
Advanced Glycation End Products Increase Salivary Gland Hypofunction in d-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
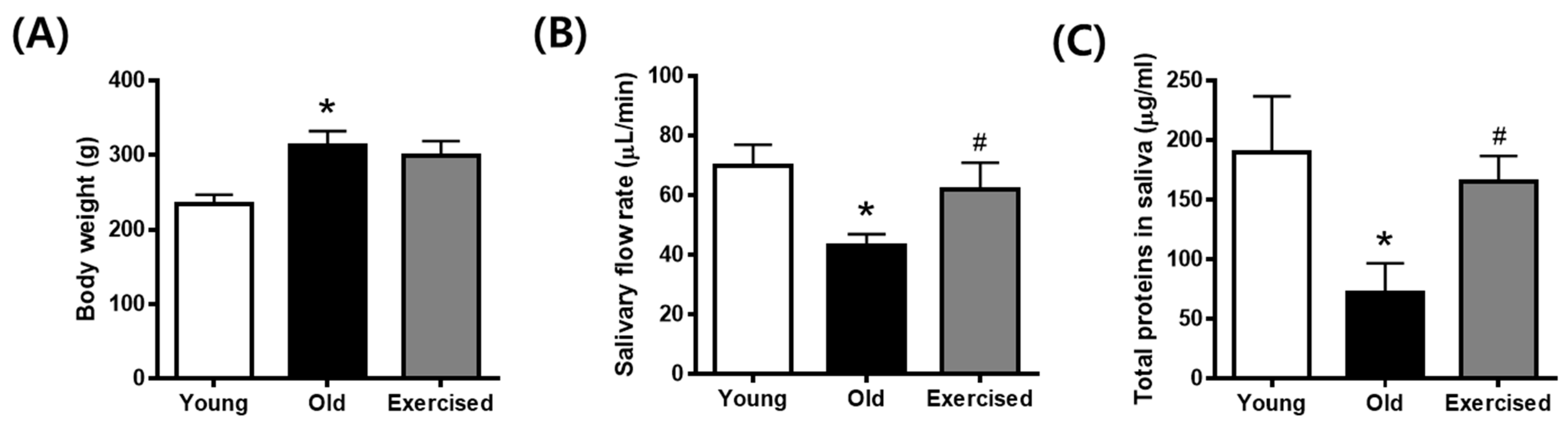
2.1. Physical Exercise Improves Hyposalivation in d-Galactose-Induced Aging Rats
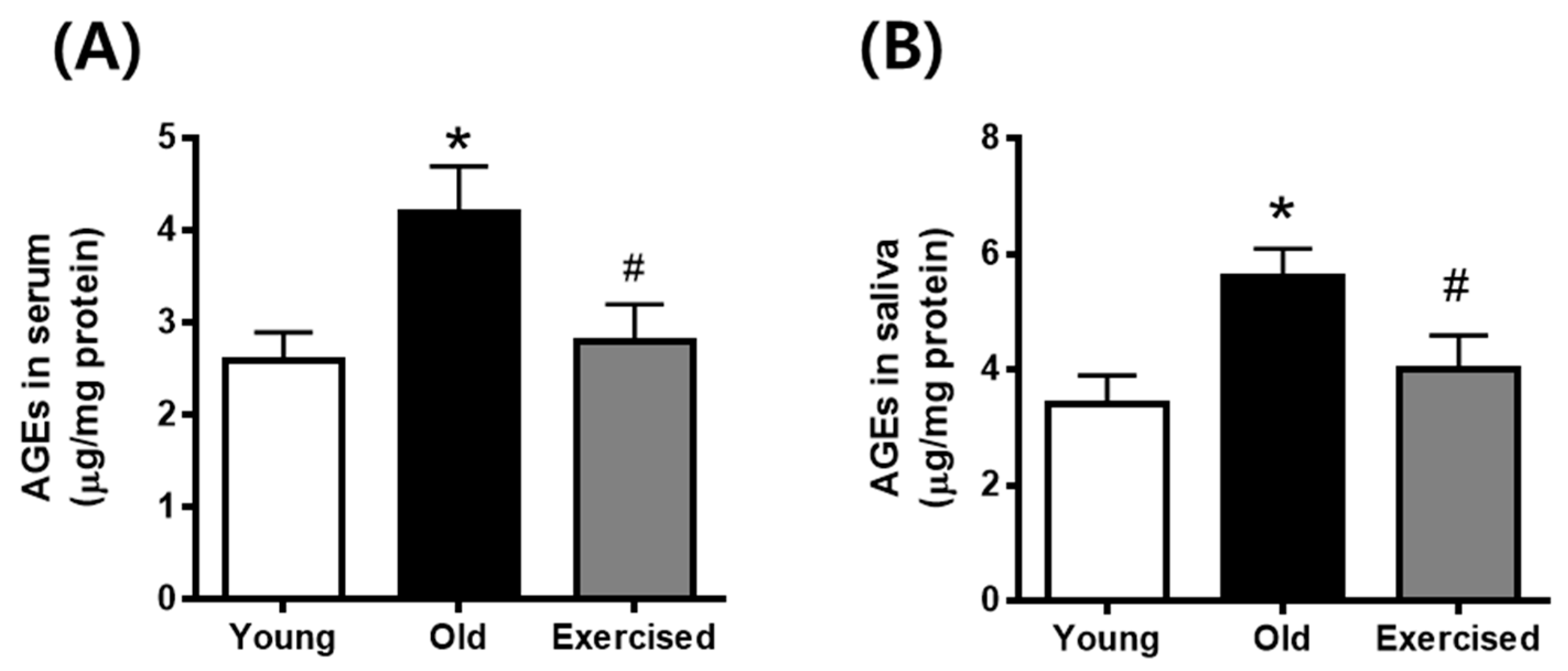
2.2. Physical Exercise Decreases the Levels of Circulating AGEs in Serum and Secreted AGEs in Saliva
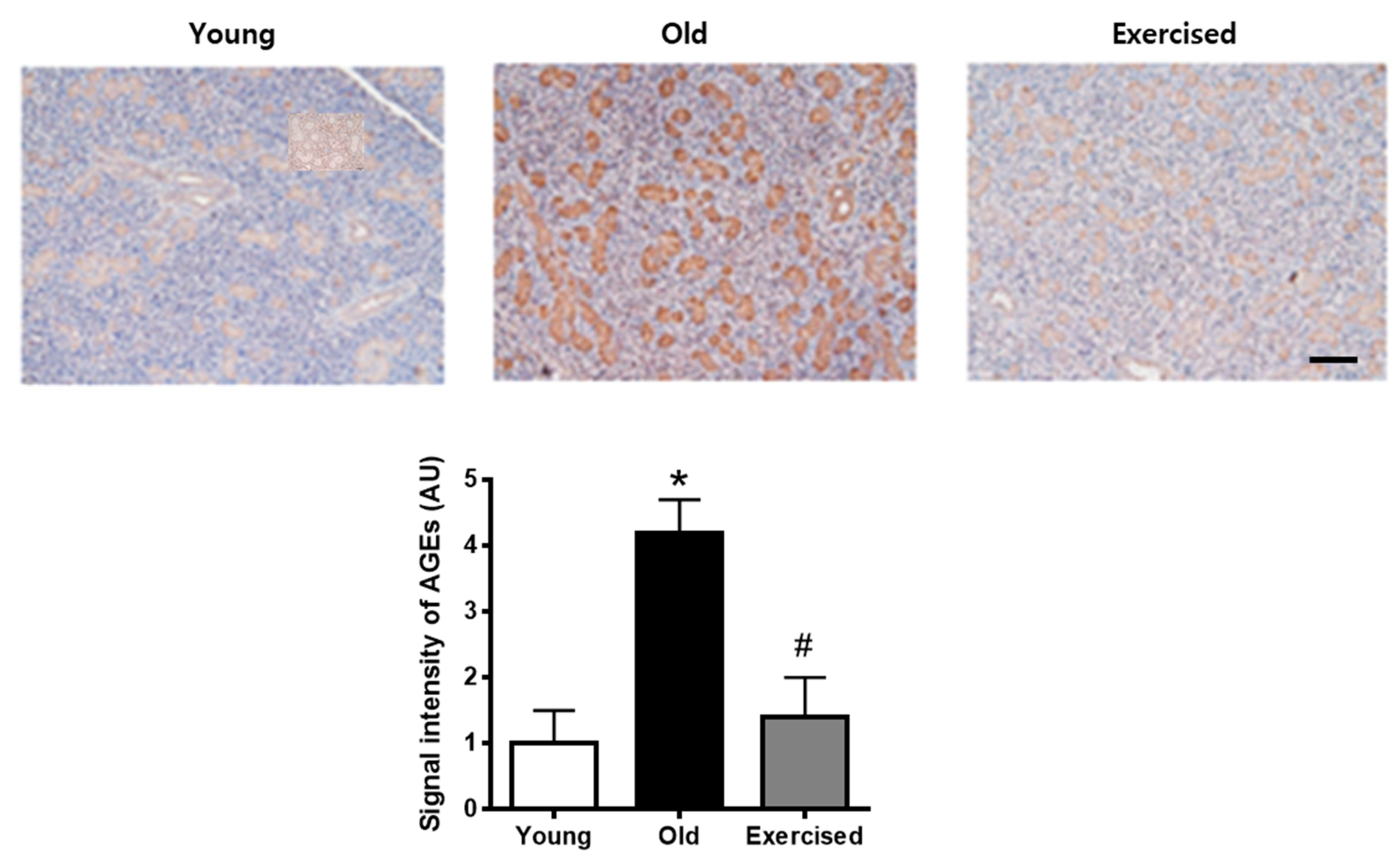
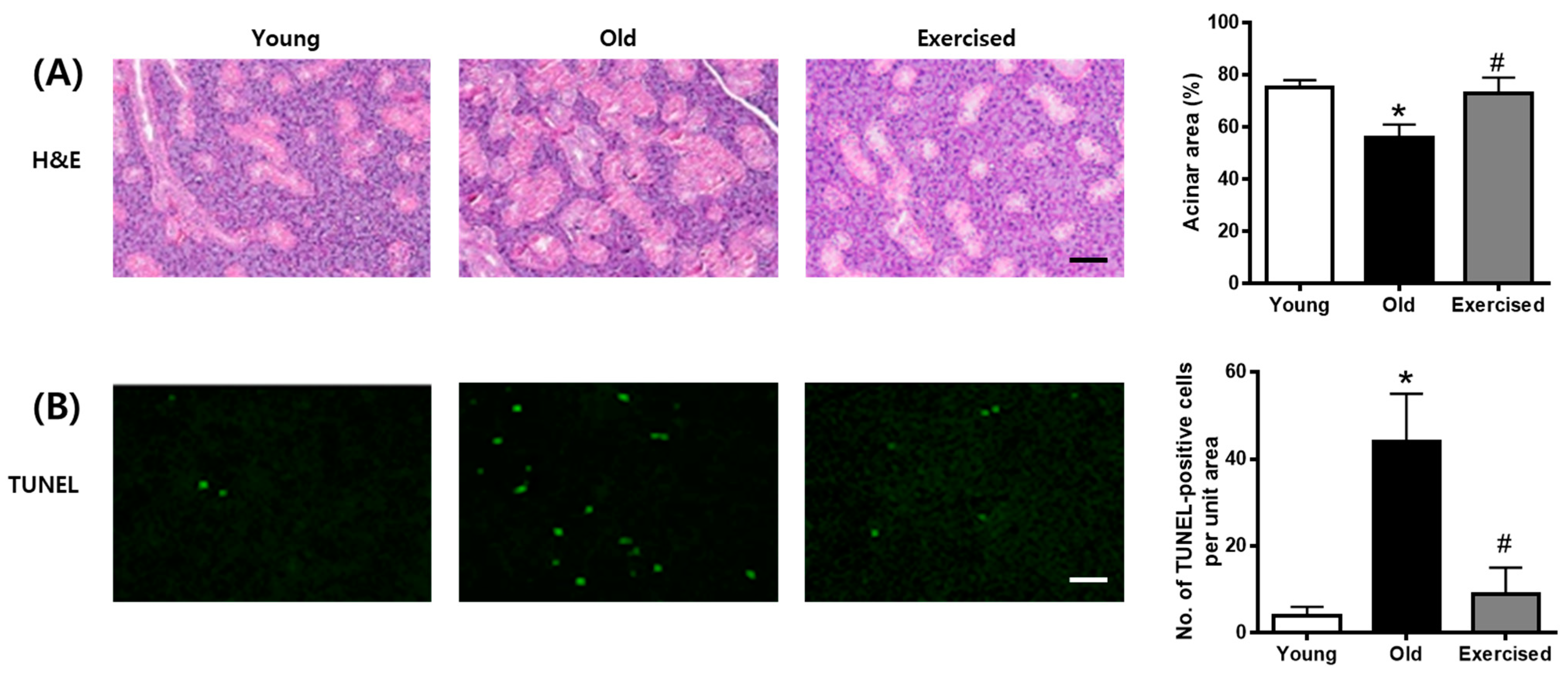
2.3. Physical Exercise Decreases the Accumulation of AGEs in the Salivary Gland
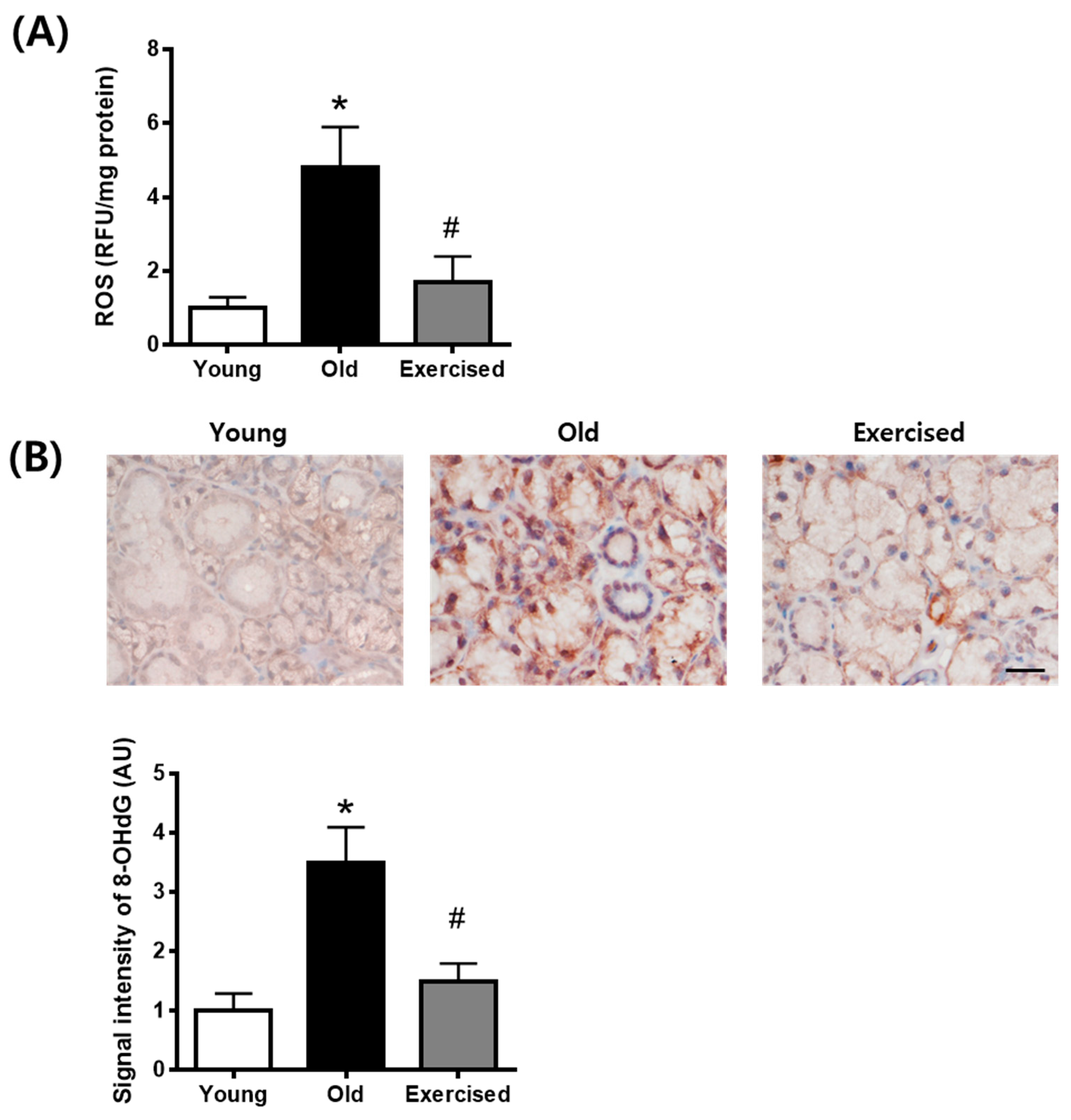
2.4. Physical Exercise Decreases the Generation of ROS in the Salivary Gland
2.5. Physical Exercise Inhibits Apoptosis in the Salivary Glands of d-Galactose-Induced Aging Rats
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Quantification of AGEs in Saliva and Blood
4.3. Oxidative Stress Assay in Salivary Gland
4.4. Histopathological Analysis
4.5. Apoptosis Analysis
4.6. Immunohistochemistry
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- López-Pintor, R.M.; Casañas, E.; González-Serrano, J.; Serrano, J.; Ramírez, L.; de Arriba, L.; Hernández, G. Xerostomia, hyposalivation, and salivary flow in diabetes patients. J. Diabetes Res. 2016, 2016, 4372852. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.K.J.; Chatra, L.; Shenai, P.; Veena, K.; Prabhu, R.V.; Kushraj, T.; Shetty, P.; Hameed, S. Xerostomia: Few dry facts about dry mouth. Arch. Med. Health Sci. 2014, 2, 190. [Google Scholar] [CrossRef]
- Mason, A.L.; Xu, L.; Guo, L.; Garry, R.F. Retroviruses in autoimmune liver disease: Genetic or environmental agents? Arch. Immunol. Et Ther. Exp. 1999, 47, 289–297. [Google Scholar]
- Saleh, J.; Figueiredo, M.A.Z.; Cherubini, K.; Salum, F.G. Salivary hypofunction: An update on aetiology, diagnosis and therapeutics. Arch. Oral Biol. 2015, 60, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, V.; Nix, P. Managing the patient presenting with xerostomia: A review. Int. J. Clin. Pract. 2010, 64, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.Y.; Cooper, M.E. Clinical review: The role of advanced glycation end products in progression and complications of diabetes. J. Clin. Endocrinol. Metab. 2008, 93, 1143–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessier, F.J. The Maillard reaction in the human body. The main discoveries and factors that affect glycation. Pathol. Biol. 2010, 58, 214–219. [Google Scholar] [CrossRef]
- Brownlee, M.; Cerami, A.; Vlassara, H. Advanced glycosylation end products in tissue and the biochemical basis of diabetic complications. N. Engl. J. Med. 1988, 318, 1315–1321. [Google Scholar]
- Lu, J.; Wu, D.M.; Zheng, Y.L.; Hu, B.; Zhang, Z.F.; Ye, Q.; Liu, C.M.; Shan, Q.; Wang, Y.J. Ursolic acid attenuates D-galactose-induced inflammatory response in mouse prefrontal cortex through inhibiting AGEs/RAGE/NF-kappaB pathway activation. Cereb. Cortex 2010, 20, 2540–2548. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Chen, M.; Forrester, J.V. Para-inflammation in the aging retina. Prog. Retin. Eye Res. 2009, 28, 348–368. [Google Scholar] [CrossRef]
- Ilea, A.; Babtan, A.M.; Bosca, B.A.; Crisan, M.; Petrescu, N.B.; Collino, M.; Sainz, R.M.; Gerlach, J.Q.; Campian, R.S. Advanced glycation end products (AGEs) in oral pathology. Arch. Oral Biol. 2018, 93, 22–30. [Google Scholar] [CrossRef]
- Katz, J.; Stavropoulos, F.; Bhattacharyya, I.; Stewart, C.; Perez, F.M.; Caudle, R.M. Receptor of advanced glycation end product (RAGE) expression in the minor salivary glands of patients with Sjogren’s syndrome: A preliminary study. Scand. J. Rheumatol. 2004, 33, 174–178. [Google Scholar] [CrossRef]
- Sell, D.R.; Lane, M.A.; Johnson, W.A.; Masoro, E.J.; Mock, O.B.; Reiser, K.M.; Fogarty, J.F.; Cutler, R.G.; Ingram, D.K.; Roth, G.S.; et al. Longevity and the genetic determination of collagen glycoxidation kinetics in mammalian senescence. Proc. Natl. Acad. Sci. USA 1996, 93, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Asghar, M.; George, L.; Lokhandwala, M.F. Exercise decreases oxidative stress and inflammation and restores renal dopamine D1 receptor function in old rats. Am. J. Physiol. Ren. Physiol. 2007, 293, F914–F919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, A.; Gomez, C.; Lopez-Cepero, J.M.; Boveris, A. Beneficial effects of moderate exercise on mice aging: Survival, behavior, oxidative stress, and mitochondrial electron transfer. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R505–R511. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Cabrera, M.C.; Domenech, E.; Vina, J. Moderate exercise is an antioxidant: Upregulation of antioxidant genes by training. Free Radic. Biol. Med. 2008, 44, 126–131. [Google Scholar] [CrossRef]
- Ferreira, R.O.; Correa, M.G.; Magno, M.B.; Almeida, A.; Fagundes, N.C.F.; Rosing, C.K.; Maia, L.C.; Lima, R.R. Physical activity reduces the prevalence of periodontal disease: Systematic review and meta-analysis. Front. Physiol. 2019, 10, 234. [Google Scholar] [CrossRef]
- Ligtenberg, A.J.; Brand, H.S.; van den Keijbus, P.A.; Veerman, E.C. The effect of physical exercise on salivary secretion of MUC5B, amylase and lysozyme. Arch. Oral. Biol. 2015, 60, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Mealey, B.L.; Ocampo, G.L. Diabetes mellitus and periodontal disease. Periodontology 2000 2007, 44, 127–153. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.; Hayes, C.; Taylor, G.W. Glycemic control of type 2 diabetes and severe periodontal disease in the US adult population. Community Dent. Oral Epidemiol. 2002, 30, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Matsuno, T.; Omata, K.; Satoh, T. Relationship between hyposalivation and oxidative stress in aging mice. J. Clin. Biochem. Nutr. 2017, 61, 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, W.B.; Zhang, B.L. Biochemical changes in D-galactose induced subacute toxicity and mimetic aging in mice. Chinese J Pharm Toxicol 1990, 4, 309–310. [Google Scholar]
- Gong, G.Q.; Xu, F.B. Study of aging model in mice. J. China Pharm. Univ. 1991, 22, 101–103. [Google Scholar]
- Li, W.B.; Wei, F.; Fan, M.; Zhang, J.L.; Zhang, B.L.; Ma, X.C.; Yang, W.P.; Wei, W. Mimetic brain aging effect induced by D-galactose in mice. Chin. J. Pharm. Toxicol. 1995, 9, 93–95. [Google Scholar]
- Vlassara, H.; Bucala, R.; Striker, L. Pathogenic effects of advanced glycosylation: Biochemical, biologic, and clinical implications for diabetes and aging. Lab. Invest. 1994, 70, 138–151. [Google Scholar] [PubMed]
- Song, X.; Bao, M.; Li, D.; Li, Y.M. Advanced glycation in D-galactose induced mouse aging model. Mech. Ageing Dev. 1999, 108, 239–251. [Google Scholar] [CrossRef]
- Brownlee, M. Advanced protein glycosylation in diabetes and aging. Annu. Rev. Med. 1995, 46, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Pokupec, R.; Kalauz, M.; Turk, N.; Turk, Z. Advanced glycation endproducts in human diabetic and non-diabetic cataractous lenses. Graefes Arch. Clin. Exp. Ophthalmol. 2003, 241, 378–384. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamamoto, H. Interaction of receptor for advanced glycation end products with advanced oxidation protein products induces podocyte injury. Kidney Int. 2012, 82, 733–735. [Google Scholar] [CrossRef] [Green Version]
- Hiona, A.; Leeuwenburgh, C. The role of mitochondrial DNA mutations in aging and sarcopenia: Implications for the mitochondrial vicious cycle theory of aging. Exp. Gerontol. 2008, 43, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Cuzzocrea, S.; McDonald, M.C.; Filipe, H.M.; Costantino, G.; Mazzon, E.; Santagati, S.; Caputi, A.P.; Thiemermann, C. Effects of tempol, a membrane-permeable radical scavenger, in a rodent model of carrageenan-induced pleurisy. Eur. J. Pharm. 2000, 390, 209–222. [Google Scholar] [CrossRef]
- Pechter, U.; Maaroos, J.; Mesikepp, S.; Veraksits, A.; Ots, M. Regular low-intensity aquatic exercise improves cardio-respiratory functional capacity and reduces proteinuria in chronic renal failure patients. Nephrol. Dial. Transpl. 2003, 18, 624–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moinuddin, I.; Leehey, D.J. A comparison of aerobic exercise and resistance training in patients with and without chronic kidney disease. Adv. Chronic Kidney Dis. 2008, 15, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Park, S.; Kim, J. The role of glycation in the pathogenesis of aging and its prevention through herbal products and physical exercise. J. Exerc. Nutr. Biochem. 2017, 21, 55–61. [Google Scholar] [CrossRef]
- Kim, C.S.; Park, S.; Chun, Y.; Song, W.; Kim, H.J.; Kim, J. Treadmill exercise attenuates retinal oxidative stress in naturally-aged mice: An immunohistochemical study. Int. J. Mol. Sci. 2015, 16, 21008–21020. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kim, C.S.; Lee, J.; Suk Kim, J.; Kim, J. Effect of regular exercise on the histochemical changes of d-galactose-induced oxidative renal injury in high-fat diet-fed rats. Acta Histochem. Cytochem. 2013, 46, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boor, P.; Celec, P.; Behuliak, M.; Grancic, P.; Kebis, A.; Kukan, M.; Pronayova, N.; Liptaj, T.; Ostendorf, T.; Sebekova, K. Regular moderate exercise reduces advanced glycation and ameliorates early diabetic nephropathy in obese Zucker rats. Metabolism 2009, 58, 1669–1677. [Google Scholar] [CrossRef]
- Stephens, L.C.; Schultheiss, T.E.; Price, R.E.; Ang, K.K.; Peters, L.J. Radiation apoptosis of serous acinar cells of salivary and lacrimal glands. Cancer 1991, 67, 1539–1543. [Google Scholar] [CrossRef]
- Mahali, S.; Raviprakash, N.; Raghavendra, P.B.; Manna, S.K. Advanced glycation end products (AGEs) induce apoptosis via a novel pathway: Involvement of Ca2+ mediated by interleukin-8 protein. J. Biol. Chem. 2011, 286, 34903–34913. [Google Scholar] [CrossRef] [Green Version]
- Li, D.X.; Deng, T.Z.; Lv, J.; Ke, J. Advanced glycation end products (AGEs) and their receptor (RAGE) induce apoptosis of periodontal ligament fibroblasts. Braz. J. Med. Biol. Res. 2014, 47, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hu, X.; Luo, J.; Li, L.; Chen, X.; Huang, R.; Pei, Z. Physical exercise improves functional recovery through mitigation of autophagy, attenuation of apoptosis and enhancement of neurogenesis after MCAO in rats. BMC Neurosci. 2013, 14, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, H.B. Effects of aging and exercise training on apoptosis in the heart. J. Exerc. Rehabil. 2013, 9, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.L.; Mouisel, E.; Roblot, N.; Melki, J. Inter- and intrastrain variation in mouse critical running speed. J. Appl. Physiol. 2005, 98, 1258–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, W.K.; Park, S.-B.; Kim, H.R.; Ryu, H.Y.; Kim, Y.H.; Kim, J. Advanced Glycation End Products Increase Salivary Gland Hypofunction in d-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise. Curr. Issues Mol. Biol. 2021, 43, 2059-2067. https://doi.org/10.3390/cimb43030142
Jung WK, Park S-B, Kim HR, Ryu HY, Kim YH, Kim J. Advanced Glycation End Products Increase Salivary Gland Hypofunction in d-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise. Current Issues in Molecular Biology. 2021; 43(3):2059-2067. https://doi.org/10.3390/cimb43030142
Chicago/Turabian StyleJung, Woo Kwon, Su-Bin Park, Hyung Rae Kim, Hwa Young Ryu, Yong Hwan Kim, and Junghyun Kim. 2021. "Advanced Glycation End Products Increase Salivary Gland Hypofunction in d-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise" Current Issues in Molecular Biology 43, no. 3: 2059-2067. https://doi.org/10.3390/cimb43030142
APA StyleJung, W. K., Park, S.-B., Kim, H. R., Ryu, H. Y., Kim, Y. H., & Kim, J. (2021). Advanced Glycation End Products Increase Salivary Gland Hypofunction in d-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise. Current Issues in Molecular Biology, 43(3), 2059-2067. https://doi.org/10.3390/cimb43030142