Linezolid Resistance in Staphylococci




Abstract
:1. Introduction

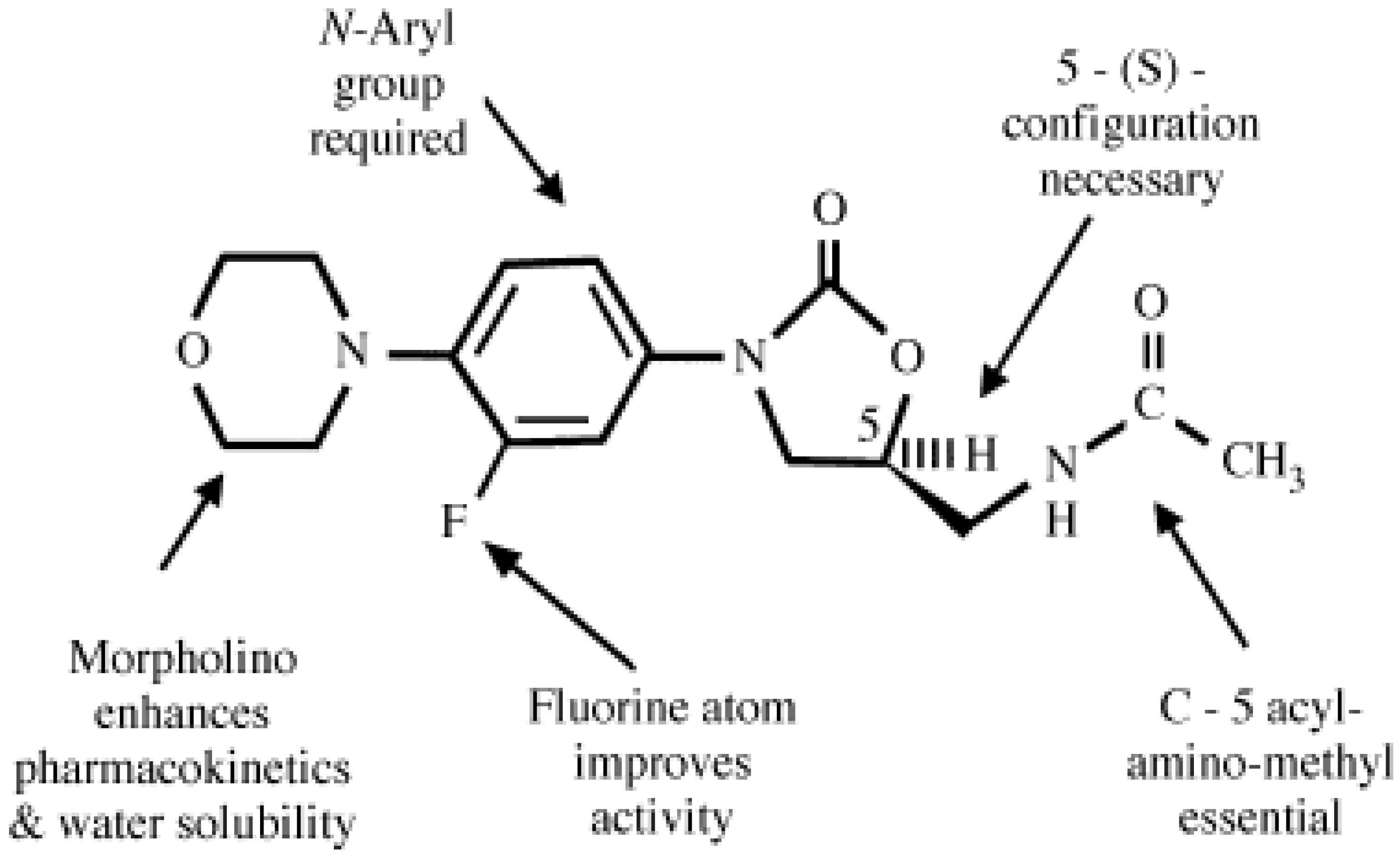
2. Linezolid: Spectrum of in Vitro Activity
3. The Mechanism of Action of Oxazolidinones
4. Mechanisms of Resistance: in General
5. Resistance in Domain V of the 23S rRNA and Related Proteins.

{kind=link}
{kind=link}
| Genetic mechanisms | Site mutations or ribosomal protein mutations* | Microrganisms | References |
|---|---|---|---|
| Mutations in domain V | G2576T | S. aureus | [34] |
| CoNS | [13] | ||
| Viridans streptococci | [35] | ||
| Enterococci | [37] | ||
| S. cohnii | [38] | ||
| S. simulans | Personal data—Stefani et al. ms. in preparation | ||
| S. hominis | Personal data—Stefani et al. ms. in preparation | ||
| G2505A | Enterococci | ||
| G2512T | Enterococci | [39] | |
| G2513T | Enterococci | [39] | |
| C2610G | Enterococci | [39] | |
| G2447T | S. aureus (lab mutant) | [39] | |
| T2500A | S. aureus | [37] | |
| C2192T | S. aureus | [40] | |
| G2447T | S. aureus | [41] | |
| A1743T | S. pneumoniae | [30] | |
| A2503G | S. aureus, S. pneumoniae (lab mutant) | [42] | |
| T2504C | S. aureus | [42] | |
| G2766T | S. aureus | [44] | |
| G2631T | S. epidermidis | [43] | |
| C2543T | S. epidermidis | [43] | |
| C2576T | S. epidermidis | [43] | |
| Mutations in rplD (L4 r-protein) * | 65WR66 and 68KG69 deletions | S. pneumoniae | [45] |
| A202C | S. aureus (lab mutant) | [46] | |
| K68N, L108S, N158S substitutions | S. epidermidis, S. pneumoniae | [43] | |
| Mutations in rplC (L3 r-protein) * | G455A, G463C | S. aureus | [46] |
| A505T, Δser145 | |||
| cfr Methyltransferase | A2503 | S. aureus | |
| S. epidermidis | [49] | ||
| S. sciuri | [52] |
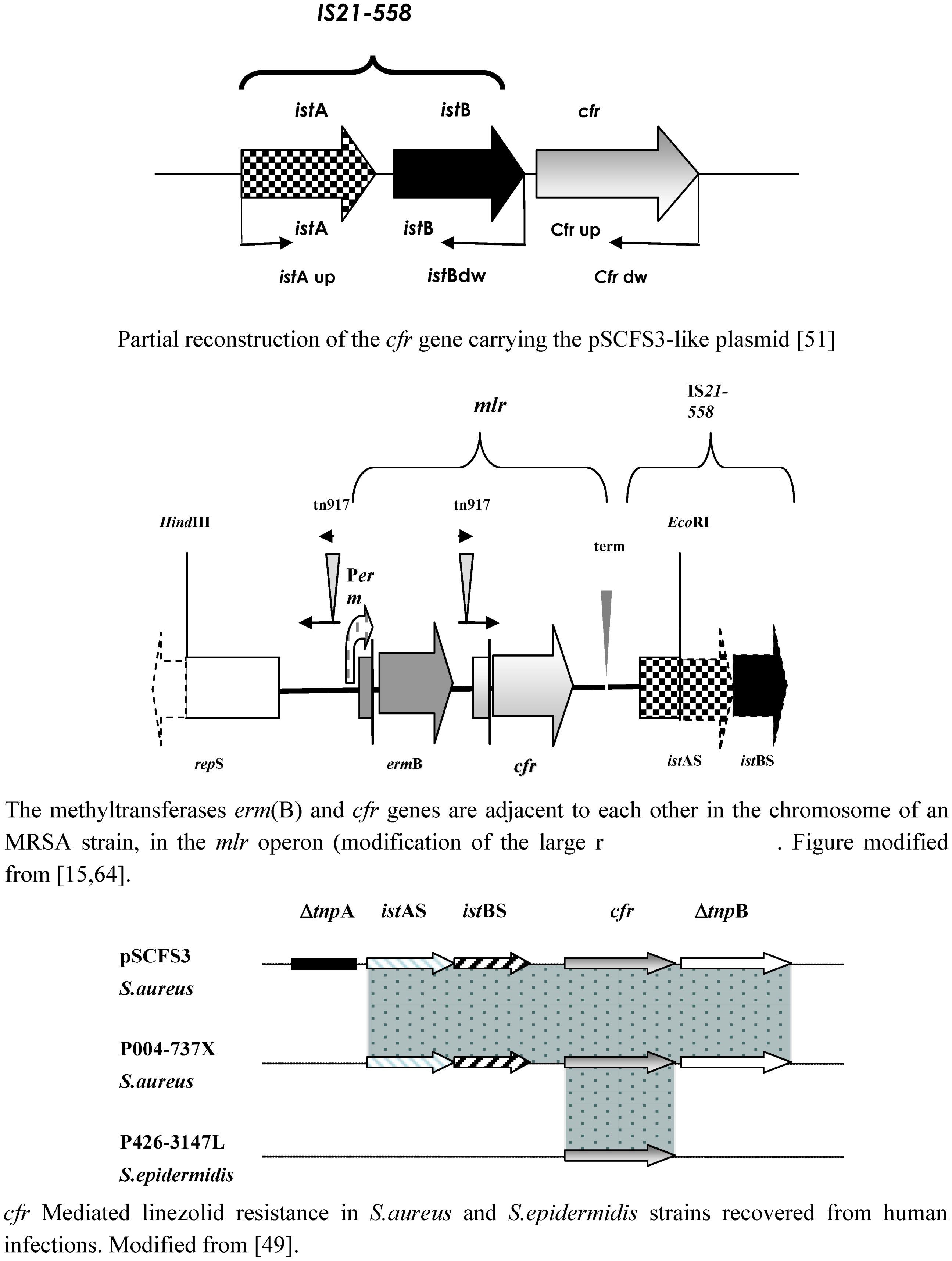
6. The Cfr Mechanism

7. Epidemiological Data on S. aureus and CoNS
8. Problems with in vitro Detection of Resistance
9. Oxazolidinones: the Future
10. Conclusion
Acknowledgements
References
- Ford, C.; Hamel, J.; Stapert, D.; Moerman, J.; Hutchinson, D.; Barbachyn, M.; Zurenko, G. Oxazolidinones: A new class of antimicrobials. Infect. Med. 1999, 16, 435–445. [Google Scholar]
- Livermore, D.M. Linezolid in vitro: Mechanism and antibacterial spectrum. J. Antimicrob. Chemother. 2003, 51 Suppl. 2, ii9–ii16. [Google Scholar] [PubMed]
- FDA talk paper, FDA approves ZYVOX, the first antimicrobial drug in a new class. Food and Drugs Administration, 18 April 2000.
- Clinical and Laboratory Standard Institute, Performance Standard for Antimicrobial Susceptibility Testing; Twenties informational supplement.; 2010; pp. M100–S20.
- EUCAST. Clinical breakpoints. The European Committee on Antimicrobial Susceptibility Testing 2010. Available online: http://eucast.www137.server1.mensemedia.net/clinical_breakpoints/ (accessed on June 18).
- Diekema, D.I.; Jones, R.N. Oxazolidinones: A review. Drugs 2000, 59, 7–16. [Google Scholar]
- Zurenko, G.E.; Yagi, B.H.; Schaadt, R.D.; Allison, J.W.; Kilburn, J.O.; Glickman, S.E.; Hutchinson, D.K.; Barbachyn, M.R.; Brickner, S.J. In vitro activities of U-100592 and U-100766, novel oxazolidinone antibacterial agents. Antimicrob. Agents Chemother. 1996, 40, 839–845. [Google Scholar] [PubMed]
- Buysse, J.; Demyan, W.; Dunyak, D.; Stapert, D.; Hamel, J.; Ford, C. Mutation of the AcrAB antibiotic efflux pump in Escherichia coli confers susceptibility to oxazolidinone antibiotics. In Program and Abstracts of the 36th Interscience Conference on Antimicrobial Agents and Chemotherapy, New Orleans, LA, USA, 15-18 September 1996; American Society for Microbiology: Washington, DC, USA, 1996. Abstract. C-42. [Google Scholar]
- Wallace, R.J.J.; Brown-Elliott, B.A.; Ward, S.C.; Crist, C.J.; Mann, L.B.; Wilson, R.W. Activities of linezolid against rapidly growing mycobacteria. Abstr. ICAAC 2001, 45, 764–767. [Google Scholar]
- Peters, J.; Kondo, K.; Lee, R.; Lin, C.; Inderlied, C. In-vitro activity of oxazolidinones against mycobacterium avium complex. J. Antimicrob. Chemother. 1995, 35, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Vera-Cabrera, L.; Gonzalez, E.; Redon, A.; Ocampo-Candiani, J.; Welsh, O.; Velazquez-Moreno, V.; Choi, S.H.; Molina-Torres, C. In vitro Activities of DA-7157 and DA-7218 against mycobacterium tuberculosis and nocardia brasiliensis. Antimicrob. Agents Chemother. 2006, 50, 3170–3172. [Google Scholar] [PubMed]
- Brown-Elliott, B.A.; Ward, S.C.; Crist, C.J.; Mann, L.B.; Wilson, R.W.; Wallace, R.J.J. In vitro activities of linezolid against multiple nocardia species. Antimicrob. Agents Chemother. 2001, 45, 1295–1297. [Google Scholar] [PubMed]
- Vardakas, K.Z.; Kioumis, I.; Falagas, M.E. Association of pharmacokinetic and pharmacodynamic aspects of linezolid with infection outcome. Curr. Drug Metab. 2009, 10, 2–12. [Google Scholar]
- Potoski, B.A.; Adams, J.; Clarke, L.; Shutt, K.; Linden, P.K.; Baxter, C.; Pasculle, A.W.; Capitano, B.; Peleg, A.Y.; Szabo, D.; Paterson, D.L. Epidemiological profile of linezolid-resistant coagulase-negative staphylococci. Clin. Infect. Dis. 2006, 43, 165–171. [Google Scholar]
- Toh, S.M.; Xiong, L.; Arias, C.A.; Villegas, M.V.; Lolans, K.; Quinn, J.; Mankin, A.S. Acquisition of a natural resistance gene renders a clinical strain of methicillin-resistant Staphylococcus aureus resistant to the synthetic antibiotic linezolid. Mol. Microbiol. 2007, 64, 1506–1514. [Google Scholar]
- Leach, K.L.; Swaney, S.M.; Colca, J.R.; McDonald, W.G.; Blinn, J.R.; Thomasco, L.M.; Gadwood, R.C.; Shinabarger, D.; Xiong, L.; Mankin, A.S. The site of action of oxazolidinone antibiotics in living bacteria and in human mitochondria. Mol. Cell 2007, 26, 393–402. [Google Scholar]
- Ippolito, J.A.; Kanyo, Z.F.; Wang, D.; Franceschi, F.J.; Moore, P.B.; Steitz, T.A.; Duffy, E.M. Crystal structure of the oxazolidinone antibiotic linezolid bound to the 50S ribosomal subunit. J. Med. Chem. 2008, 51, 3353–3356. [Google Scholar]
- Wilson, D.; Schluenzen, F.; Harms, J.; Starosta, A.; Connell, S.; Fucini, P. The oxazolidinone antibiotics perturb the ribosomal peptidyl-transferase center and effect tRNA positioning. Proc. Natl. Acad. Sci. USA 2008, 105, 13339–13344. [Google Scholar]
- Davidovich, C.; Bashan, A.; Yonath, A. Structural basis for cross-resistance to ribosomal PTC antibiotics. Proc. Natl. Acad. Sci. USA 2008, 105, 20665–20670. [Google Scholar]
- Qin, Y.; Polacek, N.; Vesper, O.; Staub, E.; Einfeldt, E.; Wilson, D.N.; Nierhaus, K.H. The highly conserved LepA is a ribosomal elongation factor that back-translocates the ribosome. Cell 2006, 127, 721–733. [Google Scholar]
- Thompson, J.; O'Connor, M.; Mills, J.A.; Dahlberg, A.E. The protein synthesis inhibitors, oxazolidinones and chloramphenicol, cause extensive translational inaccuracy in vivo. J. Mol. Biol. 2002, 322, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C. Update on macrolide-lincosamide-streptogramin, ketolide, and oxazolidinone resistance genes. FEMS Microbiol. Lett. 2008, 282, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, K.; Pakulat, N.; Fleer, S.; Schnaith, A.; Utermöhlen, O.; Krut, O.; Müller, S.; Krönke, M. Subinhibitory concentrations of linezolid reduce Staphylococcus aureus virulence factor expression. Antimicrob. Agents Chemother. 2004, 48, 546–555. [Google Scholar]
- Bressler, A.M.; Zimmer, S.M.; Gilmore, J.L.; Somani, J. Peripheral neuropathy associated with prolonged use of linezolid. Lancet Infect. Dis. 2004, 4, 528–531. [Google Scholar]
- Lee, E.; Burger, S.; Shah, J.; Melton, C.; Mullen, M.; Warren, F.; Press, R. Linezolid-associated toxic optic neuropathy: A report of 2 cases. Clin. Infect. Dis. 2003, 37, 1389–1391. [Google Scholar]
- Nagiec, E.E.; Wu, L.; Swaney, S.M.; Chosay, J.G.; Ross, D.E.; Brieland, J.K.; Leach, K.L. Oxazolidinones inhibit cellular proliferation via inhibition of mitochondrial protein synthesis. Antimicrob. Agents Chemother. 2005, 49, 3896–3902. [Google Scholar]
- Locke, J.B.; Hilgers, M.; Shaw, K.J. Mutations in ribosomal protein L3 are associated with oxazolidinone resistance in staphylococci of clinical origin. Antimicrob. Agents Chemother. 2009, 53, 5275–5278. [Google Scholar]
- Kloss, P.; Xiong, L.; Shinabarger, D.L.; Mankin, A.S. Resistance mutations in 23 S rRNA identify the site of action of the protein synthesis inhibitor linezolid in the ribosomal peptidyl transferase center. J. Mol. Biol. 1999, 294, 93–101. [Google Scholar]
- Toh, S.M.; Mankin, A.S. An indigenous posttranscriptional modification in the ribosomal peptidyl transferase center confers resistance to an array of protein synthesis inhibitors. J. Mol. Biol. 2008, 380, 593–597. [Google Scholar]
- Feng, J.; Lupien, A.; Gingras, H.; Wasserscheid, J.; Dewar, K.; Legare, D.; Ouellette, M. Genome sequencing of linezolid-resistant Streptococcus pneumoniae mutants reveals novel mechanisms of resistance. Genome Res. 2009, 19, 1214–1223. [Google Scholar]
- Franceschi, F.; Kanyo, Z.; Sherer, E.C.; Sutcliffe, J. Macrolide resistance from the ribosome perspective. Curr. Drug Targets Infect. Disord. 2004, 4, 177–191. [Google Scholar]
- Prystowsky, J.; Siddiqui, F.; Chosay, J.; Shinabarger, D.L.; Millichap, J.; Peterson, L.R.; Noskin, G.A. Resistance to linezolid: Characterization of mutations in rRNA and comparison of their occurrences in vancomycin-resistant enterococci. Antimicrob. Agents Chemother. 2001, 45, 2154–2156. [Google Scholar]
- Hill, R.L.; Kearns, A.M.; Nash, J.; North, S.E.; Pike, R.; Newson, T.; Woodford, N.; Calver, R.; Livermore, D.M. Linezolid-resistant ST36 methicillin-resistant Staphylococcus aureus associated with prolonged linezolid treatment in two paediatric cystic fibrosis patients. J. Antimicrob. Chemother. 2010, 65, 442–445. [Google Scholar]
- Tsiodras, S.; Gold, H.S.; Sakoulas, G.; Eliopoulos, G.M.; Wennersten, C.; Venkataraman, L.; Moellering, R.C.; Ferraro, M.J. Linezolid resistance in a clinical isolate of Staphylococcus aureus. Lancet 2001, 358, 207–208. [Google Scholar]
- Pillai, S.K.; Sakoulas, G.; Wennersten, C.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Ferraro, M.J.; Gold, H.S. Linezolid resistance in Staphylococcus aureus: Characterization and stability of resistant phenotype. J. Infect. Dis. 2002, 186, 1603–1607. [Google Scholar]
- Arias, C.A.; Vallejo, M.; Reyes, J.; Panesso, D.; Moreno, J.; Castaneda, E.; Villegas, M.V.; Murray, B.E.; Quinn, J.P. Clinical and microbiological aspects of linezolid resistance mediated by the cfr gene encoding a 23S rRNA methyltransferase. J. Clin. Microbiol. 2008, 46, 892–896. [Google Scholar]
- Meka, V.G.; Pillai, S.K.; Sakoulas, G.; Wennersten, C.; Venkataraman, L.; DeGirolami, P.C.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Gold, H.S. Linezolid resistance in sequential Staphylococcus aureus isolates associated with a T2500A mutation in the 23S rRNA gene and loss of a single copy of rRNA. J. Infect. Dis. 2004, 190, 311–317. [Google Scholar]
- Petinaki, E.; Kanellopoulou, M.; Damani, A.; Foka, A.; Spiliopoulou, I.; Skalmoutsou, N.; Raitsiou, B.; Valakis, K.; Papafragas, E. Linezolid-resistant Staphylococcus cohnii, Greece. Emerg. Infect. Dis. 2009, 15, 116–118. [Google Scholar]
- North, S.E.; Ellington, M.J.; Johnson, A.P.; Livermore, D.M.; Woodford, N. Novel pyrosequencing assays to detect T2500A and other mutations conferring linezolid resistance in Staphylococcus aureus (abstract C2-272). In Program and Abstracts of the 45th Interscience Conference on Antimicrobial Agents and Chemotherapy, Washington Convention Center Washington, DC, USA, 16-19 December 2005; American Society for Microbiology: Washington, DC, USA, 2005; p. 102. [Google Scholar]
- Howe, R.A.; Wootton, M.; Noel, A.R.; Bowker, K.E.; Walsh, T.R.; MacGowan, A.P. Activity of AZD2563, a novel oxazolidinone, against Staphylococcus aureus strains with reduced susceptibility to vancomycin or linezolid. Antimicrob. Agents Chemother. 2003, 47, 3651–3652. [Google Scholar] [CrossRef] [PubMed]
- Swaney, S.M.; Shinabarger, D.L.; Schaadt, R.D.; Bock, J.H.; Slightom, J.L.; Zurenko, G.E. Oxizaolidinone resisitance is associated with a mutation in the peptidyl transferase region of 23S rRNA. In Program and Abstracts of the 38 Interscience Conference on Antimicrobial Agents and Chemotherapy, San Diego, CA USA, 24-27 September 1998; American Society for Microbiology: Washington, DC, USA, 1998; p. 98, Abstract 104. [Google Scholar]
- Livermore, D.M.; Warner, M.; Mushtaq, S.; North, S.; Woodford, N. In vitro activity of the oxazolidinone RWJ-416457 against linezolid-resistant and -susceptible staphylococci and enterococci. Antimicrob. Agents Chemother. 2007, 51, 1112–1114. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.; Reddy, S.P.; Smyth, D.S.; Aguero-Rosenfeld, M.E.; Sakoulas, G.; Robinson, D.A. Polyphyletic emergence of linezolid-resistant staphylococci in the United States. Antimicrob. Agents Chemother. 2010, 54, 742–748. [Google Scholar]
- Livermore, D.M.; Mushtaq, S.; Warner, M.; Woodford, N. Activity of oxazolidinone TR-700 against linezolid-susceptible and -resistant staphylococci and enterococci. J. Antimicrob. Chemother. 2009, 63, 713–715. [Google Scholar]
- Wolter, N.; Smith, A.M.; Farrell, D.J.; Schaffner, W.; Moore, M.; Whitney, C.G.; Jorgensen, J.H.; Klugman, K.P. Novel mechanism of resistance to oxazolidinones, macrolides, and chloramphenicol in ribosomal protein L4 of the pneumococcus. Antimicrob. Agents Chemother. 2005, 49, 3554–3557. [Google Scholar] [PubMed]
- Locke, J.B.; Hilgers, M.; Shaw, K.J. Novel ribosomal mutations in Staphylococcus aureus strains identified through selection with the oxazolidinones linezolid and torezolid (TR-700). Antimicrob. Agents Chemother. 2009, 53, 5265–5274. [Google Scholar]
- Gregory, S.T.; Dahlberg, A.E. Erythromycin resistance mutations in ribosomal proteins L22 and L4 perturb the higher order structure of 23S ribosomal RNA. J. Mol. Biol. 1999, 289, 827–834. [Google Scholar]
- Kehrenberg, C.; Schwarz, S.; Jacobsen, L.; Hansen, L.H.; Vester, B. A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 2005, 57, 1064–1073. [Google Scholar]
- Mendes, R.E.; Deshpande, L.M.; Castanheira, M.; DiPersio, J.; Saubolle, M.A.; Jones, R.N. First report of cfr-mediated resistance to linezolid in human staphylococcal clinical isolates recovered in the United States. Antimicrob. Agents Chemother. 2008, 52, 2244–2246. [Google Scholar]
- Kehrenberg, C.; Aarestrup, F.M.; Schwarz, S. IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr. Antimicrob. Agents Chemother. 2007, 51, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Campanile, F.; Bongiorno, D.; Borbone, S.; Mongelli, G.; Baldi, M.; Provenzani, R.; Stefani, S. Non-mutational cfr-mediated linezolid resistance Staphylococcus epidermidis isolates. the 19th European Congress of Clinical Microbiology and Infectious Diseases, Helsinki, Finland, 16-19 May 2009. Abstr. P929.
- Schwarz, S.; Werckenthin, C.; Kehrenberg, C. Identification of a plasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri. Antimicrob. Agents Chemother. 2000, 44, 2530–2533. [Google Scholar]
- Marshall, S.H.; Donskey, C.J.; Hutton-Thomas, R.; Salata, R.A.; Rice, L.B. Gene dosage and linezolid resistance in Enterococcus faecium and Enterococcus faecalis. Antimicrob. Agents Chemother. 2002, 46, 3334–3336. [Google Scholar]
- Xiong, L.; Kloss, P.; Douthwaite, S.; Andersen, N.M.; Swaney, S.; Shinabarger, D.L.; Mankin, A.S. Oxazolidinone resistance mutations in 23S rRNA of Escherichia coli reveal the central region of domain V as the primary site of drug action. J. Bacteriol. 2000, 182, 5325–5331. [Google Scholar]
- Sander, P.; Belova, L.; Kidan, Y.G.; Pfister, P.; Mankin, A.S.; Bottger, E.C. Ribosomal and non-ribosomal resistance to oxazolidinones: Species-specific idiosyncrasy of ribosomal alterations. Mol. Microbiol. 2002, 46, 1295–1304. [Google Scholar]
- Besier, S.; Ludwig, A.; Zander, J.; Brade, V.; Wichelhaus, T.A. Linezolid resistance in Staphylococcus aureus: Gene dosage effect, stability, fitness costs, and cross-resistances. Antimicrob. Agents Chemother. 2008, 52, 1570–1572. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois-Nicolaos, N.; Kharrat, P.; Butel, M.J.; Doucet-Populaire, F. Fitness cost of linezolid resistance in Enterococcus faecalis. the 18th European Congress of Clinical Microbiology and Infectious Diseases, Barcelona, Spain, 19-22 April 2008. Abstr. O345.
- Mazur, W.; Knob, C.; Fraimow, H. Quantification of 23S rRNA mutations and relative fitness of clinical isolates of linezolid-resistant Enterococcus faecalis. In Program and abstracts of the 42nd Interscience Conference on Antimicrobial Agents and Chemotherapy, San Diego, CA, USA, 27-30 September 2002; American Society for Microbiology: Washington, DC, USA, 2002; p. 75. [Google Scholar]
- Malbruny, B.; Canu, A.; Bozdogan, B.; Fantin, B.; Zarrouk, V.; Dutka-Malen, S.; Feger, C.; Leclercq, R. Resistance to quinupristin-dalfopristin due to mutation of L22 ribosomal protein in Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2200–2207. [Google Scholar]
- Musher, D.M.; Dowell, M.E.; Shortridge, V.D.; Flamm, R.K.; Jorgensen, J.H.; Le Magueres, P.; Krause, K.L. Emergence of macrolide resistance during treatment of pneumococcal pneumonia. N. Engl. J. Med. 2002, 346, 630–631. [Google Scholar]
- Carsenti-Dellamonica, H.; Galimand, M.; Vandenbos, F.; Pradier, C.; Roger, P.M.; Dunais, B.; Sabah, M.; Mancini, G.; Dellamonica, P. In vitro selection of mutants of Streptococcus pneumoniae resistant to macrolides and linezolid: Relationship with susceptibility to penicillin G or macrolides. J. Antimicrob. Chemother. 2005, 56, 633–642. [Google Scholar]
- Jones, R.N.; Fritsche, T.R.; Sader, H.S.; Ross, J.E. LEADER surveillance program results for 2006: an activity and spectrum analysis of linezolid using clinical isolates from the United States (50 medical centers). Diagn. Microbiol. Infect. Dis. 2007, 59, 309–317. [Google Scholar]
- Jones, R.N.; Fritsche, T.R.; Sader, H.S.; Ross, J.E. Zyvox Annual Appraisal of Potency and Spectrum Program Results for 2006: An activity and spectrum analysis of linezolid using clinical isolates from 16 countries. Diagn. Microbiol. Infect. Dis. 2007, 59, 199–209. [Google Scholar]
- Smith, L.K.; Mankin, A.S. Transcriptional and translational control of the mlr operon, which confers resistance to seven classes of protein synthesis inhibitors. Antimicrob. Agents Chemother. 2008, 52, 1703–1712. [Google Scholar]
- Giessing, A.M.; Jensen, S.S.; Rasmussen, A.; Hansen, L.H.; Gondela, A.; Long, K.; Vester, B.; Kirpekar, F. Identification of 8-methyladenosine as the modification catalyzed by the radical SAM methyltransferase Cfr that confers antibiotic resistance in bacteria. RNA 2009, 15, 327–336. [Google Scholar]
- Kehrenberg, C.; Ojo, K.K.; Schwarz, S. Nucleotide sequence and organization of the multiresistance plasmid pSCFS1 from Staphylococcus sciuri. J. Antimicrob. Chemother. 2004, 54, 936–939. [Google Scholar]
- Kehrenberg, C.; Schwarz, S. fexA, a novel Staphylococcus lentus gene encoding resistance to florfenicol and chloramphenicol. Antimicrob. Agents Chemother. 2004, 48, 615–618. [Google Scholar]
- Long, K.S.; Poehlsgaard, J.; Kehrenberg, C.; Schwarz, S.; Vester, B. The Cfr rRNA methyltransferase confers resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics. Antimicrob. Agents Chemother. 2006, 50, 2500–2505. [Google Scholar]
- Kehrenberg, C.; Cuny, C.; Strommenger, B.; Schwarz, S.; Witte, W. Methicillin-resistant and -susceptible Staphylococcus aureus strains of clonal lineages ST398 and ST9 from swine carry the multidrug resistance gene cfr. Antimicrob. Agents Chemother. 2009, 53, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Morales, G.; Picazo, J.J.; Baos, E.; Candel, F.J.; Arribi, A.; Pelaez, B.; Andrade, R.; de la Torre, M.A.; Fereres, J.; Sanchez-Garcia, M. Resistance to linezolid is mediated by the cfr gene in the first report of an outbreak of linezolid-resistant Staphylococcus aureus. Clin. Infect. Dis. 2010, 50, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Cercenado, E.; Marin, M.; Insa, R.; Bouza, E. Emergence of linezolid-resistant Gram-positive clinical isolates due to cfrmethylase gene production associated with G2576T mutation in the 23S rRNA. the 20th European Congress of Clinical Microbiology and Infectious Diseases, Vienna, Austria, 10-13 April 2010. 16 Supplement No. 2, p. S265.
- Spiliopoulou, I.; Damani, A.; Schoina, S.; Liakopoulos, A.; Marangos, M.; Malli, E.; Leonidou, L.; Maroulis, V.; Velissaris, D.; Fligou, F.; Filos, K.; Anastassiou, E.D.; Petinaki, E. Molecular characterization of linezolid-resistant Staphylococcus epidermidis. the 20th European Congress of Clinical Microbiology and Infectious Diseases, Vienna, Austria, 10-13 April 2010. 16 Supplement No. 2, p. S264.
- Seral, C.; Sáenz, Y.; Algarate, S.; Durán, E.; Luque, P.; Rubio-Calvo, C.; Torres, C.; Castillo, F.J. Nosocomial outbreak of methicillin and linezolid-resistant Staphylococcus epidermidis associated with catheter-related bloodstream infections in intensive care unit patients. the 20th European Congress of Clinical Microbiology and Infectious Diseases, Vienna, Austria, 10-13 April 2010. 16 Supplement No. 2, S265 same as 71?.
- Ross, J.E.; Jones, R.; Sader, H.; Uchino, U. Report of linezolid resistance from the Zyvox® Annual Appraisal of Potency and Spectrum Programme (Europe, Latin America, Asia-Pacific). the 20th European Congress of Clinical Microbiology and Infectious Diseases, Vienna, Austria, 10-13 April 2010. 16 Supplement No. 2, S491.
- Baos, E.; Morales, G.; Picazo, J. The resistance mechanism mediated by the cfr gene is predominant in clinical isolates of Staphylococcus epidermidis linezolid-resistant obtained in a Spanish hospital. the 20th European Congress of Clinical Microbiology and Infectious Diseases, Vienna, Austria, 10-13 April 2010. 16 Supplement No. 2, S264.
- Farrell, D.J.; Mendes, R.E.; Ross, J.E.; Jones, R.N. Linezolid surveillance program results for 2008 (LEADER Program for 2008). Diagn. Microbiol. Infect. Dis. 2009, 65, 392–403. [Google Scholar]
- Kehrenberg, C.; Schwarz, S. Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob. Agents Chemother. 2006, 50, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Gemmell, C.G. Susceptibility of a variety of clinical isolates to linezolid: A European inter-country comparison. J. Antimicrob. Chemother. 2001, 48, 47–52. [Google Scholar]
- Tenover, F.C.; Williams, P.P.; Stocker, S.; Thompson, A.; Clark, L.A.; Limbago, B.; Carey, R.B.; Poppe, S.M.; Shinabarger, D.; McGowan, J.E., Jr. Accuracy of six antimicrobial susceptibility methods for testing linezolid against staphylococci and enterococci. J. Clin. Microbiol. 2007, 45, 2917–2922. [Google Scholar]
- Vara Prasad, J. New oxazolidinones. Curr. Opin. Microbiol. 2007, 10(5), 454–460. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, L.; Danese, P.; De Vito, J.; Franceschi, F.; Sutcliffe, J. In vitro activities of the Rx-01 oxazolidinones against hospital and community pathogens. Antimicrob. Agents Chemother. 2008, 52, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, S.; Kosowska-Shick, K.; Appelbaum, P.C.; Verween, G.; Tulkens, P.M.; Van Bambeke, F. Cellular pharmacodynamics of the novel biaryloxazolidinone radezolid: Studies with infected phagocytic and non-phagocytic cells, using Staphylococcus aureus, Staphylococcus epidermidis, Listeria monocytogenes, and Legionella pneumophila. Antimicrob. Agents Chemother. 2010. [Google Scholar]
- Kim, E.; Choi, S.; Im, W.; Rhee, J. Pharmacokinetics of TR-701 (DA-7218): A new oxazolidinone in mice, rats and dogs. the 47th Interscience Conference on Antimicrobial Agents and Chemotherapy, Chicago, IL, USA, 17-20 September 2007; Abstr. F1-1692. American Society for Microbiology: Washington, DC, USA.
- Shaw, K.J.; Poppe, S.; Schaadt, R.; Brown-Driver, V.; Finn, J.; Pillar, C.M.; Shinabarger, D.; Zurenko, G. In vitro activity of TR-700, the antibacterial moiety of the prodrug TR-701, against linezolid-resistant strains. Antimicrob. Agents Chemother. 2008, 52, 4442–4447. [Google Scholar] [PubMed]
- Brown, S.D.; Traczewski, M.M. Comparative in vitro antimicrobial activities of torezolid (TR-700), the active moiety of a new oxazolidinone, torezolid phosphate (TR-701), determination of tentative disk diffusion interpretive criteria, and quality control ranges. Antimicrob. Agents Chemother. 2010, 54, 2063–2069. [Google Scholar] [CrossRef] [PubMed]
- Foleno, B.D.; Abbanat, D.; Goldschmidt, R.M.; Flamm, R.K.; Paget, S.D.; Webb, G.C.; Wira, E.; Macielag, M.J.; Bush, K. In vitro antibacterial activity of the pyrrolopyrazolyl-substituted oxazolidinone RWJ-416457. Antimicrob. Agents Chemother. 2007, 51, 361–365. [Google Scholar] [PubMed]
- Hilliard, J.J.; Fernandez, J.; Melton, J.; Macielag, M.J.; Goldschmidt, R.; Bush, K.; Abbanat, D. In vivo activity of the pyrrolopyrazolyl-substituted oxazolidinone RWJ-416457. Antimicrob. Agents Chemother. 2009, 53, 2028–2033. [Google Scholar] [PubMed]
- Skripkin, E.; McConnell, T.S.; De Vito, J.; Lawrence, L.; Ippolito, J.A.; Duffy, E.M.; Sutcliffe, J.; Franceschi, F. R chi-01, a new family of oxazolidinones that overcome ribosome-based linezolid resistance. Antimicrob. Agents Chemother. 2008, 52, 3550–3557. [Google Scholar]
- Sood, R.; Rao, M.; Singhal, S.; Rattan, A. Activity of RBx 7644 and RBx 8700, new investigational oxazolidinones, against Mycobacterium tuberculosis infected murine macrophages. Int. J. Antimicrob. Agents 2005, 25, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Bhadauriya, T.; Rao, M.; Gautam, R.; Malhotra, S.; Barman, T.K.; Upadhyay, D.J.; Rattan, A. Antimycobacterial activities of oxazolidinones: A review. Infect. Disord. Drug Targets 2006, 6, 343–354. [Google Scholar]
- Ednie, L.M.; Rattan, A.; Jacobs, M.R.; Appelbaum, P.C. Antianaerobe activity of RBX 7644 (ranbezolid), a new oxazolidinone, compared with those of eight other agents. Antimicrob. Agents Chemother. 2003, 47, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stefani, S.; Bongiorno, D.; Mongelli, G.; Campanile, F. Linezolid Resistance in Staphylococci. Pharmaceuticals 2010, 3, 1988-2006. https://doi.org/10.3390/ph3071988
Stefani S, Bongiorno D, Mongelli G, Campanile F. Linezolid Resistance in Staphylococci. Pharmaceuticals. 2010; 3(7):1988-2006. https://doi.org/10.3390/ph3071988
Chicago/Turabian StyleStefani, Stefania, Dafne Bongiorno, Gino Mongelli, and Floriana Campanile. 2010. "Linezolid Resistance in Staphylococci" Pharmaceuticals 3, no. 7: 1988-2006. https://doi.org/10.3390/ph3071988
APA StyleStefani, S., Bongiorno, D., Mongelli, G., & Campanile, F. (2010). Linezolid Resistance in Staphylococci. Pharmaceuticals, 3(7), 1988-2006. https://doi.org/10.3390/ph3071988