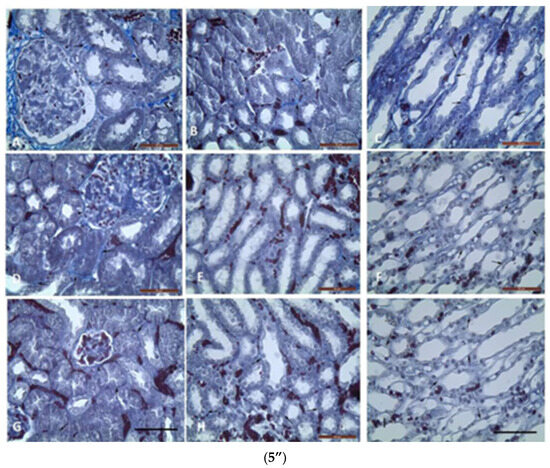
Figure 5.
Metachromatic mast cells (5’) in renal cortex (5’A,5’C,5’E) and upper medulla (5’B,5’D,5’F): (5’A,5’B) in GM treated mice; (5’C,5’D) in GM + S. nigra mice; ((5’E,5’F)-MCs absence) in controls. Arrows—MCs between nephron tubules; arrowheads—MCs in glomerulus (Bar × 50 µm). Kidney’s cortex (5”) (5”A,5”D,5”G) and upper (5”B,5”E,5”H) and inner (5”C,5”F,5”I) medulla sections obtained from mice and stained by Azan technique. Groups: (5”A,5”B,5”C) mice with GM-induced fibrosis and intensive blue staining and large collagen deposition (collagen fibers and bundles); (5”B) GM-treated group and subsequent treatment with S. nigra—thinner collagen fibers are present than in GM group; (5”C) controls with delicate (thinnest) collagen fibers comparing to GM only and GM + S. nigra therapy (Bar × 200 µm). Arrows—collagen fibers and bundles between nephron tubules; arrowheads—collagen fibers and bundles around renal corpuscle capsule. The data were processed by one-way ANOVA followed by Tukey Kramer’s test (GraphPadPrism 6 for Windows; GraphPad Software, Inc., USA), as mean ± SD; p < 0.05 was considered statistically significant.
In GM + S. nigra group, MCs densities in mentioned renal structures were not significantly higher, close to controls, and significantly lower in the GM group. In the upper medulla, the MC number in the GM group was four-fold higher compared to the controls and GM + S. nigra group. It is important to note that there was not a statistical significance in MCs density between controls and GM + S. nigra therapy. In the upper medulla of all treated groups, MCs were located between nephron tubules and collecting ducts. MCs were present in the GM group but absent in controls and GM + S. nigra therapy. Therefore, S. nigra administration caused a significant MC decrease in the renal cortex and medulla vs. GM group. Azan staining allowed for the detection of CFT and bundles in the renal cortex and medulla in order to define collagen deposition (Figure 5”).
The micro-morphometric study was used to estimate the thickness of collagen fibers and bundles in the renal cortex and medulla. In the renal cortex and upper medulla of the GM group, collagen fibers and bundles between nephron tubules were significantly thicker vs. controls and GM + S. nigra combined therapy.
2.6. S. nigra Extract Ameliorated Renal Hydroxyproline Content and Protein Oxidation
Acute GM-nephrotoxicity significantly increased HYP content compared to controls (875.18 ± 111.4 mg g−1 vs. 425.71 ± 72.9 mg g−1, t-test, p < 0.05). Figure 6A presents a protective S. nigra effect against GM renal injury. The hydroxyproline content was statistically significantly decreased in the GM + S. nigra group (875.18 ± 111.4 mg g−1 vs. 595.09 ± 88.1 mg g−1 tissue, p < 0.005). The 10 days of protection with S. nigra extract statistically insignificant reduced collagen deposition versus controls (401.22 ± 35.4 mg g−1 vs. 425.71 ± 72.9 mg g−1).
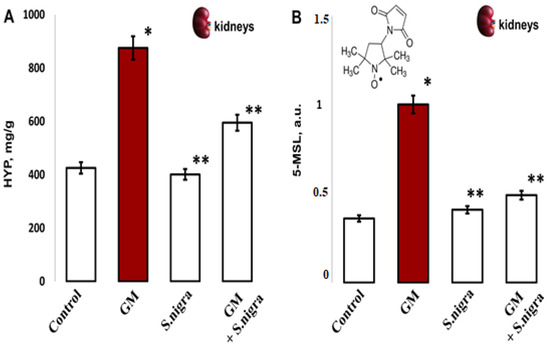
Figure 6.
The S. nigra extract effects on GM-induced nephrotoxicity and oxidative changes in (A) renal hydroxyproline content (mg/g) and (B) protein oxidation levels by 5-MSL spin–adduct formations, expressed in arbitrary units (a.u.) after tree independent EPR measurements. The results are presented as mean ± SD (n = 6). One-way ANOVA with Student’s t-test was used to determine significant differences in relation to (*) p < 0.05 vs. controls and (**) p < 0.005 vs. GM-treated mice.
In vivo, remodeled protein oxidation was determined after 5-MSL-albumin/protein conjugation (Figure 6B). Compared with controls, renal protein expression was significantly increased (0.361 ± 0.04 vs. 1.002 ± 0.52 a.u., p < 0.005, t-test) after GM intoxication. S. nigra treatment significantly reduced GM-induced kidney protein dysregulation compared to the GM group (0.409 ± 0.07 vs. 1.002 ± 0.52 a.u., p < 0.002, t-test). Notably, S. nigra administration resulted in inhibition of renal protein expression and reduced OS, with a value comparable to controls (0.41 ± 0.03 vs. 0.361 ± 0.04 a.u., p < 0.05, t-test).
2.7. S. nigra Extract Ameliorated Renal and Serum Injures
Data analysis of renal and blood concentrations of KIM-1, cystatin C, GST, gamma-GT, Cre, and urea in control, S. nigra, GM + S. nigra groups were performed to detect the S. nigra extract ability to protect against acute kidney injury (Figure 7). GM-induced tubular injury and nephrotoxicity resulted in a significant increase in renal KIM-1 expression vs. controls (6.73 ± 0.21 ng/mL vs. 2.56 ± 0.1 ng/mL, t-test, p < 0.05), the S. nigra stimulation (6.73 ± 0.17 ng/mL vs. 2.61 ± 0.09 ng/mL, t-test, p < 0.005), and the GM + S. nigra combination (6.73 ± 0.21 ng/mL vs. 3.86 ± 0.37 ng/mL, t-test, p < 0.05) (Figure 7A). No significant differences were reported between the controls and after S. nigra extract protection.

Figure 7.
The S. nigra extract effects on GM-induced nephritic and serum injury changes in (A) kidney injury molecule-1 (KIM-1) (ng/mL), (B) Cisplatin C (ng/mL), (C) glutathione-S-transferase (GST, nmol/gPr), (D) sera creatinine (ng/dL), (E) sera urea (mg/dL), and (F) sera gamma-glutamyl-transpeptidase levels (gamma-GT, ng/mL). The results are presented as mean ± SD (n = 6). One-way ANOVA with multiple comparisons using Student’s t-test was used to determine significant differences in relation to (*) p < 0.05 vs. controls, (**) p < 0.005 vs. GM-treated mice.
GM treatment vs. controls had statistically significantly increased renal Cys C expression (0.866 ± 0.07 ng/mL vs. 0.301 ± 0.016 ng/mL, t-test, p < 0.05; Figure 7B) and increased GST (892.12 ± 55.14 nmol/gPr vs. 512.3 ± 58.59 nmol/gPr, t-test, p < 0.001; Figure 7C). Also, an incensement was measured in serum Cre (1.86 ± 0.23 ng/dL vs. 0.73 ± 0.1 ng/dL, p < 0.005), urea (104.38 ± 22.35 mg/dL vs. 64.82 ± 15.35 mg/dL, p < 0.005) (Figure 7D,E), and gamma-GT (2.11 ± 0.09 ng/mL vs. 1.43 ± 0.012 ng/mL, p < 0.005, Figure 7F), confirming acute renal injuries, oxidative changes, and neutrophil cells death.
Renal enzyme activity in the GM-administrated group was statistically significantly higher versus GM + S. nigra combined therapy. The induction of Cys C (0.866 ± 0.07 ng/mL vs. 0.67 ± 0.04 ng/mL, t-test, p < 0.05, Figure 7B), GST (892.12 ± 55.14 nmol/gPr vs. 647 ± 33.59 nmol/gPr, t-test, p < 0.005, Figure 7C), sera creatinine (1.86 ± 0.23 vs. 0.84 ± 0.3, p < 0.005, Figure 7D), sera urea (104.38 ± 22.35 vs. 76.85 ± 11.44, p < 0.05; Figure 7E), and sera gamma-GT (2.11 ± 0.09 vs. 1.75 ± 0.04, p < 0.05, Figure 7F) were significantly reduced in renal tissue and sera after S. nigra-lyophilized extract protection. Significant enzymatic activity was not reported between the controls and S. nigra-stimulated animals.
2.8. S. nigra Extract Normalize Renal Enzyme Activities and Ameliorated Lipid Peroxidation
Next, we explored the effects of GM administration on the renal enzymatic activities. The renal SOD (1.291 ± 0.35 vs. 4.74 ± 0.86 IU/gHb, p < 0.005; Figure 8A), CAT (1.62 ± 0.13 vs. 3.78 ± 0.05 IU/gPr, p < 0.003; Figure 8B), GPx1 (18.9 9 ± 2.86 vs. 68.84 ± 5.39 IU/gPr, p < 0.05, Figure 8C), and GSH (21.99 ± 3.22 vs. 59.85 ± 2.376 IU/gPr, p < 0.04, Figure 8D) significantly decreased versus controls, which indicates an impairment in kidney functions. However, GM considerably increased renal MDA concentration (5.87 ± 0.9 vs. 3.09 ± 0.4 µmol/mL, p < 0.05; Figure 8E) versus controls.
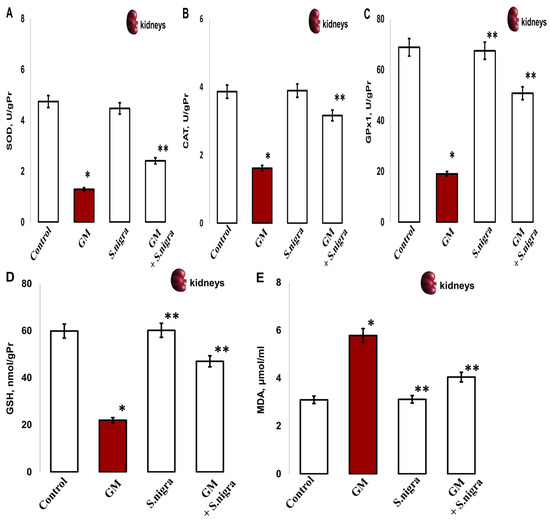
Figure 8.
The S. nigra extract effects on GM-induced nephritic antioxidant enzyme activities and lipid peroxidation changes in (A) superoxidedismutase (SOD, IU/gPr), (B) catalase (CAT, IU/gPr), (C) glutathioneperoxidase-1 (GPx1, IU/gPr), (D) reduced glutathione (GSH; IU/gPr), and (E) malondialdehyde (MDA, µmol/mL). The results are presented as mean ± SD (n = 6). One-way ANOVA with multiple comparisons using Student’s t-test was used to determine significant differences in relation to (*) p < 0.05 vs. controls, (**) p < 0.005 vs. GM-treated mice.
Conversely, the S. nigra co-administration at 120 mg kg−1 showed significant GM inhibition and substantial incensement in the enzymatic defense by OS amelioration in kidneys. Markedly, induced activities of SOD (1.291 ± 0.35 vs. 3.81 ± 0.41 IU/gHb, p < 0.05), CAT (1.62 ± 0.13 vs. 3.17 ± 0, 19 IU/gPr, p < 0.05), GPx1 (18.99 ± 2.86 vs. 50.75 ± 3.39 IU/gPr, p < 0.05), GSH (21.99 ± 3.22 vs. 46.97 ± 5.39, p < 0.001), and MDA (5.87 ± 0.9 vs. 4.041 ± 0.12 µmol/mL, p < 0.05) were registered vs. controls (Figure 8A–D).
2.9. S. nigra Extract Ameliorated ROS, NO• u •O2− Stress Levels
To confirm the protective role of S. nigra extracts against GM-induced renal oxidative stress, the redox-modulated activity was investigated. Significantly, GM treatment increased ROS (2.77 ± 0.35 vs. 0.89 ± 0.06 a.u., p < 0.005; Figure 9A), NO• (55.61 ± 5.35 vs. 14.88 ± 1.41 a.u., p < 0.002; Figure 9B), and •O2− levels (3.4 1 ± 0.72 vs. 0.97 ± 0.09 a.u., p < 0.005, Figure 9C) in the renal cortex compared to controls. The highest protection of S. nigra extract was observed in the renal cortical ROS reduction (2.77 ± 0.35 vs. 1.36 ± 0.11 a.u., p < 0.05) and in NO• (55.61 ± 5.35 vs. 33.7 ± 2.09 a.u., p < 0.002) and •O2− (3.41 ± 0.72 vs. 1.994 ± 0.12 a.u., p < 0.005) concentrations. These findings highlight the S. nigra protective effects on renal OS compared to GM-treated mice. Interestingly, S. nigra extract co-administration alleviated renal cortical oxidative stress and promoted exogenous–endogenous enzymatic defense by restoring the redox–homeostatic imbalance versus the GM-intoxicated group.
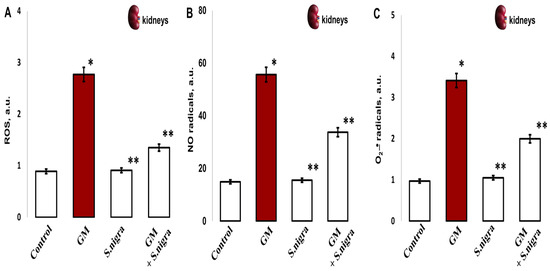
Figure 9.
The S. nigra extract effects on GM-induced nephrotoxicity and oxidative changes in (A) ROS production, (B) nitric radicals (NO•), and (C) superoxide anion radical (•O2-). The radicals were scavenged in triplicate by EPR spectroscopy using Win-EPR and Sim-Fonia software and expressed in arbitrary units (a.u.). The results are presented as mean ± SD (n = 6). One-way ANOVA with Student’s t-test was used to determine statistically significant differences in relation to * p < 0.05 vs. controls and ** p < 0.005 vs. GM-treated mice.
2.10. S. nigra Extract Protected the Kidney Against GM-Induced Acute Inflammation
Furthermore, we assessed the influence of S. nigra on GM-induced inflammation. The oxidative stress promotes renal damage and provokes fibrotic processes and loss of renal function by concentration-inactivating collagen deposition (PGC-1α) and inflammatory expression (Figure 10A–F). Significantly, GM administration reduced PGC-1α (65%, p < 0.001; Figure 10A) deposition and induced the release of inflammatory IL-1β (78.3%), IL-6 (27.4%), IL-10 (29.5%), TNF-α (55.3%), and IFN-γ (41.9%) versus controls (p < 0.005).
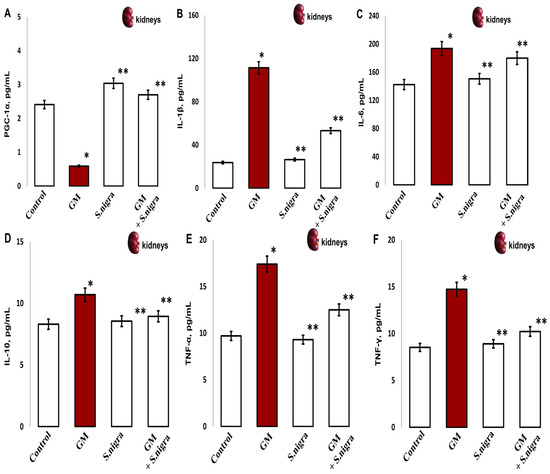
Figure 10.
The S. nigra extract effects on GM-induced nephrotoxicity and pro-inflammatory changes in (A) PGC-1 (pg mL−1), (B) IL-1β (pg mL−1), (C) IL-6 (pg mL−1), (D) IL-10 (pg mL−1), (E) TNF-α (pg mL−1), and (F) IFN-γ (pg mL−1). The results are presented as mean ± SD (n = 6). One-way ANOVA with Student’s t-test was used to determine statistically significant differences in relation to (*) p < 0.05 vs. controls and (**) p < 0.001 vs. GM-treated mice.
S. nigra extract co-administration potentially rescues renal cells and modulates cellular response by increasing PGC-1α deposition and suppressing inflammation induced by GM intoxication (p < 0.01). Furthermore, S. nigra extract stimulates renal adaptive immunity and regulates the inflammatory response, almost comparable to controls (p < 0.05).
2.11. Correlation Dependences Between Parameters for Protective Properties of S. nigra After GM Intoxication
The results of the established correlations between collagen deposition, lipid peroxidation, cytokine expression and markers of oxidative stress in relation to the S. nigra extract action of against GM nephrotoxicity are presented in Table 4.
Table 4.
Correlation dependences between parameters for protective properties of S. nigra extract after GM intoxication.
3. Discussion
Receptor-mediated endocytosis via the multi-ligand receptors megalin and tubulin promotes GM deposition in renal proximal tubules, altering lysosomal aggregation, phospholipid metabolism, and mitochondrial toxicity [3,13], thus promoting ROS, RNS accumulation, and oxidative stress [30]. Aberrant mitochondrial modulation of ROS and RNS by metabolic, hormonal, and pro-inflammatory factors, as well as endogenous–exogenous antioxidant deactivation [6,7], caused OS and organ changes [31]. GM stimulates the mitochondrial respiratory chain to accumulate H2O2 synthesis, i.e., •O2−, 1O2, HO•, OH–, NO•, and peroxy radicals. GM-induced acute renal inflammation and accumulation of abnormalities in the renal tubules and glomeruli is a consequence of the activation of inflammatory cells and physiologically impaired redox homeostasis, which predisposes to increased lipid peroxidation [7], chromosomal aberrations, and protein denaturation [31,32,33].
Biologically active flavonoids and polyphenols support acute nephron protection due to antioxidant and anti-inflammatory effects and decreased H2O2-mediated Fe2+/Fe3+ mobilization from mitochondria [7,24,32,33]. Based on these facts, we aimed to establish the protective effect of S. nigra extract against acute kidney injury and renal lesions directly caused by GM therapy. In addition, we investigated the S. nigra antioxidant regulative mechanism by maintaining the redox–homeostatic imbalance and cytoprotective effect, leading to protein oxidation remodeling and cytokine inflammation.
Ultrasonic extraction at 80 kHz improved the extraction yields of rutin (8754 μg mL−1) and epigallocatechin (3514 µg mL−1) (in the maximum amount) [12], the main flavonoids in used S. nigra extract. Rutin, epigallocatechin, and myrcetin flavonoids scavenged ROS and RNS and powerfully modulated oxidative stress changes under GM accumulation [24,31]. Previous studies have demonstrated significant amounts of polyphenolic metabolites deposited in renal tissue after a polyphenolic diet after GM-nephrotoxicity [34,35,36,37]. Rutin and epigallocatechin-rich extracts obtained from Opuntia ficus Indica, Morus alba L., and S. nigra had a nephroprotective effect [38,39,40,41]. The antioxidant capacity, recording 86.5 ± 0.037% ROS-inactivation in vitro, is probably due to the high rutin > epigallocatechin > myricetin > quercetin amounts. The relationship between phenolic content and antioxidant activity of S. nigra extract [24] was confirmed. Putri et al. [42] provided evidence that S. nigra extracts were characterized by antioxidant activity, reaching 89.25% concerning DPPH radicals responsible for the H2O2 detoxification and renal function improvement [43].
The cytoprotective efficacy test, which directly assesses the inhibition of genotoxicity, mutagenesis, and carcinogenesis by flavonoid-containing plants [44], attracted our attention. S. nigra extract (4 µL mL−1) showed a non-toxic, protective effect against lymphocyte cultures without exhibiting clastogenic activity compared to controls. Furthermore, S. nigra treatment reduced by 0.7 ± 0.02% (Cramer’s V = 0.058; p < 0.2) the deviation rate, demonstrating adequate antigenicity in lymphocytes. Genotoxic mutations induced at cytogenetic levels represent the first step toward carcinogenesis. Understanding the chromosomal mechanisms of action of S. nigra suggests a stable antimutagenic potential associated with an enhanced cytoprotective activity [44], minimizing the GM side effects. Furthermore, elderberry flavonoids modulate specific and non-specific immune responses and cytoprotective effects in various acute renal injury models [45].
Mice pretreatment with 120 mg kg−1 S. nigra extract reduced the initial degenerative, hypermeric, inflammatory, and vascular changes in renal tissue after GM intoxication [12,46]. This is consistent with reduced serum levels at Cre, gamma-GT, urea, and Cys C and significantly improved renal function, eliciting a reduction in the chain inflammatory response. S. nigra extract reduced the extracellular matrix and cell proliferation by reducing fibrotic areas in the renal cortex and medulla. Herbal antioxidants reduced GM accumulation by direct GM cytotoxicity mitigation, collagen reduction, suppression of vasoconstriction, and antioxidant–anti-inflammatory action [45]. This motivated us to conduct MCs density, taking into account their role in GM-accumulated fibrosis and to clarify the modulated S. nigra effect. S. nigra administration alone or in a GM + S. nigra combination statistically significantly ameliorates MCs and collagen deposition in the renal cortex (fourfold) and medulla and terminates renal fibrotic, probably by redox-modulative mechanism. Several studies reported that S. nigra exerts enhanced antioxidant activity even in patients with idiopathic nephrolithiasis without altering urine pH or H+ concentrations [47].
GM increases connective tissue volume and renal fibrosis by increasing TGF-β production, myofibroblast activation, and epithelial–myofibroblast transdifferentiation. Fibroblast TGF-β receptors activate both the Smad pathway and fibroblast collagen expression [48,49] and induce an increased amount of detectable MCs-TGF-β in the initial phase [50] under GM nephrotoxicity. MCs influence fibroblast activity by releasing TGF-β1 and TNF-α mediators [51,52]. The present study reports that S. nigra extract directly suppresses the initial phase of GM nephrotoxicity and renal inflammatory response, reducing TGF-β1 and TNF-α by inactivating MCs and collagen proliferation. Similarly, the C-6 ring structure of flavonoids has been reported to inhibit the expression of TGFβ/Smad and TNF-α and regulate collagen synthesis. The flavonoid ring structure suppresses the free radical overexpression [52], and matrix deposition is terminated. Together, rutin, epicatechin, and quartzetin have been shown to reduce •O2−, H2O2, OH•, and ONOO− cascade in the mitochondrial respiratory chain by enzyme reactivation and fibrotic deactivation. However, flavonoid-containing extracts of S. nigra restored TGF-β1, TNF-α, IL-6, serum urea, and Cre [53] to controls through appropriate antioxidant modulation after renal GM disorders [54].
This study focused on identifying the possible mechanisms of S. nigra actions on the transmembrane tubular protein KIM-1 responsible for proximal tubular injury [55] and 5-MSL conjugating albumin accounts for conformational changes in –SH in the albumin molecule and remodeling of protein oxidation [56]. GM treatment significantly increased renal KIM-1 expression and structurally changed -SH albumin groups, consistent with previous studies [27,55]. Interestingly, S. nigra treatment attenuated KIM-1 expression and protein oxidation to levels comparable to the controls. Acute kidney injuries are alleviated by herbal nutrients through suppressed KIM-1 [57,58] in animals. S. nigra extract exhibits a protective effect in kidney injuries [57,58] due to its high protein levels and seven essential amino acids [32,33,43,54], which probably directly inhibits the •O2−, H2O2, and HO• production and regulates albumin oxidation [43,59]. In parallel, the urinary albumin–creatinine ratio is reduced by attenuating proximal tubular injury and renal fibrosis [60,61]. Probably, S. nigra promotes the quinones production and ability to form irreversible complexes with proteins (adhesins), i.e., alters the cellular membrane potential (hyperpolarization) and modulates immune decrease in KIM-1, Cys C, and TNF-α in kidneys [61,62]. Seven-day oral exposure to S. nigra extract resulted in changes in redox homeostasis, enhanced antioxidant response to increased lipid peroxidation, and inhibition of oxidized albumin -SH groups (cysteine residues), suggesting renal mitochondrial and cytoplasmic H2O2 detoxification [60,62]. Lee et al. [63] proved that elderberry extract containing flavonoids and phytochemicals acts as an intracellular antioxidant, protecting proteins and enzymes from H2O2-induced ROS and oxidative stress.
Oxidative stress disrupts endogenous and exogenous homeostasis, which is critical in GM-induced renal pathogenesis. Free radicals promote lipid peroxidation in the kidneys, trigger pro-inflammatory pathways, and activate cytokines [10,11,24]. The fact that in the GM-treated group, the MDA and ROS products were almost two-fold increased, but SOD, CAT, and GPx1 enzymes were two times decreased is another sign of GSH depletion and nephrotoxicity activation. Our findings are consistent with previous studies [7,24,63]. In contrast, 120 mg kg−1 protection with S. nigra extract, both alone and in the GM + S. nigra combination, changes hyperpolarization and restores antioxidant homeostasis after a possible respiratory chain reversal to controls. S. nigra extract has metal-chelating properties and prevents the initiation of •OH− radicals, resulting in the termination of membrane lipids and DNA fragmentation [7,64]. Therefore, S. nigra restores SOD, CAT, and GSH enzymes by further activating GPx1, leading to H2O2 and lipid peroxide (L-OOH) detoxification. Additionally, extract limits the hydroperoxide synthesis from fatty acids, which allows for the restoration of renal anti-lipid peroxidation and GSH inhibition, even at the peroxisomal level [24,65,66]. S. nigra extract containing flavonoids activates a compensatory reaction of the body’s antioxidant defense and improves redox homeostasis by directly mitigating renal toxicity [66,67,68] and fibrotic [69,70].
Exogenous cell-permeable spin probes based on aminoxyl, nitronylnitroxide, or hydroxylamine radicals (CPTIO.K, CMH) permit ROS and RNS determination and directly reflect differences in redox status in vivo [71]. Redox-sensitive agents are reduced to the corresponding diamagnetic forms and act as catalysts in the disposal of renal NO• to the formation of an imino–nitroxide radical or the dismutation of renal •O2− to H2O2 and O2 [72,73]. In this context, GM penetrates the membrane by an oxygen-dependent transport mechanism [74] and reduces NO bioavailability in the renal cortex and medulla in harmony with upregulated KIM-1, CysC, and TNF-α activations [75]. Whereas S. nigra extract (120 mg kg−1) alleviated GM production of NO• and •O2− by reducing radicals and normalizing renal cytosolic and mitochondrial SOD and GSH deficits. The extract containing rutin, epigallocatechin, and quercetin prevents the •O2− transformation into ONOO¯ and •OH radicals, simultaneously restoring the function of NO• and NO as a signaling molecule regulating blood flow in the renal cortex. Likewise, S. nigra stimulates SOD synthesis [24,25,26] and inhibits the xanthine oxidase enzyme [9,67]. Protection (7 days) with the lyophilized extract shows significant catalysis of the •O2− and NO• into ONOO¯ radicals by reducing renal stress, followed by an antigen-specific immune response that activates the mitochondrial respiratory chain and reduces GM-induced collagen deposition and cell-mediated fibrotic [13,67]. In accordance, Olas et al. [62] demonstrated that flavonoids, isoflavonoids, and saponins promoted renal blood flow, increased glomerular filtration, and remodulated the Th2 immune response by reducing the expression of inducible NO synthase (iNOS). Therefore, the quinones production by S. nigra [73] alters the renal •O2− and •NO redox-microenvironment by promoting modifications in redox-sensitive amino acids (proline), regulating the selectivity for -SH modifications, i.e., ending renal stress deposition [74,76,77,78] (Scheme 1).
GM-induced oxidative stress acutely activates NF-κB pathways, which suppress mitochondrial biogenesis and stimulate the release of pro-inflammatory cytokines in the peripheral renal tissues. PGC-1α, as a coactivator protein, controls mitochondrial biogenesis and adaptive cellular thermogenesis, directly characterizing renal disorders [79].
The presented results show that GM presumably increased the production of IL-1β, IL-6, and IL-10, including TNF-α and IFN-γ. Early-phase inflammatory mediators significantly upregulated PGC-1α. Therefore, 7-day GM accumulation inhibited PGC-1α levels and impaired mitochondrial synthesis, which exacerbated renal stress and promoted nephrotoxicity progression [79,80]. We hypothesize that treatment with lyophilized extract of S. nigra regulates Nrf2 signaling pathways by provoking an antioxidant response (SOD, CAT, GPx1, including localized peroxidase and oxidoreductases) against acute-phase inflammatory mediators. In a compositional ratio containing polyphenols, S. nigra extract has the potential to suppress ROS, NO•, and •O2− generations directly related to lipid peroxidation, i.e., sharply reduces electron transport [81], in the mitochondrial membrane and increases the PGC-1α activity [82]. Presumably, the 7-day-lasting antioxidant compensatory response of S. nigra, containing rutin, epigallocatechin, and quercetin, stimulates Th1/Th2 response and protects renal mitochondrial dysfunction by improving renal tubular dynamics and arresting the fibrosis process [83]. Moreover, the potential of the synergistic phytochemicals rutin, epigallocatechin, quercetin, and myricetin in S. nigra extract is expressed in the complete antagonism/HO•, •O2−, NO• scavenging, inhibition of lipid peroxidation, and complete oxidative stress modulation. It has been shown that the synergistic phytochemicals functionally have a ROS/PNH scavenging capacity 100–300 times higher than the mannitol [10,84]. S. nigra extract disrupts extracellular matrix components that allow for full penetration of antibiotics, enhance their effect, and simultaneously prevent the accumulation of acute renal dysfunction.
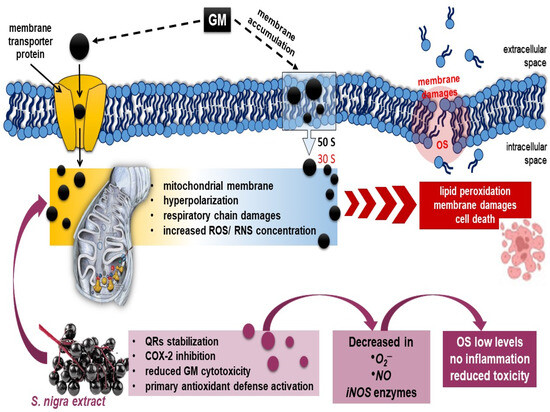
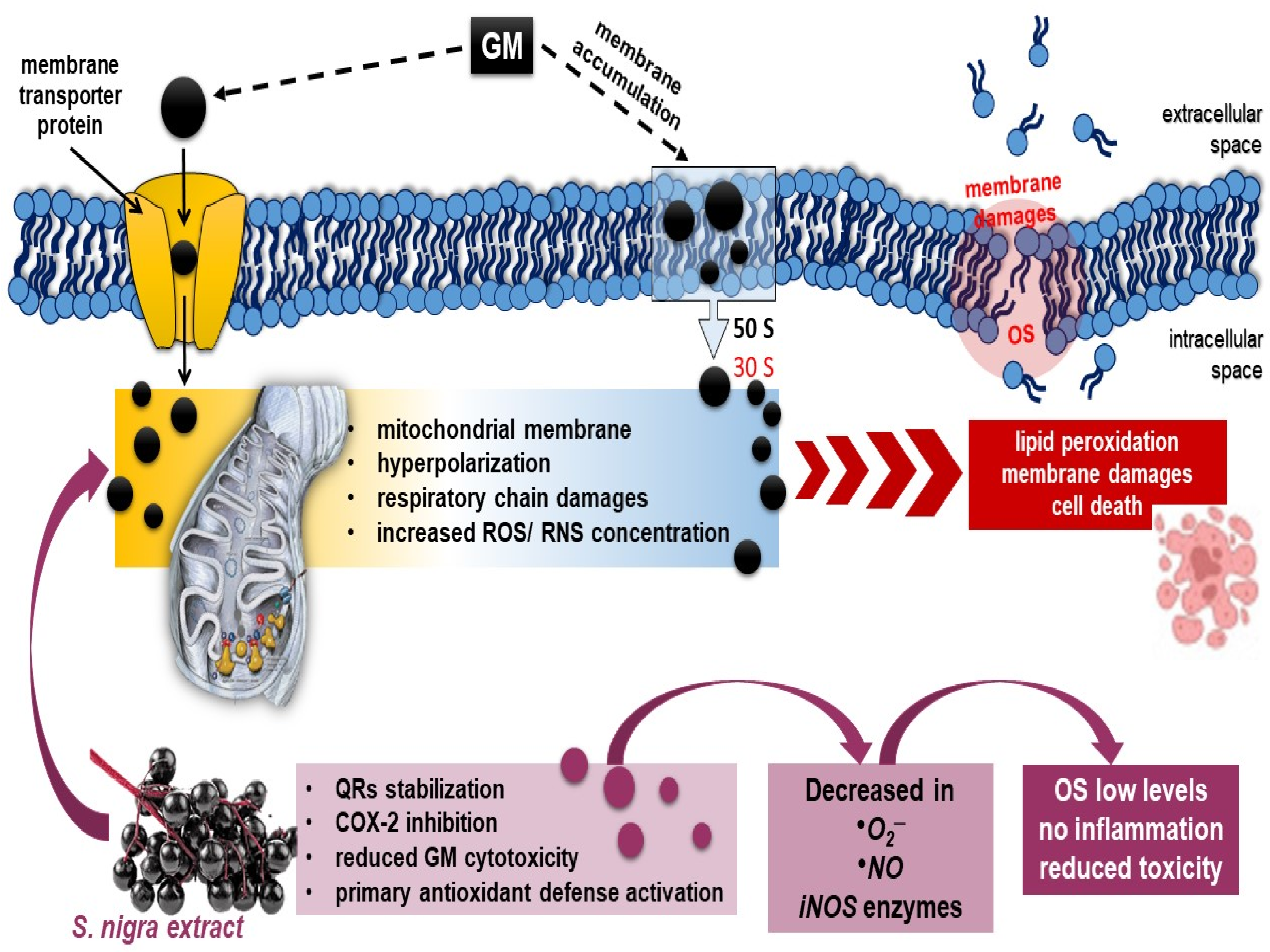
Scheme 1.
Possible mechanism of (1) GM-binding in 30S–50S ribosomal subunit and penetration by oxygen-dependent transport mechanism, hyperpolarization, and renal dysfunction [1,74]; (2) oral 7-day antioxidant compensatory response of S. nigra-lyophilized extract (containing rutin, epigallocatechin, and quercetin) in stimulation of renal mitochondrial dysfunction, improving renal tubular dynamics and halting the fibrotic process through complete ROS, NO•, and •O2− generation neutralization and OS modulation [81,82,83]. As a sequence, S. nigra-lyophilized extract increases GM uptake and decreases GM nephrotoxicity. Abbreviations: GM, gentamycin; S. nigra; OS, oxidative stress; ROS, NO•, and •O2−, free radicals.
Scheme 1.
Possible mechanism of (1) GM-binding in 30S–50S ribosomal subunit and penetration by oxygen-dependent transport mechanism, hyperpolarization, and renal dysfunction [1,74]; (2) oral 7-day antioxidant compensatory response of S. nigra-lyophilized extract (containing rutin, epigallocatechin, and quercetin) in stimulation of renal mitochondrial dysfunction, improving renal tubular dynamics and halting the fibrotic process through complete ROS, NO•, and •O2− generation neutralization and OS modulation [81,82,83]. As a sequence, S. nigra-lyophilized extract increases GM uptake and decreases GM nephrotoxicity. Abbreviations: GM, gentamycin; S. nigra; OS, oxidative stress; ROS, NO•, and •O2−, free radicals.
4. Materials and Methods
4.1. S. nigra Fruit Phytochemical Analysis and Ultrasound-Assisted Extraction
The S. nigra fruits were collected from Alino, Bulgaria, 2023 g. The juice was obtained by manual pressing and straining through sterile gauze and ultrasonically sonicated. For ultrasonic treatment (80 kHz), a bath with a piezoceramic emitter (Elmasonic P 30 H, Elma, Singen, Germany; volume 2.8 L; frequency 37–80 kHz; 22 °C) was used. The extract was filtered through cotton and a 0.8 μm nylon-membrane syringe filter (Acrodisc, Sigma-Aldrich, Sofia, Bulgaria). It has been established that with prolonged storage and an increase in pH and temperature, the anthocyanin content decreased. S. nigra fruits juice was subjected to lyophilization (300 rpm; 10 min; layer thickness 1 cm; −45 °C) and vacuum-sublimation drying in a TG 16.50 installation (Hochvacuum, Munich, Germany) for 24 ± 1 h [85,86].
4.2. High-Performance Liquid Chromatography (HPLC—DAD)
The high-performance liquid chromatography method (1100 HPLC, Agilent Technologies, Santa Clara, CA, USA) with a HPLC–DAD diode detector (G1315B, Agilent Technologies, Santa Clara, CA, USA) operated by an HP Chemstation was used to determine 9 components: (+)-epicatechin gallate; (-)-catechin; (-) epigallocatechin; rutin; quercetin; myricetin; kaempferol; and gallic and vanillic acids [87]. The optimized method was performed at 278 nm–368 nm wavelength, with a C18 reversed-phase column (Purospher star, Hiber RT 125-4; RP18, Purospher star, Merck, Sofia, Bulgaria). The new HPLC method is specific, sensitive, linear, and precise [88]. Separation was performed using a linear gradient elution program with 0.1% TCA (A) and 100% acetonitrile (B) for 40 min. The gradient elution program started with 5% B, 15% B at 16.5 min, 33% B at 22.5 min, 100% B at 30.5 min, and 5% B at 35 min until the 40th equilibration. The flow rate was 1.6 μL min−1, and the column temperature was −25 °C. The injection volume was set to 30 μL, and the DAD acquisition peaks were monitored simultaneously in the 200 ÷ 400 nm range. HPLC–DAD calibration curves were obtained by plotting the limits of the peak area against the standard concentrations (μg mL−1). All samples were filtered through a 0.46 µm acrodisc syringe filter before injection. Calibration curves consisted of five points, from 20 to 100 μg mL−1, for all analytics.
4.3. Antioxidant Ability In Vitro
The antioxidant ability of S. nigra-lyophilized extract was determined using a DPPH (2.2-diphenyl-1-picrylhydrazyl) assay, measured by the Electron Paramagnetic Resonance (EPR-X-Bruker-EMX-micro) spectroscopy [89]. Briefly, 80 mM DPPH ethanol solution was homogenized and incubated with S. nigra-lyophilized extract for 5 min (5 mM, 0.5 mg mL−1). The 0.20 µL was transferred in the Micro-221-EPR cavity at 23 °C. DPPH-H/R generation started immediately. The results were presented as % and recalculated as µg/DPPH radicals possible to be neutralized by 1 mL extract.
4.4. Lymphocyte Cultures and Chromosome Aberration Assay
Lymphocyte cultures were prepared from peripheral vein blood in heparin (30 IU mL−1) of healthy, nonsmoking, non-drinking (female), and drug-free donors aged 38 to 43 years.
Lymphocyte cultures (10 mL) were cultured in sterile flasks containing RPMI-1640 cell medium, 3 mL heat-inactivated normal calf serum, 0.2 mL re-substituted PHA (2%), and antibiotics (penicillin-gentamicin, 100 U mL−1: 50 μg mL−1) at 37 °C for 47 h. S. nigra extract (50 μL, 120 mg kg−1) was added to the lymphocytes at 37 °C for 47 h under aerobic conditions. After 47 h, the cells were incubated with colchicine solution (0.5 g mL−1) according to the classical culture protocol for 60 min. Finally, the culture medium was treated with 0.075 M KCl and 4 × methanol/acetic acid (3:1, v/v) at 23 °C, and the clastogenic effect was examined using the method of Evans, 1984 [90]. Microscopy of chromosomal metaphases stained with 10% Giemsa (Merck); chromosome aberration analysis was at ×1000 magnification (Olympus BX-41, Hamburg, Germany), with a minimum of 100 metaphases (Scheme 2). Untreated lymphocyte samples were used as controls. All procedures followed the Declaration of Helsinki, approved by the Ethics and Academic Integrity Committee of Medical Faculty, Trakia University (No. 7.5.1 OD_4.1.5.9/protocol 18/15.04.2022). The lymphocyte cultures were stored in the Laboratory for Chromosomal Diagnostics and Genetic Monitoring and Screening in the Department of Molecular Biology, Immunology, and Medical Genetics, Medical Faculty, Trakia University, Stara Zagora, Bulgaria. All donors were informed about this experimental procedure and signed written informed consent forms.
Scheme 2.
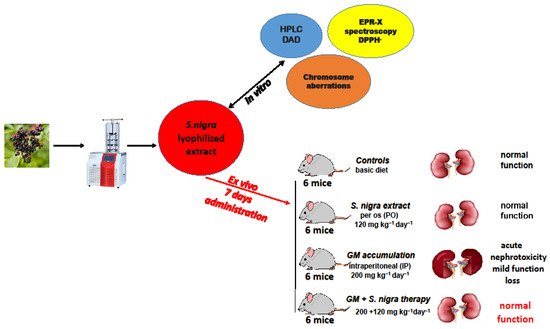
Experimental protocol design of S. nigra fruits juice at lyophilization process, in vitro investigations, and in vivo acute administration.
4.5. Nephrotoxicity Induction and Therapeutic Protection
Thirty-six Balb/c mice (37–39.4 g; 9 weeks old, Neurobiology Institute, Slivnitsa, Bulgaria) used in this study were kept in the animal care facility by the Bioethics Committee, TrU, Stara Zagora, Bulgaria. Animals were maintained at a controlled temperature (21 °C), humidity (52%), and 12 h dark/light 10-day cycle of adaptive feeding and acclimatization, with a license (317/6000-0333/09.12.2021) following Directive 2010/63/EU on the animals’ protection used for experimental and other scientific work.
The GM-induced nephrotoxicity murine model was developed by a daily IP injection of 200 mg kg−1 day−1 for 10 consecutive days, according to previous models [7,15,24]. The GM inducement caused oxidative stress disturbances and generated a redox–homeostatic imbalance cascade, directly affecting the proximal renal tubules.
The S. nigra fruit extract has shown pharmacological properties at doses ranging from 15 to 600 mg kg−1 [91]. Its pharmacological effects may be different depending on the concentration of flavonoid and polyphenolic components. Based on our preliminary in vitro studies [92], the robust antioxidant activity and cytoprotective effect, 120 mg kg−1 S. nigra fruit extract was used in the present study.
The animals were divided into four groups (n = 6, Scheme 2), according to the (1) controls; basal diet (19.6% protein, 4.03% fat, 6.89% fiber, 10.71% moisture; 8.97% ash) was injected IP with 1 mL ice-cold NaCl isotonic solution (0.9%); (2) S. nigra extract only administered per os (PO) (120 mg kg−1 day−1 bw); (3) GM only; GM was injected IP (200 mg kg−1 day−1 b) to induce acute nephrotoxicity; (4) GM + S. nigra therapy; GM was injected IP (200 mg kg−1 bw) and received PO S. nigra-lyophilized extract (120 mg kg−1 day−1 bw) after 2 h.
The S. nigra extract was mixed in cold NaCl isotonic solution (0.9%). The animals’ physiological state and behavior were monitored daily. Twenty-four hours after the last dose, the mice were weighed and anesthetized by IP injection (Nembutal, 50 mg kg−1). Blood samples collected by intra-cardiac technique in serum tubes were centrifuged (4000 rpm, 10 min, 4 °C) and analyzed immediately. The mice’s kidneys were weighed, and the left kidney was fixed in 10% formalin buffer for histological analysis. The right kidney was placed in ice-cold 0.05 M PBS (pH 7.5; 4 °C), homogenized individually, and examined.
4.6. Histopathological Analysis
The left kidney tissue was embedded in paraffin after being perfused, dehydrated by a graded series of ethanol, and fixed in 10% phosphate-buffered formalin for 24 h. The kidney tissues, cut into sections (5 µm), were mounted on gelatin-coated slides, xylene deparaffinized, and 0.1% hematoxylin–eosin (H&E) stain to distinguish significant kidney injuries such as tubular necrosis, fibrosis, inflammatory cells infiltration, tubular dilation, and cast formation.
4.7. Mast Cell (MCs) Number and Collagen Fiber Thickness (CFT)
Next, left kidney tissues were embedded in paraffin after being fixed in 10% formaldehyde/aqueous solution (7 days) and dehydrated. The kidney tissues were cut into slices (5 µm) and stained by toluidine blue (MCs differentiation) and Azan technique (collagen fibrils differentiation and indicating interstitial fibrosis). In short, paraffin sections were xylene deparaffinized, hydrated (ethanol 100%, ethanol 96%, ethanol, ethanol 80%, ethanol 70%), and washed in water. Toluidine blue (0.1%) in McLivane’s buffer (pH = 3) was used for metachromatic MCs [93], and the Azan stain was used for collagen fiber (CFT) visualization stained with blue, erythrocytes with orange, and muscle cells and nuclei with red [94].
4.8. Renal Hydroxyproline (HYP) u Protein Oxidation Analysis
Renal hydroxyproline analysis (at 110 °C for 24 h; hydrolysis with 6N HCl, incubation at 110 °C), used to quantify fibrotic changes, was determined at 550 nm absorption by the Woessner method [57] and expressed as μg/HYR per gram kidney tissue.
The protein oxidation analysis (albumin injuries) in kidneys was assessed by the EPR method in vivo, using spin-conjugation with spin-trap 3-maleimido proxyl (5-MSL). Right kidney tissue (10 mg) was mixed with 20 mM 5-MSL dissolved in 900 μL dimethyl sulfoxide (DMSO). The mixture was centrifuged (1000 rpm; 15 min) at 4 °C. The protein/albumin conformational (-SH) changes were recorded in triplicate, with the following parameters: 3505 G; 6.42 MW power; 5 G amplitude; 12 modulations, in random units, by the method described earlier [58].
4.9. Renal Functional Markers
To monitor renal functional damages, commercial kits to measure the kidney injury molecule-1 (KIM-1; No. MBS175125), cystatin C (CysC), glutathione-S-transferase (GST), and serum levels of gamma-glutamyl-transpeptidase (gamma-GT), creatinine (Cre), and urea (U) were used.
4.10. Oxidative Stress Analysis in Renal Tissue
To monitor renal antioxidant enzymes activities, catalase (CAT; No. ab83464), superoxide dismutase (SOD; No. ab65354), glutathione (GSH; No. ab142044), glutathione peroxidase 1 (GPx1; No. ab41464), as well as the pro-oxidants such as malondialdehyde (MDA; No. ab233471) were investigated by ELISA kits.
The ROS production: A total of 100 µL homogenized kidney tissue was mixed with 900 μL (50 mM) N-tert-butyl-alpha-phenylnitrone (PBN) dissolved in DMSO. The mixture was centrifuged at 4000× g, 10 min at 4 °C, by [95].
The nitric oxide (•NO) generation relative to the spin–adduct formed between the spintrap carboxy 2-(4-carboxyphenyl)-4,4,5,5-tetramethyl (CPTIO.K) and •NO in kidney tissue were based on established EPR methods [96,97]. Briefly, 50 μM CPTIO.K was dissolved in a mixture of 50 mM Tris (pH = 7.5), and DMSO (9:1) was centrifuged at 4000× g for 10 min at 4 °C. Then, 100 μL kidney samples were mixed in 100 μL CPTIO.K, and spin–adducts were recorded.
The superoxide (•O2−) concentration in kidney tissue was determined relative to the spin–adduct formed using the spin-trap CMH (1-hydroxy-3-methoxycarbonyl-2,2,5,5-tetramethylpyrrolidine), based on methods [98,99]. For this purpose, 30 µL kidney tissue was activated in 30 µL CMH (1:1) on an ice bath, and after 5 min incubation, it was prepared. All EPR analyses were performed with fivefold measurement in recorded EPR spectra, with the following characteristics: 3503–3515 G center field; 6.42–20.00 mW microwave power; 5–10 G modulation per sample, and the results are presented in arbitrary units (a.u.)
4.11. Pro-Collagen I Alpha 1, Heme Oxygenase-1, and Pro-Inflammatory Cytokines in Renal Tissue
The PGC-1, the pro-inflammatory IL-1, IL-10, IL-6, ITF-α, and TNF-γ cytokines were measured with ELISA kits.
4.12. Statistical Methods
The aberrance and aberrant cell degrees were processed by one-way ANOVA followed by χ2-Cramer’s V test (GraphPadPrism 6/Windows; GraphPad Software, Inc., USA). Data are presented as mean ± SD; p < 0.05 was considered statistically significant.
The mast cell (MCs) density (number/field of view × 400) was determined on a ×200 microscopic field with an area of 0.163 mm2 in kidney sections of each animal, using a light research microscope (LEIKA DM 1000, Leica Camera AG, Wetzlar, Germany) equipped with a digital camera (LEIKA DFC 290). The data were processed by one-way ANOVA followed by Tukey Kramer’s test (GraphPadPrism 6 for Windows; GraphPad Software, Inc., USA). Data are presented as mean ± SD; p < 0.05 was considered statistically significant. The remaining statistical analyses were performed using Excel version 10.0 software, StaSoft, Inc., San Diego, CA, USA, and presented as mean ± SD.
The EPR processing was performed using WIN-EPR SimFonia 1.2/6130860 software. Statistical analysis was performed using one-way ANOVA and Student’s t-test to determine differences; p < 0.05 was considered statistically significant.
4.13. Limits of This Study
The results related to our study clearly show that the lyophilized extract of S. nigra attenuated renal dysfunction caused by GM application. However, some limitations should be noted that could contribute to the overall significance of the extract on GM-induced nephrotoxicity. Due to ethical considerations, the number of animals studied per group was six, which was sufficient to reveal the most significant differences but might be insufficient to establish additional correlations between some parameters. The results indicate the involvement of some biomarkers, like KIM-1 and PGC-1α, in the process, but further investigation of markers through pathway-specific inhibitors or genetic approaches would better clarify their role. In the present study, in vivo GM activity was not investigated by the co-administration of S. nigra extract, which would also reveal additional information about the antimicrobial properties of the combination therapy. Further studies are needed to identify possible doses that have a beneficial effect on kidney intoxication with longer-term use in protecting ferroptosis processes and the molecular study of DNA damage.
5. Conclusions
The present study highlights the anti-inflammatory potential of S. nigra-lyophilized fruit extract (120 mg mL−1) by reducing fibrotic cell proliferation under GM-induced nephrotoxicity. The antioxidant protection is mediated by a reduction in cytokine expression and albumin modulation. The proposed mechanisms of action of S. nigra are directly related to the redox modulation of ROS, NO•, and •O2−, which facilitate mitochondrial antioxidant and anti-inflammatory properties, associated with the cytoprotection of GM-induced OS and reduced hyperpolarization. In conclusion, we encourage S. nigra use as a traditional supplement to restore antioxidant enzymes in patients with renal injury undergoing GM treatment without compromising the efficacy of bactericidal therapy.
Author Contributions
Conceptualization, K.P.-P. and Y.K.; methodology, K.P.-P., Y.K., I.S., J.A., and N.K.; software, E.G.; validation, K.P.-P., V.P.-T., I.S., N.K., J.A., and E.G.; formal analysis, E.G., G.N., and Y.K.; investigation, K.P.-P., G.N., N.K., and V.P.-T.; resources, K.P.-P., G.N., and Y.K.; data curation, I.S., J.A., P.H., K.P.-P., and Y.K.; writing—original draft preparation, K.P.-P., and Y.K.; writing, K.P.-P., T.G., and Y.K.; visualization, E.G., P.H., and K.P.-P., supervision, G.N.; project administration, K.P.-P.; funding acquisition, G.N. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Bulgarian Ministry of Education and Science (MES) in the frames of the National Research Program “Young Scientists and Postdoctoral Students-2”(u201/D) Trakia University, Bulgaria.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Animal Ethics Commission of the Trakia University, Stara Zagora, Bulgaria, and the Bulgarian Food Safety Agency, Sofia, Bulgaria, with a license (317/06.10.2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study (No. 7.5.1 OD_4.1.5.9/protocol 18/15.04.2022).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Acknowledgments
This study was supported by the University Projects № 10/2022 and № 14/2023, Medical Faculty, Trakia University, Bulgaria.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mahi-Birjand, M.; Yaghoubi, S.; Abdollahpour-Alitappeh, M.; Keshtkaran, Z.; Bagheri, N.; Pirouzi, A.; Karimzadeh, I. Protective effects of pharmacological agents against aminoglycoside-induced nephrotoxicity: A systematic review. Expert Opin. Drug Saf. 2020, 19, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Balgradean, M.; Cinteza, E.; Filipoiu, F.; Jinga, V. Gentamicin, infections, and acute tubular necrosis in children. Farmacia 2013, 61, 772–780. [Google Scholar]
- Sandoval, R.M.; Molitoris, B. Gentamicin traffics retrograde through the secretory pathway and is released in the cytosol via the endoplasmic reticulum. Am. J. Physiol. Renal. Physiol. 2004, 286, F617–F624. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Singh, P.; Aprajita, K.; Kapuganti, J.G. Alternative oxidase plays a role in minimizing ROS and RNS produced under salinity stress in Arabidopsis thaliana. Physiol. Plant. 2022, 2, 174. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef]
- Vilhelmova-Ilieva, N.; Petrova, Z.; Georgieva, A.; Tzvetanova, E.; Trepechova, M.; Mileva, M. Anti-Coronavirus Efficiency and Redox-Modulating Capacity of Polyphenol-Rich Extracts from Traditional Bulgarian Medicinal Plants. Life 2022, 12, 1088. [Google Scholar] [CrossRef]
- Chainy, G.B.; Sahoo, D.K. Hormones and oxidative stress: An overview. Free Radic. Res. 2020, 54, 1–26. [Google Scholar] [CrossRef]
- Ungur, R.A.; Borda, I.M.; Codea, R.A.; Ciortea, V.M.; Năsui, B.A.; Muste, S.; Martiș, G.S. A Flavonoid-Rich Extract of Sambucus nigra L. Reduced Lipid Peroxidation in a Rat Experimental Model of Gentamicin Nephrotoxicity. Materials 2022, 15, 772. [Google Scholar] [CrossRef]
- Kandemir, F.M.; Ozkaraca, M.; Yildirim, B.A.; Hanedan, B.; Kirbas, A.; Kilic, K.; Aktas, E.; Benzer, F. Rutin attenuates gentamicin-induced renal damage by reducing oxidative stress, inflammation, apoptosis, and autophagy in rats. Ren Fail. 2015, 37, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Gu, W.; Gao, Y.; Ma, N.; Fan, C.; Ci, X. Daphnetin Ameliorated GM-Induced renal injury through the suppression of oxidative stress and apoptosis in mice. Int. Immunopharmacol. 2021, 96, 107601. [Google Scholar] [CrossRef]
- Chaudhary, P.; Janmeda, P.; Docea, A.O.; Yeskaliyeva, B.; Abdull Razis, A.F.; Modu, B.; Calina, D.; Sharifi-Rad, J. Oxidative stress, free radicals and antioxidants: Potential crosstalk in the pathophysiology of human diseases. Front. Chem. 2023, 11, 1158198. [Google Scholar] [CrossRef] [PubMed]
- Pena, E.; El Alam, S.; Siques, P.; Brito, J. Oxidative Stress and Diseases Associated with High-Altitude Exposure. Antioxidants 2022, 11, 267. [Google Scholar] [CrossRef] [PubMed]
- Zeima, N.M.; EL-Gawish, A.M. The Prophylactic Effect of some herbs extract on Gentamicin Induced Nephrotoxicity in Albino Rats. Egypt. J. Nutr. Health 2021, 16, 35–52. [Google Scholar] [CrossRef]
- Solanki, S.; Modi, C.M.; Patel, H.B.; Patel, U.D.; Singh, V. Protective Effect of Polyherbal Extract on Gentamicin-Induced Renal Injury in Wistar Rats. Indian J. Pharm. Sci. 2022, 84, 369–379. [Google Scholar] [CrossRef]
- Patil, S.; Ramu, R.; Shirahatti, P.; Shivamallu, C.; Amachawadi, R. A systematic review on ethnopharmacology, phytochemistry and pharmacological aspects of Thymus vulgaris Linn. Heliyon 2021, 7, e07054. [Google Scholar] [CrossRef] [PubMed]
- Tahmouzi, S.; Salek Nejat, M. New infertility therapy effects of polysaccharides from Althaea officinalis leaf with emphasis on characterization, antioxidant and anti-pathogenic activity. Int. J. Biol. Macromol. 2020, 145, 777–787. [Google Scholar] [CrossRef]
- Jakovljević, M.R.; Grujičić, D.; Vukajlović, J.T.; Marković, A.; Milutinović, M.; Stanković, M.; Vuković, N.; Vukić, M.; Milošević-Djordjević, O. In vitro study of genotoxic and cytotoxic activities of methanol extracts of Artemisia vulgaris L. and Artemisia alba Turra. S. Afr. J. Bot. 2020, 132, 117–126. [Google Scholar] [CrossRef]
- Owczarek, A.; Kolodziejczyk-Czepas, J.; Wozniak-Serwata, J.; Magiera, A.; Kobiela, N.; Wasowicz, K.; Olszewska, M.A. Potential Activity Mechanisms of Aesculus hippocastanum Bark: Antioxidant Effects in Chemical and Biological In Vitro Models. Antioxidant 2021, 10, 995. [Google Scholar] [CrossRef]
- Georgieva, S.; Georgieva, A.; Peteva, Z.; Dimova, D. Trace elements in commonly used medicinal plants from Varna region, Bulgaria. ESPR 2021, 28, 59277–59283. [Google Scholar] [CrossRef] [PubMed]
- Koleva, P.; Tsanova-Savova, S.; Paneva, S.; Velikov, S.; Savova, Z. Polyphenols content of selected medical plants and food supplements present at Bulgarian market. Pharmacia 2021, 68, 819–826. [Google Scholar] [CrossRef]
- Ignatov, I. Spectral analysis of Sambucus nigra L. fruits and flowers for elucidation of their analgesic, diuretic, anti-inflammatory and anti-tumor effects. Plant. Cell. Biotechnol. 2021, 22, 134–140. [Google Scholar]
- Dominguez, R.; Zhang, L.; Rocchetti, G.; Lucini, L.; Pateiro, M.; Munekata, P.E.S.; Lorenzo, J.M. Elderberry (Sambucus nigra L.) as potential source of antioxidants. Characterization, optimization of extraction parameters and bioactive properties. Food Chem. 2020, 330, 127266. [Google Scholar] [CrossRef]
- Ungur, R.; Buzatu, R.; Lacatus, R.; Purdoiu, R.C.; Petrut, G.; Codea, R.; Sarpataky, O.; Biris, A.; Popovici, C.; Mircean, M. Evaluation of the Nephroprotective Effect of Sambucus nigra Total Extract in a Rat Experimental Model of Gentamicine Nephrotoxicity. Rev. Chim. 2019, 17, 1971–1974. [Google Scholar] [CrossRef]
- Kiselova-Kaneva, Y.; Nashar, M.; Roussev, B.; Salim, A.; Hristova, M.; Olczyk, P.; Komosinska-Vassev, K.; Dincheva, I.; Badjakov, I.; Galunska, B.; et al. Sambucus ebulus (Elderberry) Fruits Modulate Inflammation and Complement System Activity in Humans. Intern. J Mol. Sci. 2023, 24, 8714. [Google Scholar] [CrossRef]
- Abdelkader, R.S.E.; El-Beih, N.M.; Zaahkouk, S.A.; El-Hussieny, E.A. Ameliorative Effect of Eruca sativa Seeds and Its Rutin on Gentamicin-Induced Nephrotoxicity in Male Rats via Targeting Inflammatory Status, Oxidative Stress and Kidney Injury Molecule-1 (KIM-1)/Cystatin C Expression. Indones. Biomed. J. 2022, 14, 74–83. [Google Scholar] [CrossRef]
- Rahmani, S.; Naraki, K.; Roohbakhsh, A.; Hayes, A.W.; Karimi, G. The protective effects of rutin on the liver, kidneys, and heart by counteracting organ toxicity caused by synthetic and natural compounds. Food Sci. Nutr. 2022, 11, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Zielińska-Wasielica, J.; Olejnik, A.; Kowalska, K.; Olkowicz, M.; Dembczyński, R. Elderberry (Sambucus nigra L.) Fruit Extract Alleviates Oxidative Stress, Insulin Resistance, and Inflammation in Hypertrophied 3T3-L1 Adipocytes and Activated RAW 264.7 Macrophages. Foods 2019, 8, 326. [Google Scholar] [CrossRef]
- Marțiș, G.S.; Mureșan, V.; Marc, R.M.; Mureșan, C.C.; Pop, C.R.; Buzgău, G.; Muste, S. The Physicochemical and Antioxidant Properties of Sambucus nigra L. and Sambucus nigra Haschberg during Growth Phases: From Buds to Ripening. Antioxidants 2021, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Abu Almaaty, A.H.; Elmasry, R.A.; Farrag, M.S.; Althobaiti, F.; Aldhahrani, A.; Fayad, E.; Hussain, M.A. Impact of Human Umbilical Cord Blood Mononuclear Cells on Gentamicin-Induced Renal Injury and Genotoxicity in Rats. Front. Med. 2021, 8, 689691. [Google Scholar] [CrossRef]
- Guerreiro, Í.; Ferreira-Pêgo, C.; Carregosa, D.; Santos, C.N.; Menezes, R.; Fernandes, A.S.; Costa, J.G. Polyphenols and Their Metabolites in Renal Diseases: An Overview. Foods 2022, 11, 1060. [Google Scholar] [CrossRef] [PubMed]
- Albalawi, R.S.; Binmahfouz, L.S.; Hareeri, R.H.; Shaik, R.A.; Bagher, A.M. Parthenolide Phytosomes Attenuated Gentamicin-Induced Nephrotoxicity in Rats via Activation of Sirt-1, Nrf2, OH-1, and NQO1 Axis. Molecules 2023, 28, 2741. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Novoa, J.M.; Quiros, Y.; Vicente, L.; Morales, A.I.; Lopez-Hernandez, F.J. New Insights into the Mechanism of Aminoglycoside Nephrotoxicity: An Integrative Point of View. Kidney Int. 2011, 79, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Srivastav, S.; Sharanagat, V.S. Ultrasound assisted extraction (UAE) of bioactive compounds from fruit and vegetable processing by-products: A review. Ultrason Sonochem. 2021, 70, 105325. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Rajendran, S.; Rajagopal, P.; Selvaraj Jayaraman, J.M.; Kasturi, R.; Nagarajan, V.; Karuppan, M. A review on dietary poly phenols: Herbal neutraceuticals to combat nephrotoxicity. NVEO 2021, 8, 7584–7597. [Google Scholar]
- Terzić, M.; Majkić, T.; Zengin, G.; Beara, I.; Cespedes-Acuña, C.L.; Čavić, D.; Radojković, M. Could elderberry fruits processed by modern and conventional drying and extraction technology be considered a valuable source of health-promoting compounds? Food Chem. 2023, 405, 134766. [Google Scholar] [CrossRef]
- Teng, H.; Chen, L. Polyphenols and bioavailability: An update. Crit. Rev. Food Sci. Nutr. 2019, 59, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Hfaiedh, M.; Brahmi, D.; Zourgui, M.N.; Zourgui, L. Phytochemical analysis and nephroprotective effect of cactus (Opuntia ficus-indica) cladodes on sodium dichromate-induced kidney injury in rats. Appl. Physiol. Nutr. Metab. 2019, 44, 239–247. [Google Scholar] [CrossRef]
- Lee, D.; Yu, J.S.; Lee, S.R.; Hwang, G.S.; Kang, K.S.; Park, J.G.; Kim, H.Y.; Kim, K.H.; Yamabe, N. Beneficial Effects of Bioactive Compounds in Mulberry Fruits against Cisplatin-Induced Nephrotoxicity. Int. J. Mol. Sci. 2018, 19, 1117. [Google Scholar] [CrossRef] [PubMed]
- Putri, R.S.; Putra, A.; Chodidjah, D.M.D.; Trisnadi, S.; Thomas, S.; Amalina, N.D.; Irawan, R.C. Clitorea ternatea flower extract induces platelet-derived growth factor (PDGF) and GPx gene overexpression in ultraviolet (UV) B irradiation-induced collagen loss. Off. Publ. Med. Assoc. Zenica-Doboj Canton Bosnia Herzeg. 2023, 20, 15–21. [Google Scholar] [CrossRef]
- Przybylska-Balcerek, A.; Szablewski, T.; Szwajkowska-Michałek, L.; Świerk, D.; Cegielska-Radziejewska, R.; Krejpcio, Z.; Stuper-Szablewska, K. Sambucus nigra extracts–natural antioxidants and antimicrobial compounds. Molecules 2021, 26, 2910. [Google Scholar] [CrossRef]
- Sobreiro, M.A.; Della Torre, A.; de Araújo, M.E.M.B.; Canella, P.R.B.C.; de Carvalho, J.E.; Carvalho, P.D.O.; Ruiz, A.L.T.G. Enzymatic hydrolysis of rutin: Evaluation of kinetic parameters and anti-proliferative, mutagenic and anti-mutagenic effects. Life 2023, 13, 549. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Silva, A.M.; Nunes, F.M. Sambucus nigra L. fruits and flowers: Chemical composition and related bioactivities. Food Rev. Int. 2022, 38, 1237–1265. [Google Scholar] [CrossRef]
- Medić, B.; Stojanović, M.; Rovčanin, B.; Kekić, D.; Škodrić, S.R.; Jovanović, G.B.; Vujović, K.S.; Divac, N.; Stojanović, R.; Radenković, M.; et al. Pioglitazone Attenuates Kidney Injury in an Experimental Model of Gentamicin-Induced Nephrotoxicity in Rats. Sci. Rep. 2019, 9, 13689. [Google Scholar] [CrossRef] [PubMed]
- Walz, B.; Chrubasik, S. Effect of a proprietary Sambucus nigra L. concentrate on urinary PH. Fitother. Res. 2008, 22, 977–978. [Google Scholar] [CrossRef]
- Gamaan, M.A.; Zaky, H.S.; Ahmed, H.I. Gentamicin-induced nephrotoxicity: A mechanistic approach. Azhar Int. J. Pharm. Med. Sci. 2023, 3, 11–19. [Google Scholar] [CrossRef]
- Moghadam, A.; Khozani, T.T.; Mafi, A.; Namavar, M.R.; Dehghani, F. Effects of Platelet-Rich Plasma on Kidney Regeneration in Gentamicin-Induced Nephrotoxicity. J. Korean Med. Sci. 2017, 32, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Beshay, O.N.; Ewees, M.G.; Abdel-Bakky, M.S.; Hafez, S.M.N.A.; Abdelrehim, A.B.; Bayoumi, A.M. Resveratrol reduces gentamicin-induced EMT in the kidney via inhibition of reactive oxygen species and involving TGF-β/Smad pathway. Life Sci. 2020, 258, 118178. [Google Scholar] [CrossRef]
- Ibrahim, Y.F.; Hammady, S.H.; Rifaai, R.A.; Waz, S.; Ibrahim, M.A.; Hafez, H.M. Dose-dependent ameliorating effect of lipoxin A4 on gentamicin-induced nephrotoxicity in rats: The role of TNFα, TGF-β, ICAM-1, and JNK signaling. Chem. Interact. 2022, 366, 110139. [Google Scholar] [CrossRef]
- Khan, M.U.; Basist, P.; Zahiruddin, S.; Ibrahim, M.; Parveen, R.; Krishnan, A.; Ahmad, S. Nephroprotective potential of Boerhaavia diffusa and Tinospora cordifolia herbal combination against diclofenac induced nephrotoxicity. S. Afr. J. Bot. 2022, 151, 238–247. [Google Scholar] [CrossRef]
- Basist, P.; Parveen, B.; Zahiruddin, S.; Gautam, G.; Parveen, R.; Khan, M.A.; Krishnan, A.; Shahid, M.; Ahmad, S. Potential nephroprotective phytochemicals: Mechanism and future prospects. J. Ethnopharmacol. 2022, 283, 114743. [Google Scholar] [CrossRef] [PubMed]
- Khajevand-Khazaei, M.R.; Mohseni-Moghaddam, P.; Hosseini, M.; Gholami, L.; Baluchnejadmojarad, T.; Roghani, M. Rutin, a quercetin glycoside, alleviates acute endotoxemic kidney injury in C57BL/6 mice via suppression of inflammation and up-regulation of antioxidants and SIRT1. Eur. J. Pharmacol. 2018, 833, 307–313. [Google Scholar] [CrossRef]
- Zhou, Y.; Vaidya, V.S.; Brown, R.P.; Zhang, J.; Rosenzweig, B.A.; Thompson, K.L.; Miller, T.J.; Bonventre, J.V.; Goering, P.L. Comparison of Kidney Injury Molecule-1 and Other Nephrotoxicity Biomarkers in Urine and Kidney Following Acute Exposure to Gentamicin, Mercury, and Chromium. Toxicol. Sci. 2008, 101, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Wang, H.W.; Yao, S.; Zhou, Y.; Liu, S.C.; Shen, X.L. The natural sources, extraction, and nephroprotective function of oleanolic acid and ursolic acid: A review. Food Health 2024, 6, 13. [Google Scholar] [CrossRef]
- Woessner Jr, J.F. The determination of hydroxyproline in tissue and protein samples containing small proportions of this imino acid. Arch. Biochem. Biophys. 1961, 93, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, K.; Saito, K.; Ueda, J.I.; Anzai, K.; Ozawa, T. Kinetic study on ESR signal decay of nitroxyl radicals, potent redox probes for in vivo ESR spectroscopy, caused by reactive oxygen species. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2002, 1573, 156–164. [Google Scholar] [CrossRef]
- Gamede, M.; Mabuza, L.; Ngubane, P.; Khathi, A. Preventing the onset of diabetes-induced chronic kidney disease during prediabetes: The effects of oleanolic acid on selected markers of chronic kidney disease in a diet-induced prediabetic rat model. Biomed. Pharmacother. 2021, 139, 111570. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Kim, H.M.; Kang, J.S.; Lee, E.Y.; Yadav, D.; Kwon, M.H.; Kim, Y.M.; Kim, H.S.; Chung, C.H. Oleanolic acid and N-acetylcysteine ameliorate diabetic nephropathy through reduction of oxidative stress and endoplasmic reticulum stress in a type 2 diabetic rat model. Nephrol. Dial. Transplant. 2015, 31, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, T.C.; Frick, K.; Emde, B.; Czasch, S.; Landenberg, F.V.; Hewitt, P. Evaluation of novel acute urinary rat kidney toxicity biomarker for subacute toxicity studies in preclinical trials. Toxicol. Pathol. 2012, 40, 1031–1048. [Google Scholar] [CrossRef]
- Olas, B.; Różański, W.; Urbańska, K.; Sławińska, N.; Bryś, M. New Light on Plants and Their Chemical Compounds Used in Polish Folk Medicine to Treat Urinary Diseases. Pharmaceuticals 2024, 17, 435. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, J.; Kong, H.; Kim, Y.S. Ameliorative effects of elderberry (Sambucus nigra L.) extract and extract-derived monosaccharide-amino acid on H2O2-induced decrease in testosterone-deficiency syndrome in a TM3 Leydig cell. PLoS ONE 2024, 19, e0302403. [Google Scholar] [CrossRef]
- Yue, L.; Yang, Y.R.; Ma, W.X.; Wang, H.Y.; Fan, Q.W.; Wang, Y.Y.; Li, C.; Wang, J.; Hu, Z.M.; Wang, X.F.; et al. Epigallocatechin Gallate Attenuates Gentamicin-Induced Nephrotoxicity by Suppressing Apoptosis and Ferroptosis. Molecules 2022, 27, 8564. [Google Scholar] [CrossRef]
- Georgiev, T.; Nikolova, G.; Dyakova, V.; Karamalakova, Y.; Georgieva, E.; Ananiev, J.; Ivanov, V.; Hadzhibozheva, P. Vitamin E and silymarin reduce oxidative tissue damage during gentamycin-induced nephrotoxicity. Pharmaceuticals 2023, 16, 1365. [Google Scholar] [CrossRef]
- Yuan, M.; Briscese, K.; Hong, T.S.; Brunetti, L. Natural products for the prevention of antibiotic-associated kidney injury. Curr. Opinion Toxicol. 2022, 32, 100363. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Chen, T.; Hao, Z.; Yang, X.; Wang, M.; Zhang, Z.; Hao, S.; Lang, F.; Hao, H. Oxymatrine Alleviates Gentamicin-Induced Renal Injury in Rats. Molecules 2022, 27, 6209. [Google Scholar] [CrossRef] [PubMed]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Staniek, H.; Kidoń, M.; Łysiak, G.P. The content of selected minerals, bioactive compounds, and the antioxidant properties of the flowers and fruit of selected cultivars and wildly growing plants of Sambucus nigra L. Molecules 2020, 25, 876. [Google Scholar] [CrossRef]
- Uhl, K.; Mitchell, A.E. Elderberry, An Ancient Remedy: A Comprehensive Study of the Bioactive Compounds in Three Sambucus nigra L. Subspecies. Ann. Rev. Food Sci.Technol. 2024, 15, 27–51. [Google Scholar] [CrossRef] [PubMed]
- Akbaribazm, M.; Goodarzi, N.; Rahimi, M.; Naseri, L.; Khazaei, M. Anti-inflammatory, anti-oxidative and anti-apoptotic effects of Heracleum persicum L. extract on rats with gentamicin-induced nephrotoxicity. Asian Pac. J. Trop. Biomed. 2021, 11, 47. [Google Scholar] [CrossRef]
- Abo, M.; Weerapana, E. Chemical probes for redox signaling and oxidative stress. Antioxid. Redox Signal. 2019, 30, 1369–1386. [Google Scholar] [CrossRef] [PubMed]
- Babić, N.; Peyrot, F. Molecular probes for evaluation of oxidative stress by in vivo EPR spectroscopy and imaging: State-of-the-art and limitations. Magnetochemistry 2019, 5, 13. [Google Scholar] [CrossRef]
- Lewandowski, M.; Gwozdzinski, K. Nitroxides as Antioxidants and Anticancer Drugs. Int. J. Mol. Sci. 2017, 18, 2490. [Google Scholar] [CrossRef] [PubMed]
- Lacy, M.K.; Nicolau, D.P.; Nightingale, C.H.; Quintiliani, R. The pharmacodynamics of aminoglycosides. Clinic. Infect. Dis. 1998, 27, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Bārzdiņa, A.; Plotniece, A.; Sobolev, A.; Pajuste, K.; Bandere, D.; Brangule, A. From polymeric nanoformulations to polyphenols—Strategies for enhancing the efficacy and drug delivery of gentamicin. Antibiotics 2024, 13, 305. [Google Scholar] [CrossRef] [PubMed]
- Wani, R.; Nagata, A.; Murray, B.W. Protein Redox Chemistry: Post-Translational Cysteine Modifications That Regulate Signal Transduction and Drug Pharmacology. Front. Pharmacol. 2014, 5, 224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ye, Z.-W.; Singh, S.; Townsend, D.M.; Tew, K.D. An Evolving Understanding of the S-Glutathionylation Cycle in Pathways of Redox Regulation. Free Radic. Biol. Med. 2018, 120, 204–216. [Google Scholar] [CrossRef]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Aparicio-Trejo, O.E.; Pedraza-Chaverri, J. Mitochondrial redox signaling and oxidative stress in kidney diseases. Biomolecules 2021, 11, 1144. [Google Scholar] [CrossRef]
- Abu Shelbayeh, O.; Arroum, T.; Morris, S.; Busch, K.B. PGC-1α Is a Master Regulator of Mitochondrial Lifecycle and ROS Stress Response. Antioxidants 2023, 12, 1075. [Google Scholar] [CrossRef] [PubMed]
- Amigó, M.; Payá, M.; Braza-Boïls, A.; De Rosa, S.; Terencio, M.C. Avarol Inhibits TNF-α Generation and NF-κB Activation in Human Cells and in Animal Models. Life Sci. 2008, 82, 256–264. [Google Scholar] [CrossRef]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, J.; Zhou, F.; Wang, W.; Chen, N. PGC-1α ameliorates kidney fibrosis in mice with diabetic kidney disease through an antioxidative mechanism. Mol. Med. Rep. 2018, 17, 4490–4498. [Google Scholar] [CrossRef] [PubMed]
- Rius-Pérez, S.; Torres-Cuevas, I.; Millán, I.; Ortega, Á.L.; Pérez, S. PGC-1α, Inflammation, and Oxidative Stress: An Integrative View in Metabolism. Oxidative Med. Cell. Longev. 2020, 2020, 1452696. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Bai, Y.; Chen, X.; Wang, W.; Zhang, C.; Shang, Z.; Zhang, Z.; Li, J.; Cao, M.; Zhu, Z.; et al. Synergistic effects and mechanisms of action of rutin with conventional antibiotics against Escherichia coli. Int. J. Mol. Sci. 2024, 25, 13684. [Google Scholar] [CrossRef] [PubMed]
- Santin, J.R.; Benvenutti, L.; Broering, M.F.; Nunes, R.; Goldoni, F.C.; Patel, Y.B.K.; Quintão, N.L.M. Sambucus nigra: A traditional medicine effective in reducing inflammation in mice. J. Ethnopharmacol. 2022, 283, 114736. [Google Scholar] [CrossRef] [PubMed]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Łysiak, G.P. Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 2018, 40, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Ciurzynska, A.; Lenart, A. Freeze-drying-application in food processing and biotechnology-a review. Polish J. Food Nutr. Sci. 2011, 61, 165–171. [Google Scholar] [CrossRef]
- Dimcheva, V.; Kaloyanov, N.; Karsheva, M. HPLC-DAD method for simultaneous determination of natural polyphenols. Open J. Anal. Bioanal. Chem. 2019, 3, 039–043. [Google Scholar] [CrossRef]
- Bernardo dos Santos, D.H.S.; Silva, V.S. Bolzani. Antioxidant Properties of Plant Extracts: An EPR and DFT Comparative Study of the Reaction with DPPH, TEMPOL and Spin Trap DMPO. J. Braz. Chem. Soc. 2009, 20, 1483–1492. [Google Scholar] [CrossRef]
- Evans, H. Handbook of Mutagenicity Test Procedure; Kilbey, B., Legator, M., Nicols, W., Ramel, C., Eds.; Elsevier Science Publishers BV: Amsterdam, The Netherlands, 1984; pp. 405–427. ISBN 9780444600981. [Google Scholar]
- Cicek, B.; Danısman, B.; Yildirim, S.; Yuce, N.; Nikitovic, D.; Bolat, I.; Kuzucu, M.; Ceyran, E.; Bardas, E.; Golokhvast, K.S.; et al. Flavonoid-Rich Sambucus nigra Berry Extract Enhances Nrf2/HO-1 Signaling Pathway Activation and Exerts Antiulcerative Effects In Vivo. Int. J. Mol. Sci. 2023, 24, 15486. [Google Scholar] [CrossRef]
- Petkova-Parlapanska, K. Characterization and Utilization of Active Antioxidants from Black Elder. Ph.D. Thesis, University of Chemical Technology and Metallurgy, Sofia, Bulgaria, 2022. Reference number: 0237. Available online: https://mmu2.uctm.edu/uctm/downloads/NS_AD/NS/Kamelia_Petkova-Parlapanska/Auto_K_Parlapanska.pdf (accessed on 5 January 2025).
- Gobbo, F.; Sarli, G.; De Silva, M.; Galiazzo, G.; Chiocchetti, R.; Morini, M. A double histochemical/immunohistochemical staining for the identification of canine mast cells in light microscopy. Vet. Sci. 2021, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, J.; Schäfer, E.; Dammaschke, T. Histological determination of cariously altered collagen after dentin caries excavation with the polymer bur PolyBur P1 in comparison to a conventional bud bur. Head Face Med. 2019, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Sui, Y.; Wang, X.; Luo, Y.; Ji, L. Hydroxyl radical production and oxidative damage induced by cadmium and naphthalene in liver of Carassius auratus. Comp. Biochem. Phys. Part C Toxicol. Pharmacol. 2005, 140, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, T.; Iwamoto, N.; Lto, K. An applicationof Electron Paramagnetic Resonance to evaluate nitric oxide and its quenchers. J. Am. Soc. Nephrol. 1996, 7, 961–965. [Google Scholar] [CrossRef]
- Yokoyama, K.; Hashiba, K.; Wakabayashi, H.; Hashimoto, K.; Saton, K.; Kurihara, T.; Motohashi, N.; Sakagami, H. Inhibition of LPS-stimulated NO production in mouse macrophage-like cells by Tropolones. Anticancer Res. 2004, 24, 3917–3922. [Google Scholar]
- Gielis, J.F.; Boulet, G.A.; Briedé, J.J.; Horemans, T.; Debergh, T.; Kussé, M.; Van Schil, P.E. Longitudinal quantification of radical bursts during pulmonary ischaemia and reperfusion. Eur. J. Cardiothorac. Surg. 2015, 48, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Santacroce, A.; Longini, M.; Proietti, F.; Bazzini, F.; Buonocore, G. The free radical diseases of prematurity: From cellular mechanisms to bedside. Oxidative Med. Cell. Longev. 2018, 2018, 7483062. [Google Scholar] [CrossRef] [PubMed]
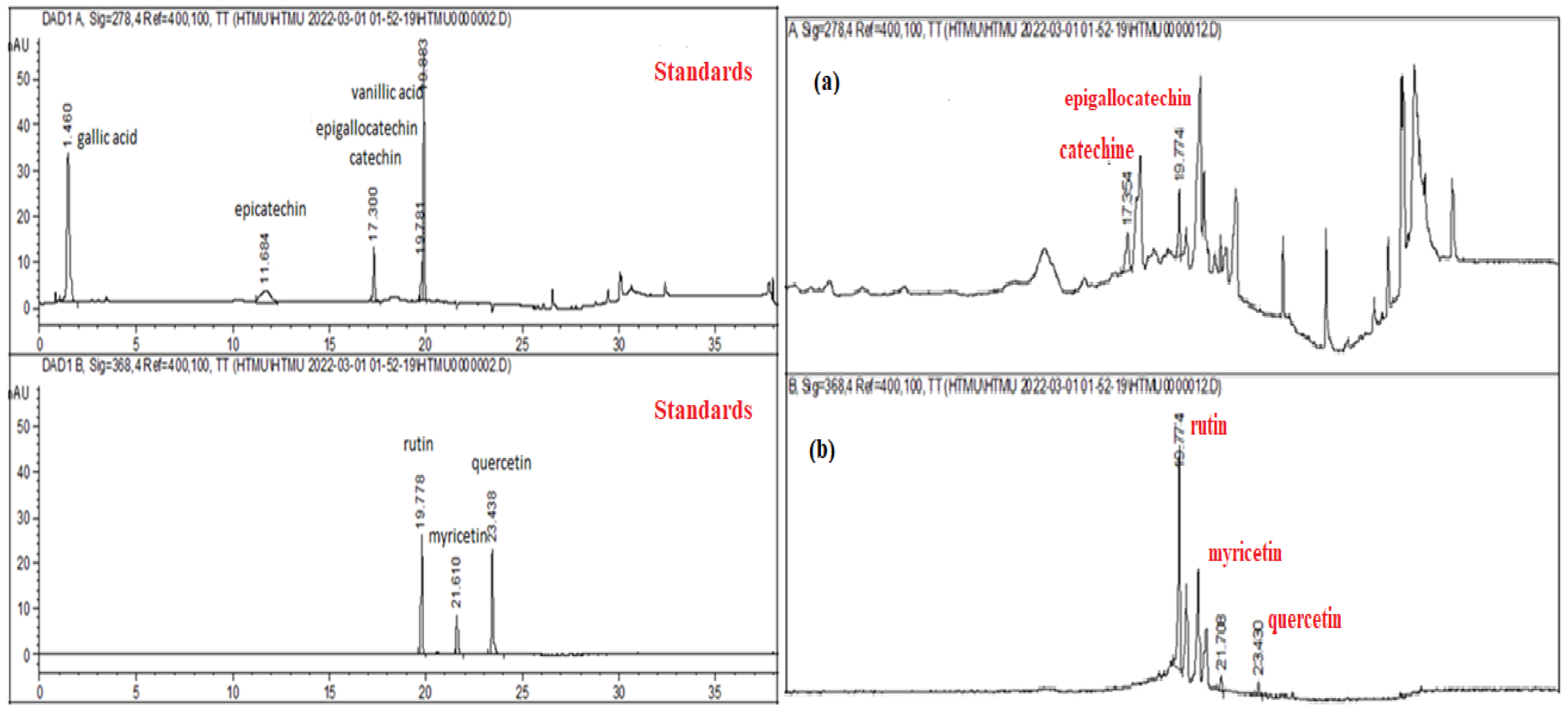
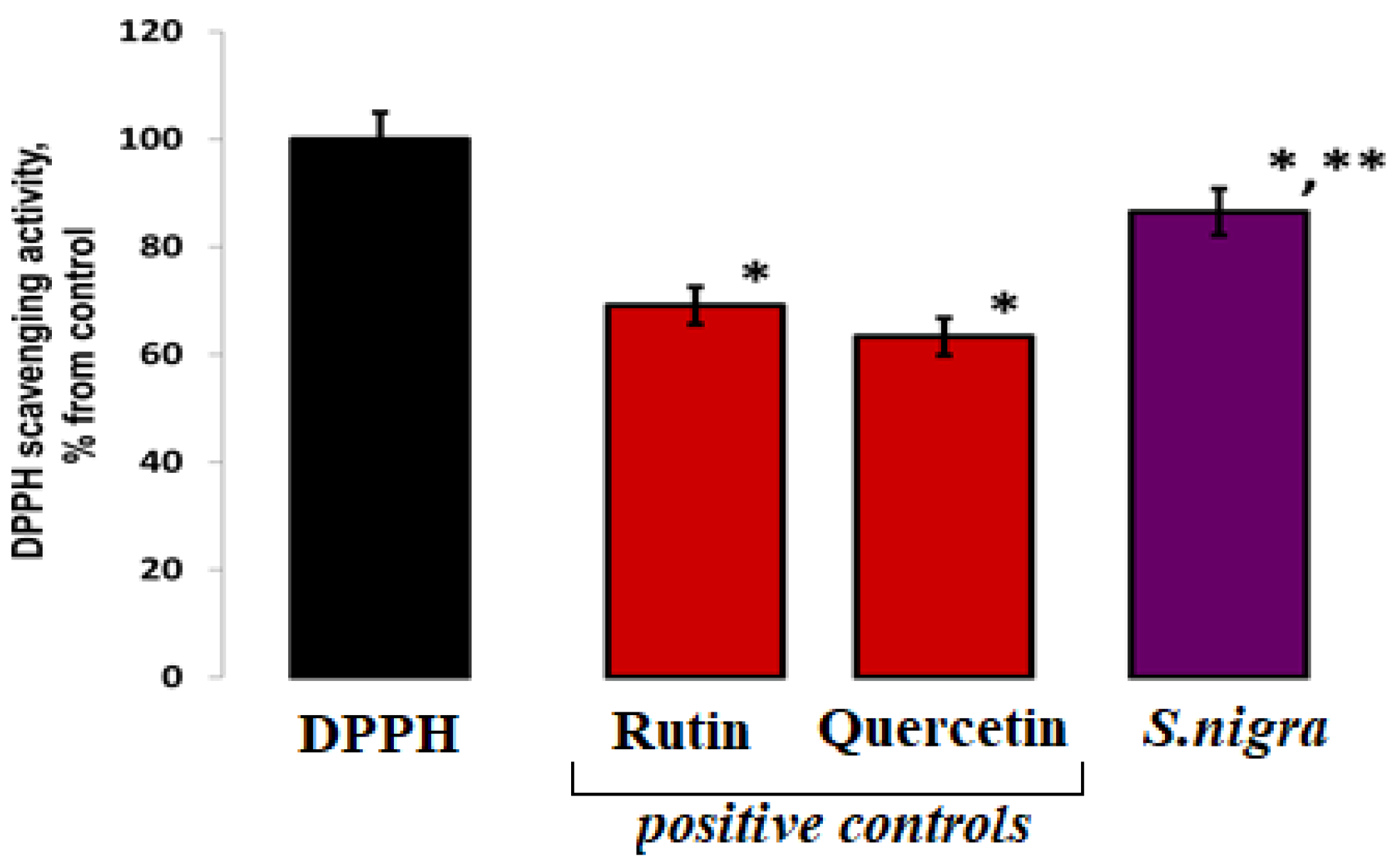
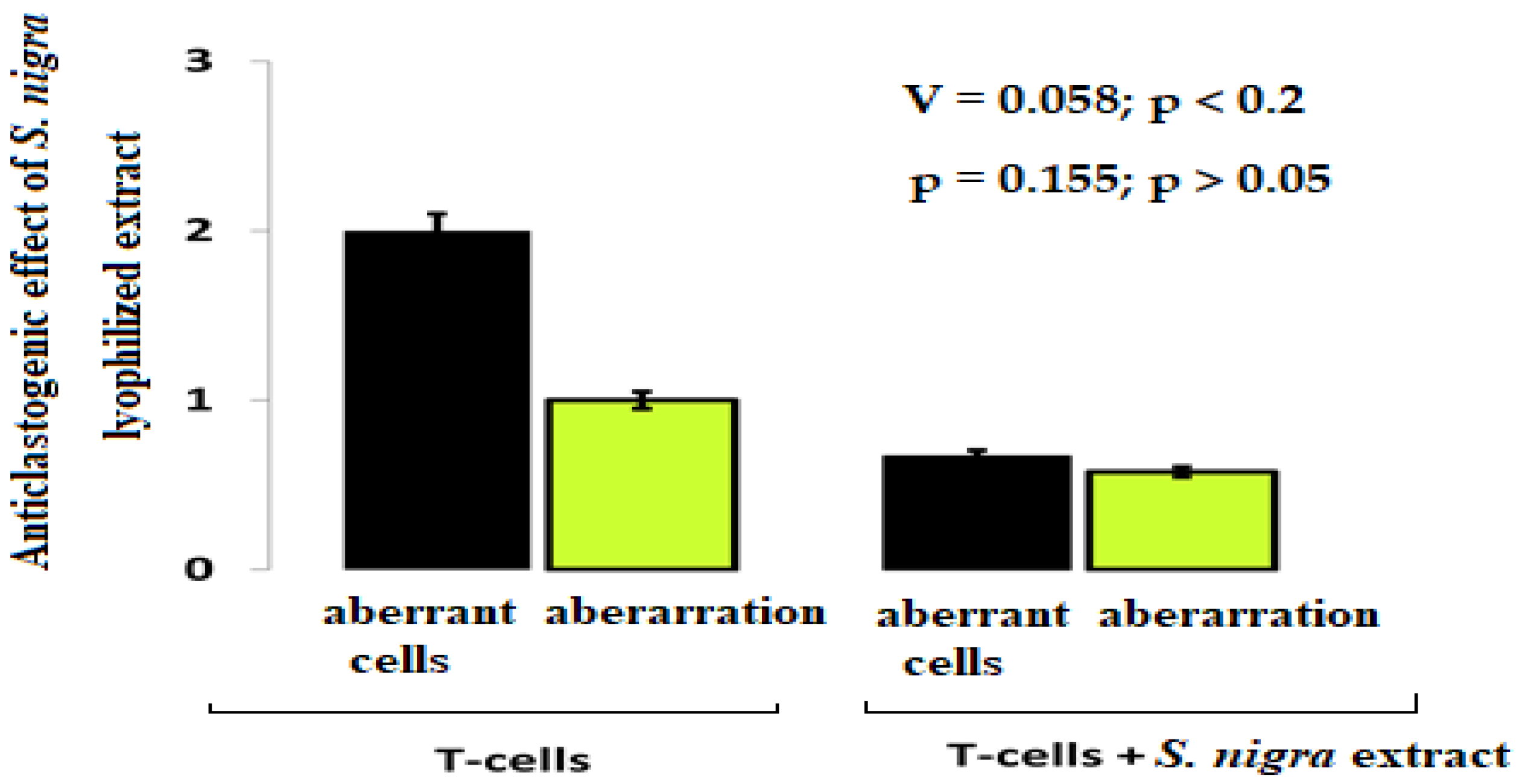
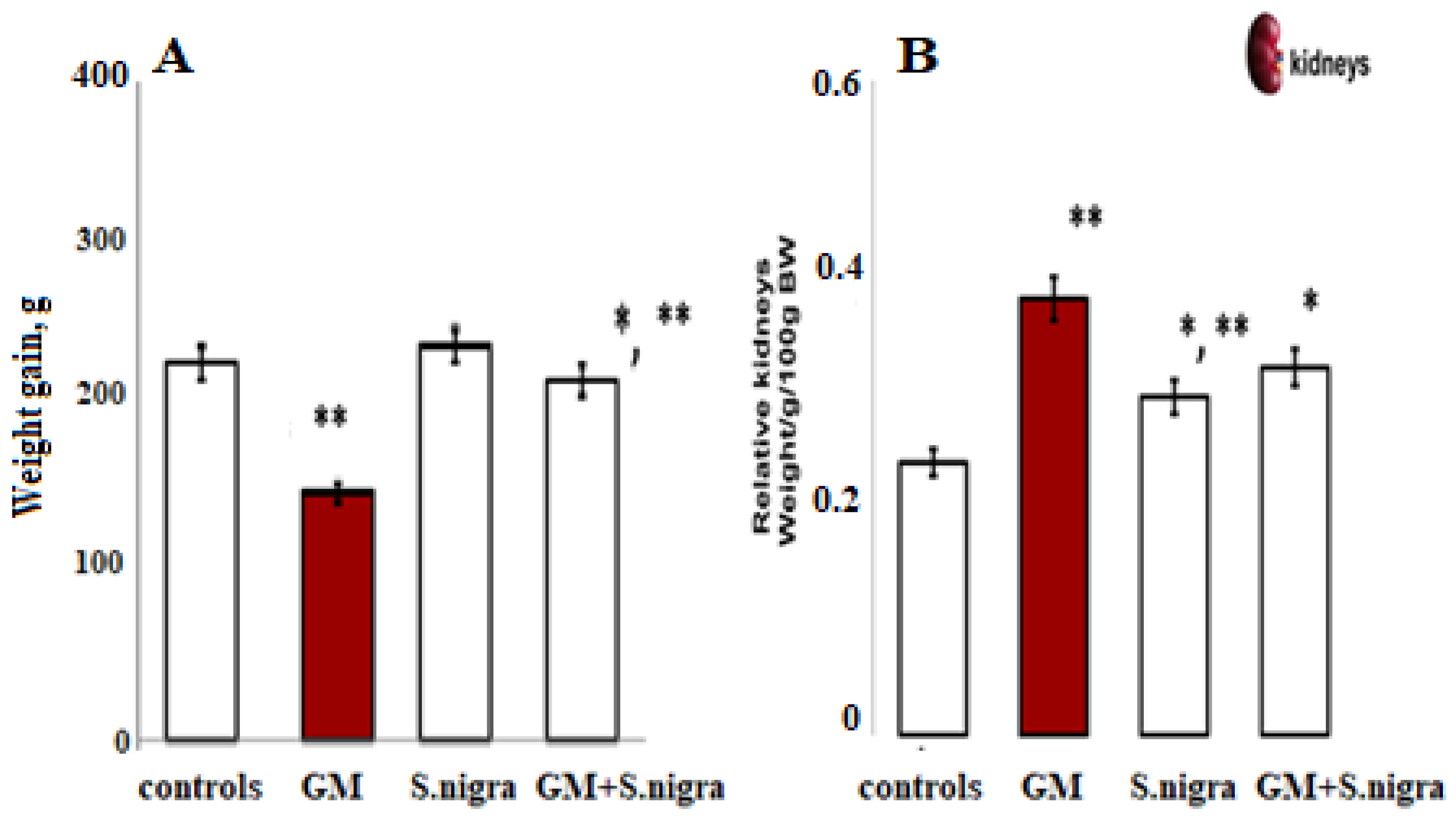
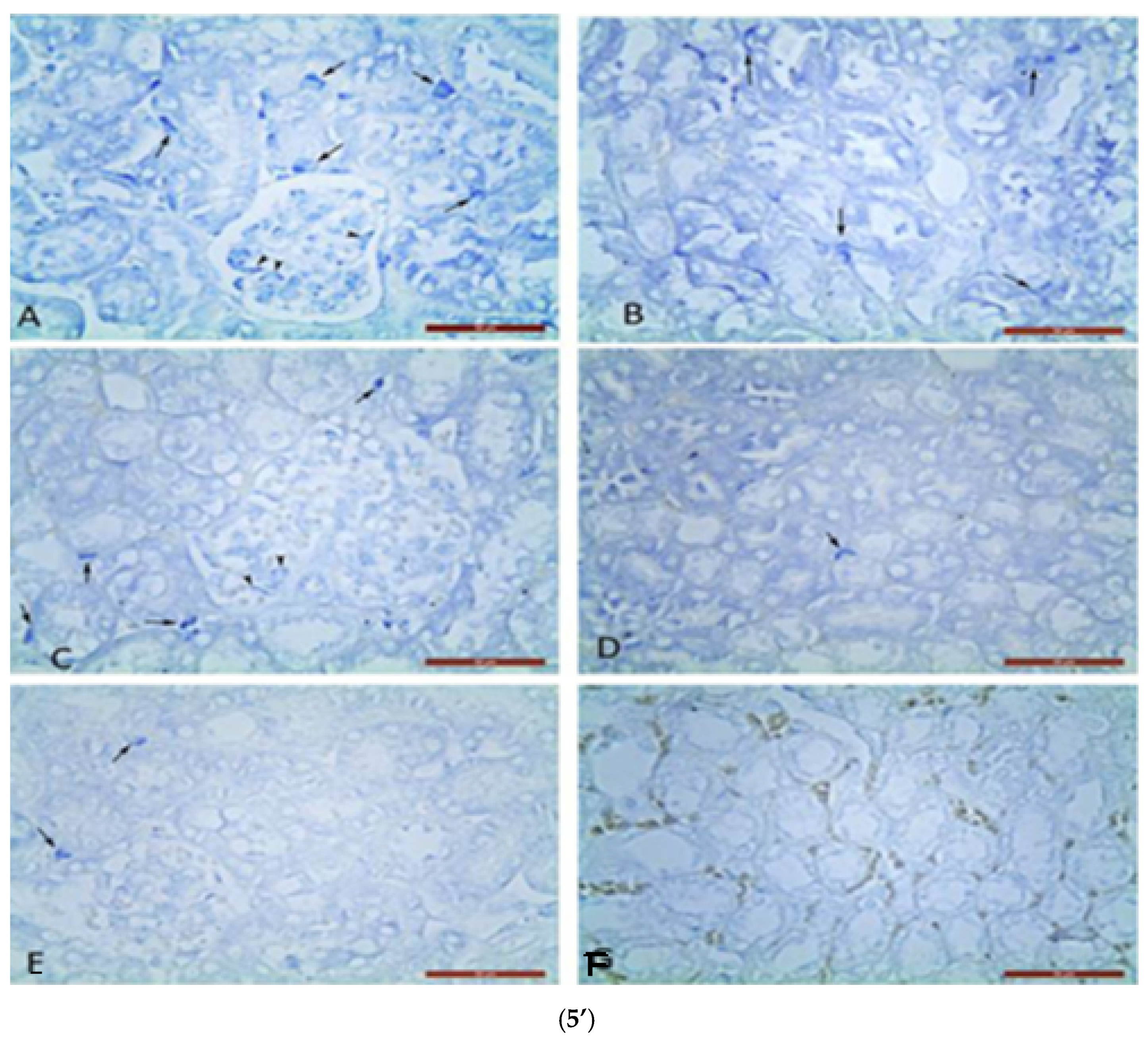

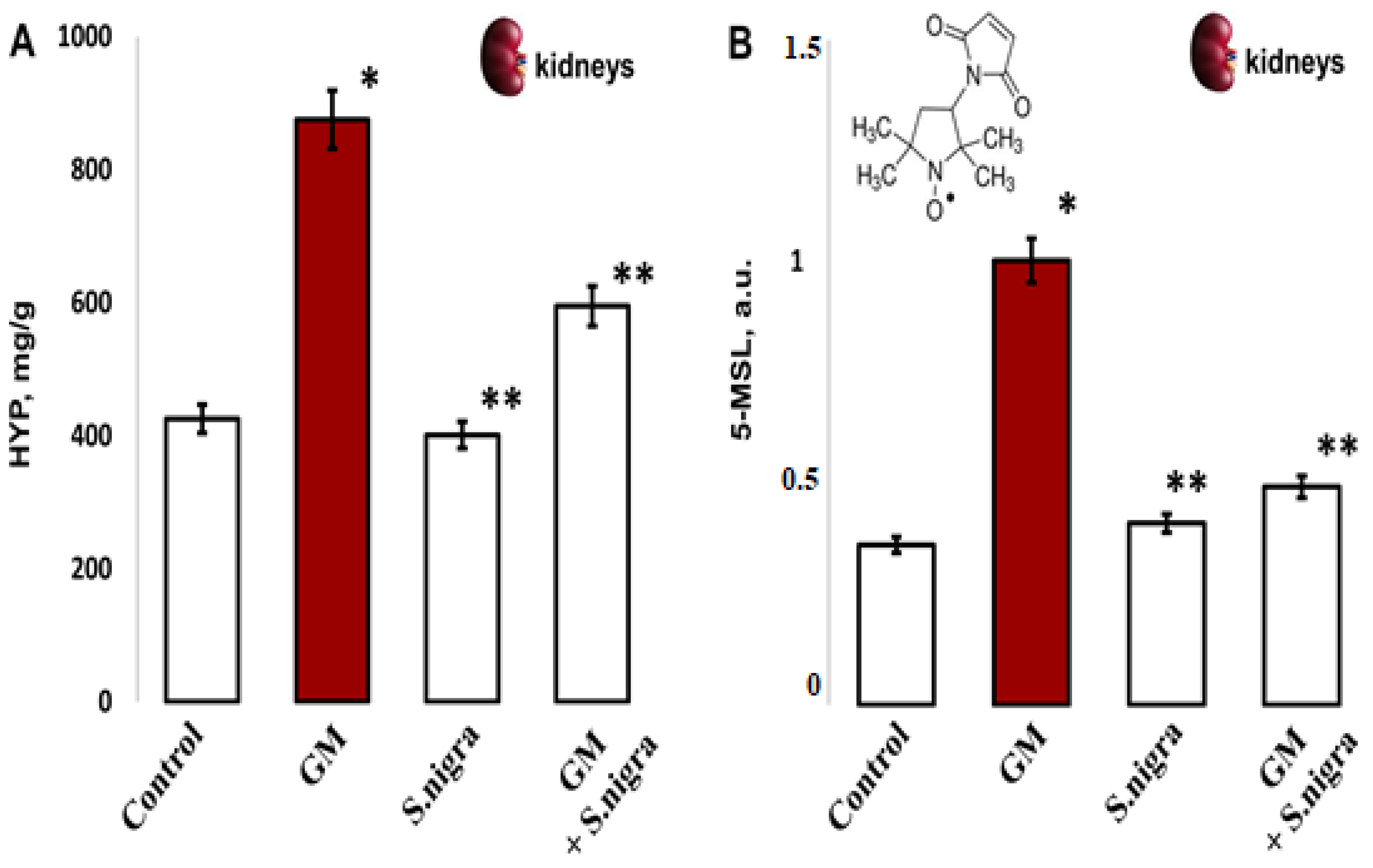
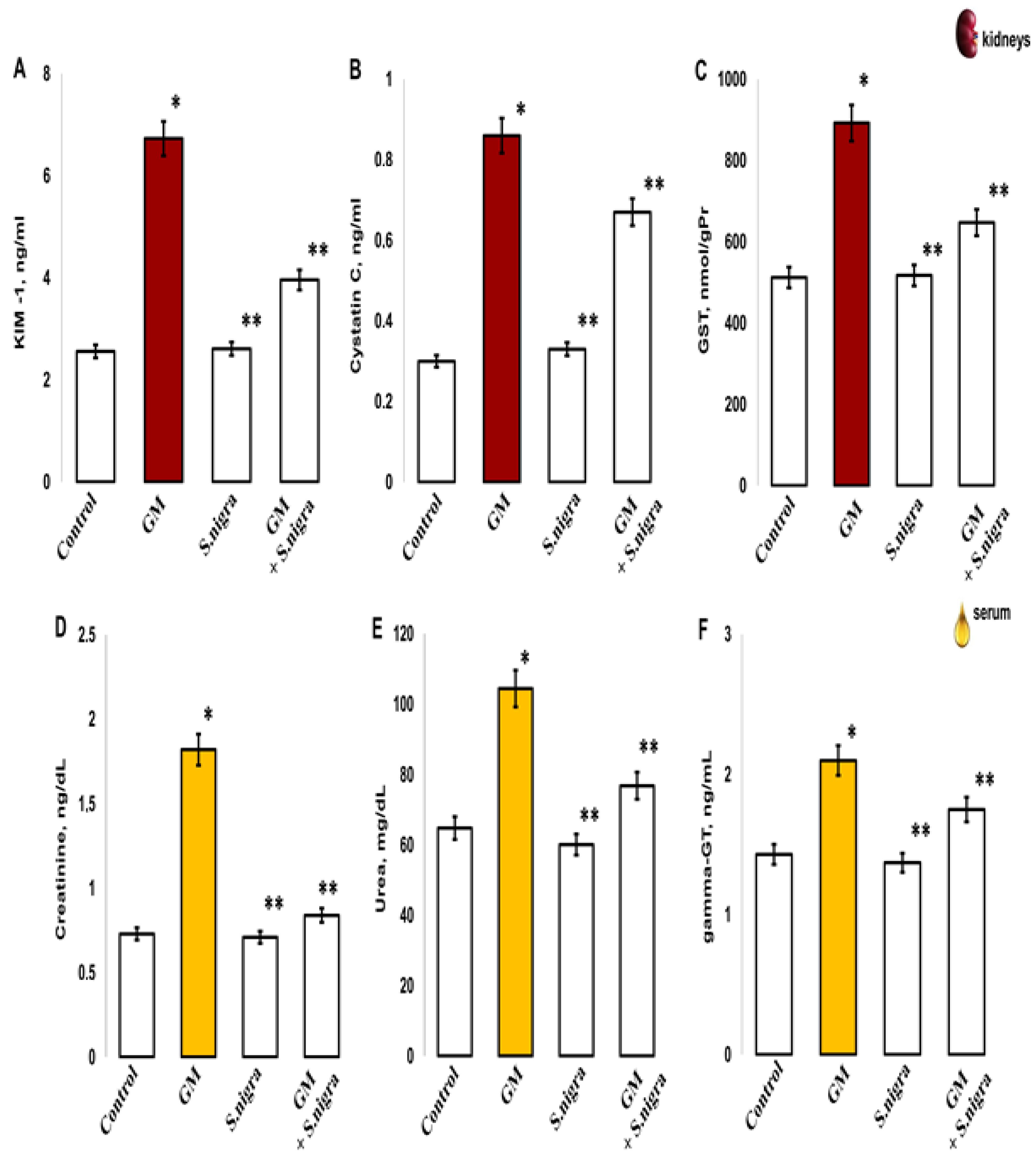
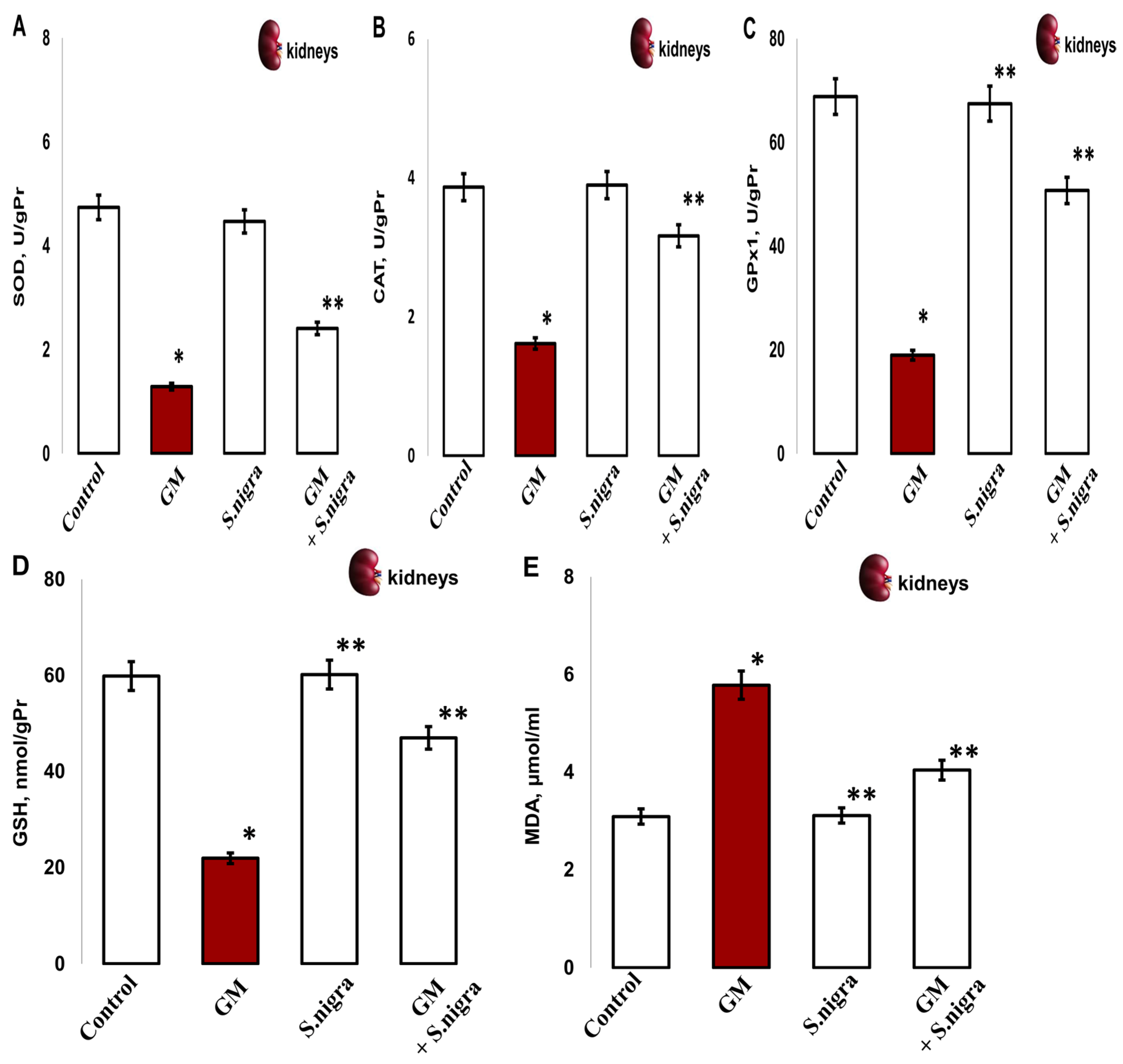
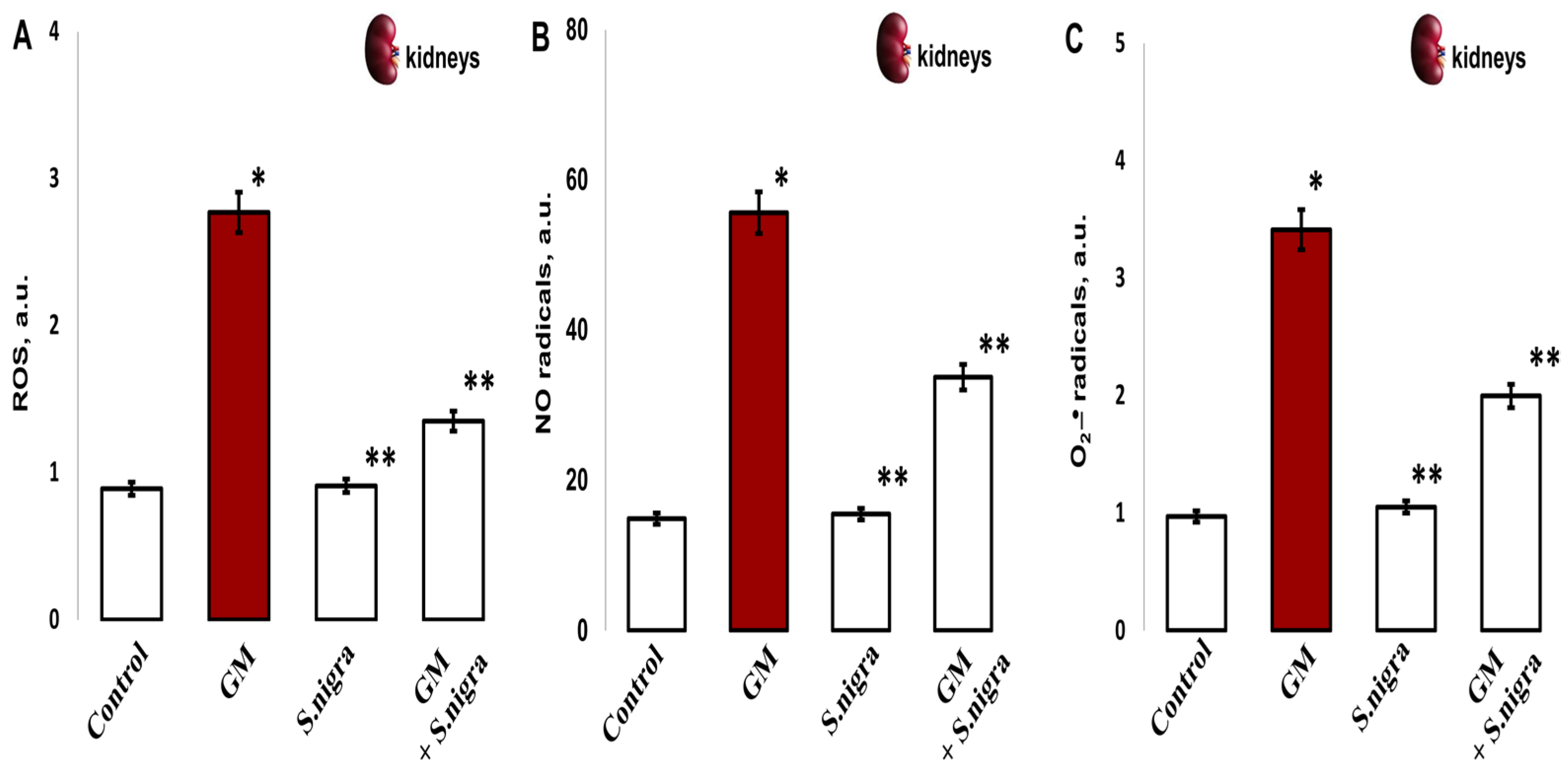
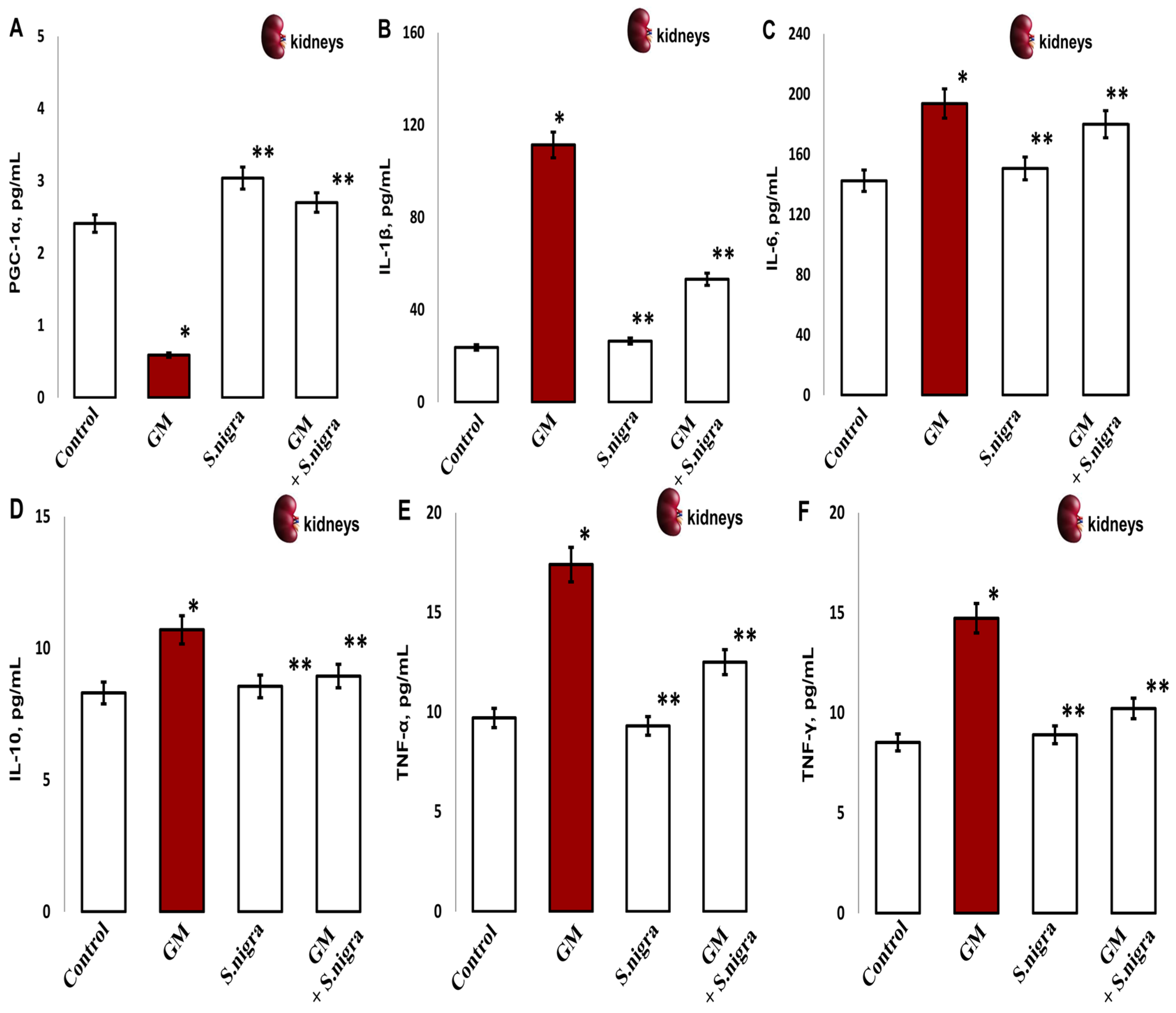
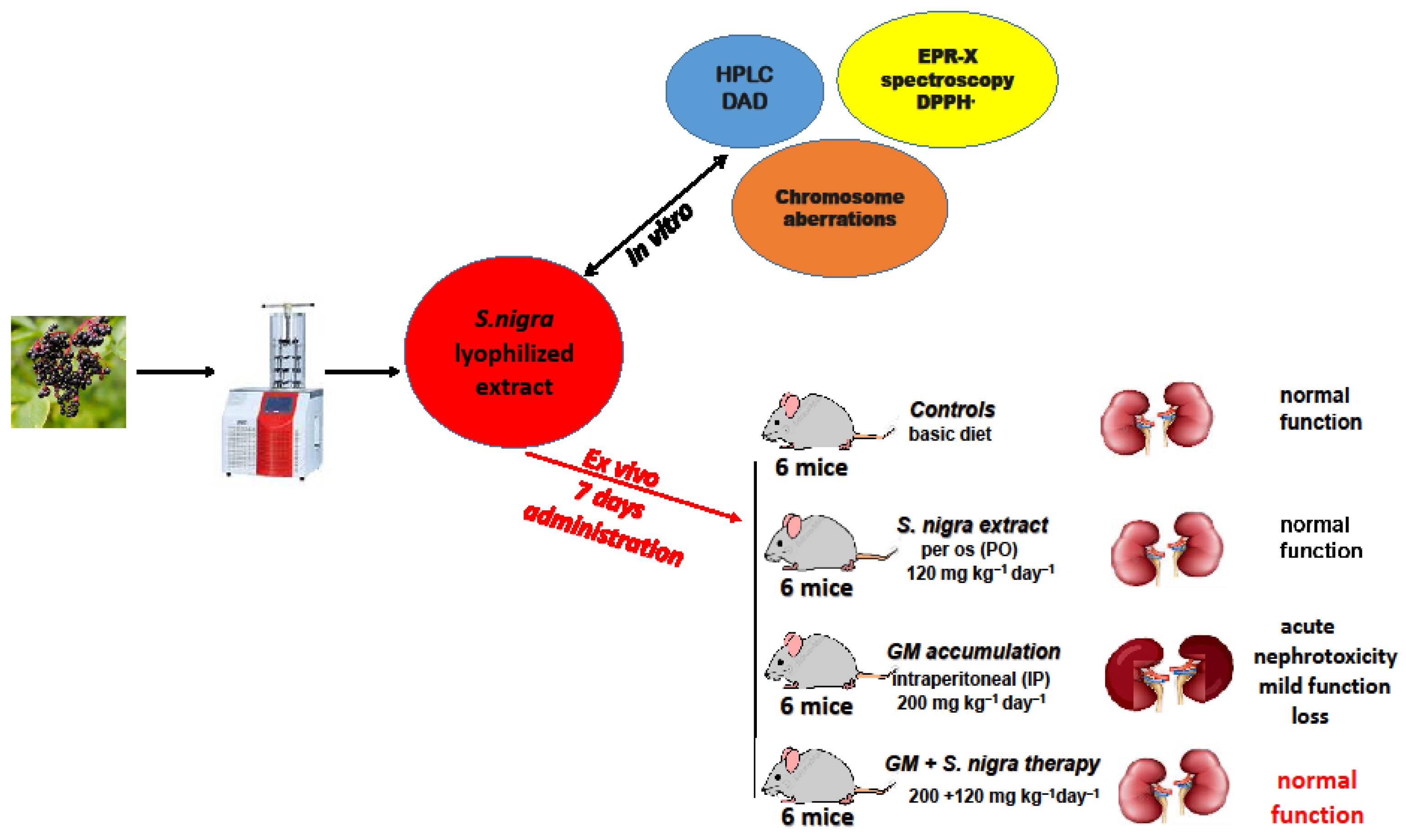
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).