
Exploring and Identifying Candidate Genes and Genomic Regions Related to Economically Important Traits in Hanwoo Cattle

Abstract
:1. Introduction
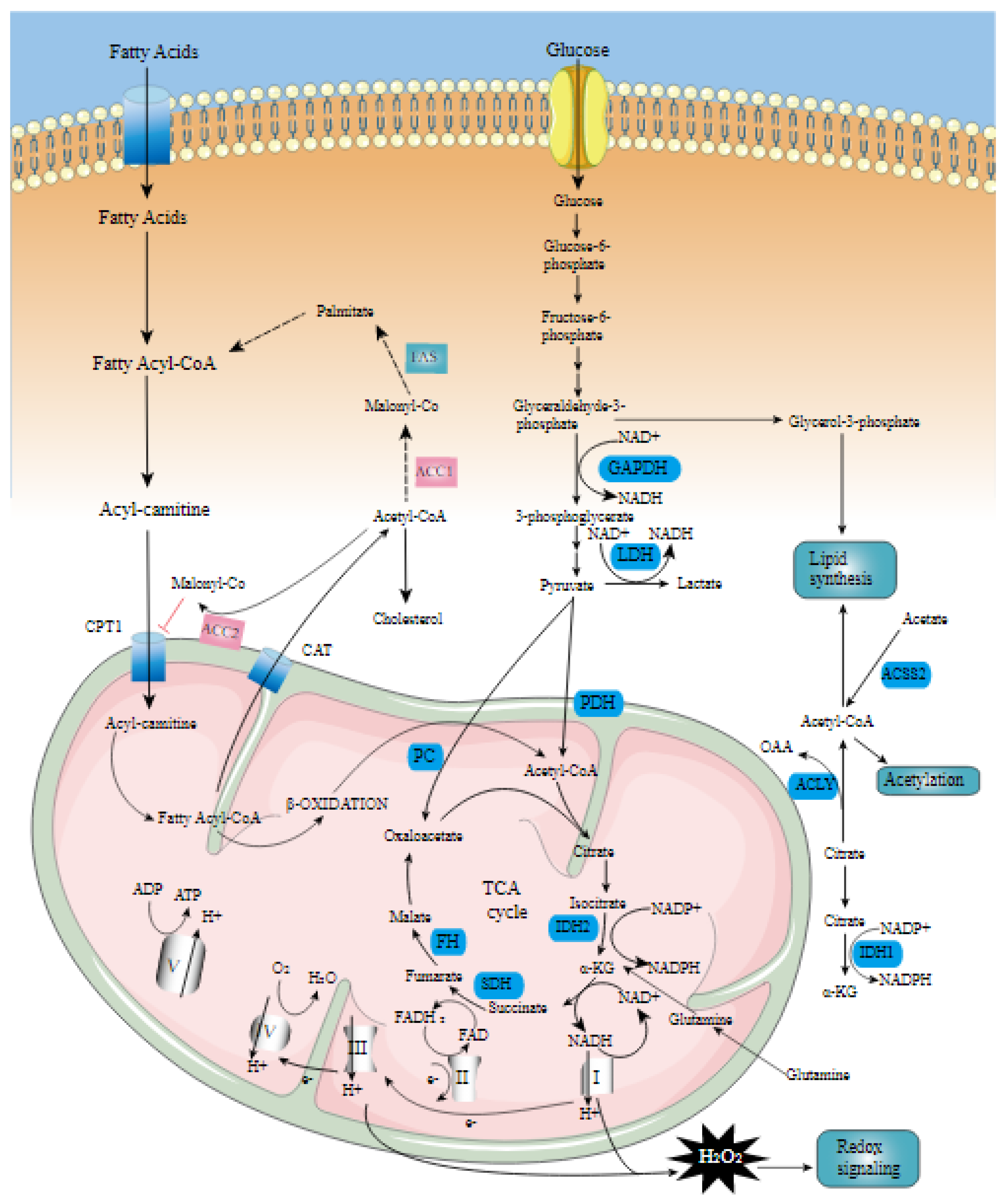
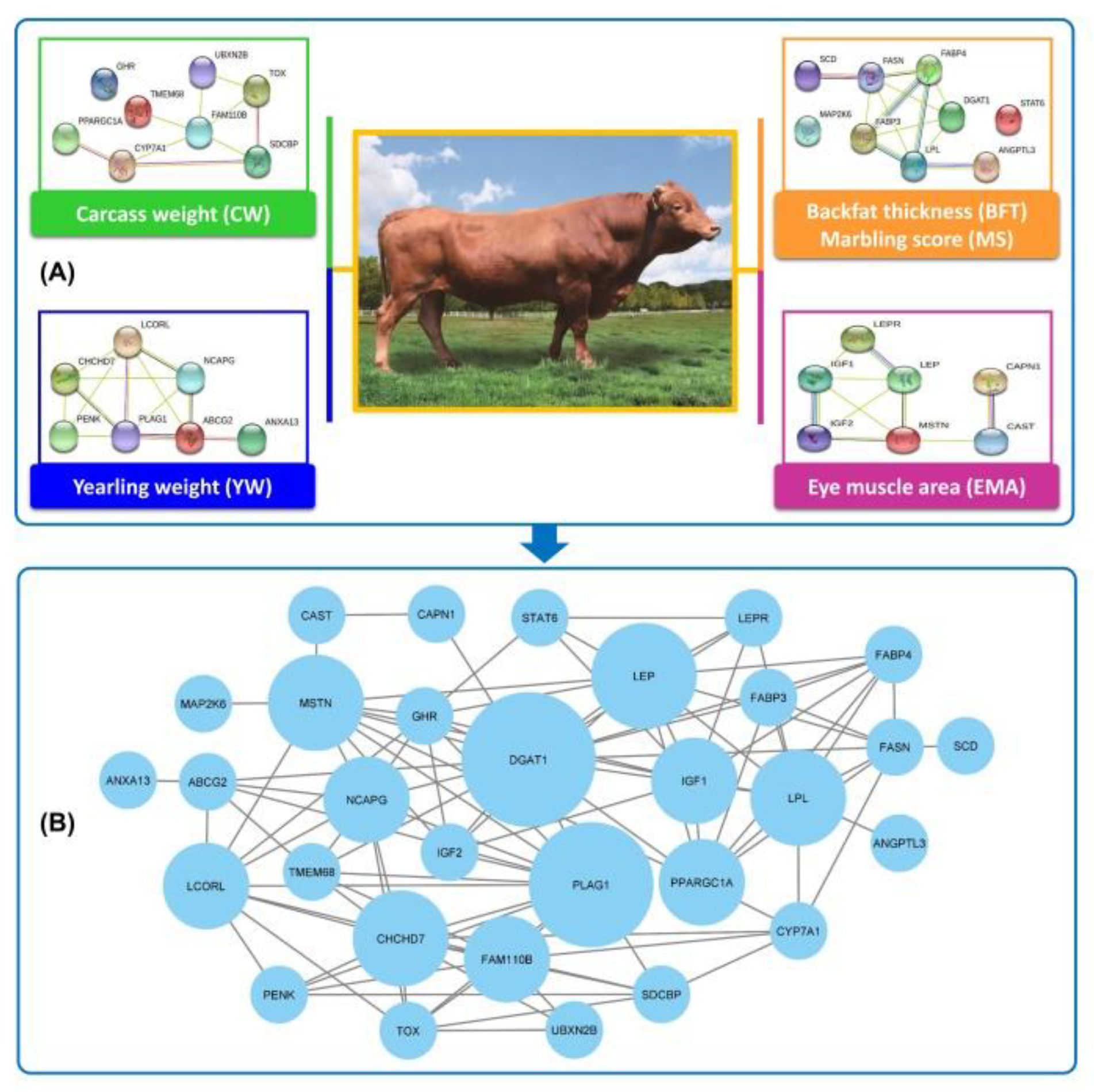
2. Candidate Genes Affecting Back Fat Thickness and Marbling Score
3. Candidate Genes Affecting Carcass Weight
4. Candidate Genes Affecting Eye Muscle Area
5. Candidate Genes Affecting Yearling Weight
6. Protein–Protein Interaction (PPI) Network and Identification of Hub Genes
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santana, M.H.d.A.; Ventura, R.V.; Utsunomiya, Y.T.; Neves, H.H.d.R.; Alexandre, P.A.; Oliveira Junior, G.A.; Gomes, R.d.C.; Bonin, M.d.N.; Coutinho, L.L.; Garcia, J.F. A genomewide association mapping study using ultrasound-scanned information identifies potential genomic regions and candidate genes affecting carcass traits in Nellore cattle. J. Anim. Breed. Genet. 2015, 132, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Jo, C.; Cho, S.; Chang, J.; Nam, K. Keys to production and processing of Hanwoo beef: A perspective of tradition and science. Anim. Front. 2012, 2, 32–38. [Google Scholar] [CrossRef]
- Lee, S.-H.; Park, B.-H.; Sharma, A.; Dang, C.-G.; Lee, S.-S.; Choi, T.-J.; Choy, Y.-H.; Kim, H.-C.; Jeon, K.-J.; Kim, S.-D. Hanwoo cattle: Origin, domestication, breeding strategies and genomic selection. J. Anim. Sci. Technol. 2014, 56, 2. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Choi, B.H.; Lim, D.; Gondro, C.; Cho, Y.M.; Dang, C.G.; Sharma, A.; Jang, G.W.; Lee, K.T.; Yoon, D. Genome-wide association study identifies major loci for carcass weight on BTA14 in Hanwoo (Korean cattle). PLoS ONE 2013, 8, e74677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Alam, M.; Park, M.N. Breeding initiatives for Hanwoo cattle to thrive as a beef industry—A review study. J. Anim. Breed. Genom. 2017, 1, 103. [Google Scholar]
- Park, B.; Choi, T.; Kim, S.; Oh, S.-H. National genetic evaluation (system) of Hanwoo (Korean native cattle). Asian-Australas. J. Anim. Sci. 2013, 26, 151. [Google Scholar] [CrossRef] [PubMed]
- Matukumalli, L.K.; Lawley, C.T.; Schnabel, R.D.; Taylor, J.F.; Allan, M.F.; Heaton, M.P.; O’Connell, J.; Moore, S.S.; Smith, T.P.; Sonstegard, T.S. Development and characterization of a high density SNP genotyping assay for cattle. PLoS ONE 2009, 4, e5350. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.-L.; Yu, Y.-W.; Chung, R.-H. Pathway analysis incorporating protein-protein interaction networks identified candidate pathways for the seven common diseases. PLoS ONE 2016, 11, e0162910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikanth, K.; Lee, S.-H.; Chung, K.-Y.; Park, J.-E.; Jang, G.-W.; Park, M.-R.; Kim, N.Y.; Kim, T.-H.; Chai, H.-H.; Park, W.C. A gene-set enrichment and protein–protein interaction network-based GWAS with regulatory SNPs identifies candidate genes and pathways associated with carcass traits in hanwoo cattle. Genes 2020, 11, 316. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, M.S.; Lim, D.; Park, M.; Lee, S.; Kim, Y.; Gondro, C.; Park, B.; Lee, S. Functional partitioning of genomic variance and genome-wide association study for carcass traits in Korean Hanwoo cattle using imputed sequence level SNP data. Front. Genet. 2018, 9, 217. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Kim, Y.-S.; Iqbal, A.; Kim, J.-J. A whole genome association study to detect additive and dominant single nucleotide polymorphisms for growth and carcass traits in Korean native cattle, Hanwoo. Asian-Australas. J. Anim. Sci. 2017, 30, 8. [Google Scholar] [CrossRef] [PubMed]
- Naserkheil, M.; Bahrami, A.; Lee, D.; Mehrban, H. Integrating single-step GWAS and bipartite networks reconstruction provides novel insights into yearling weight and carcass traits in hanwoo beef cattle. Animals 2020, 10, 1836. [Google Scholar] [CrossRef] [PubMed]
- El-Hack, A.; Mohamed, E.; Abdelnour, S.A.; Swelum, A.A.; Arif, M. The application of gene marker-assisted selection and proteomics for the best meat quality criteria and body measurements in Qinchuan cattle breed. Mol. Biol. Rep. 2018, 45, 1445–1456. [Google Scholar] [CrossRef]
- Rolf, M.M.; Taylor, J.F.; Schnabel, R.D.; McKay, S.D.; McClure, M.C.; Northcutt, S.L.; Kerley, M.S.; Weaber, R.L. Impact of reduced marker set estimation of genomic relationship matrices on genomic selection for feed efficiency in Angus cattle. BMC Genet. 2010, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Cheng, H.; Garrick, D.; Golden, B.; Dekkers, J.; Park, K.; Lee, D.; Fernando, R. Comparison of alternative approaches to single-trait genomic prediction using genotyped and non-genotyped Hanwoo beef cattle. Genet. Sel. Evol. 2017, 49, 2. [Google Scholar] [CrossRef] [PubMed]
- Lopez, B.I.; Lee, S.-H.; Shin, D.-H.; Oh, J.-D.; Chai, H.-H.; Park, W.; Park, J.-E.; Lim, D. Accuracy of genomic evaluation using imputed high-density genotypes for carcass traits in commercial Hanwoo population. Livest. Sci. 2020, 241, 104256. [Google Scholar] [CrossRef]
- Mehrban, H.; Lee, D.H.; Moradi, M.H.; IlCho, C.; Naserkheil, M.; Ibáñez-Escriche, N. Predictive performance of genomic selection methods for carcass traits in Hanwoo beef cattle: Impacts of the genetic architecture. Genet. Sel. Evol. 2017, 49, 1. [Google Scholar] [CrossRef] [Green Version]
- Mehrban, H.; Lee, D.H.; Naserkheil, M.; Moradi, M.H.; Ibáñez-Escriche, N. Comparison of conventional BLUP and single-step genomic BLUP evaluations for yearling weight and carcass traits in Hanwoo beef cattle using single trait and multi-trait models. PLoS ONE 2019, 14, e0223352. [Google Scholar] [CrossRef] [Green Version]
- Mehrban, H.; Naserkheil, M.; Lee, D.; Ibáñez-Escriche, N. Multi-Trait Single-Step GBLUP Improves Accuracy of Genomic Prediction for Carcass Traits Using Yearling Weight and Ultrasound Traits in Hanwoo. Front. Genet. 2021, 12, 692356. [Google Scholar] [CrossRef]
- Mehrban, H.; Naserkheil, M.; Lee, D.H.; Cho, C.; Choi, T.; Park, M.; Ibáñez-Escriche, N. Genomic Prediction Using Alternative Strategies of Weighted Single-Step Genomic BLUP for Yearling Weight and Carcass Traits in Hanwoo Beef Cattle. Genes 2021, 12, 266. [Google Scholar] [CrossRef]
- Park, M.N.; Alam, M.; Kim, S.; Park, B.; Lee, S.H.; Lee, S.S. Genomic selection through single-step genomic best linear unbiased prediction improves the accuracy of evaluation in Hanwoo cattle. Asian-Australas. J. Anim. Sci. 2020, 33, 1544. [Google Scholar] [CrossRef] [PubMed]
- Koohmaraie, M.; Kent, M.P.; Shackelford, S.D.; Veiseth, E.; Wheeler, T.L. Meat tenderness and muscle growth: Is there any relationship? Meat Sci. 2002, 62, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros de Oliveira Silva, R.; Bonvino Stafuzza, N.; de Oliveira Fragomeni, B.; Miguel Ferreira de Camargo, G.; Matos Ceacero, T.; Noely dos Santos Gonçalves Cyrillo, J.; Baldi, F.; Augusti Boligon, A.; Zerlotti Mercadante, M.E.; Lino Lourenco, D. Genome-wide association study for carcass traits in an experimental Nelore cattle population. PLoS ONE 2017, 12, e0169860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roudbari, Z.; Coort, S.L.; Kutmon, M.; Eijssen, L.; Melius, J.; Sadkowski, T.; Evelo, C.T. Identification of biological pathways contributing to marbling in skeletal muscle to improve beef cattle breeding. Front. Genet. 2020, 10, 1370. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Jin, S.; Kim, H.-J.; Bhuiyan, M.S.A.; Lee, D.H.; Lee, S.H.; Jang, S.B.; Han, M.H.; Lee, S.H. Validation Study of SNPs in CAPN1-CAST Genes on the Tenderness of Muscles (Longissimus thoracis and Semimembranosus) in Hanwoo (Korean Cattle). Animals 2019, 9, 691. [Google Scholar] [CrossRef] [Green Version]
- Magalhaes, A.F.; De Camargo, G.M.; Fernandes, G.A.; Gordo, D.G.; Tonussi, R.L.; Costa, R.B.; Espigolan, R.; Silva, R.M.d.O.; Bresolin, T.; De Andrade, W.B. Genome-wide association study of meat quality traits in Nellore cattle. PLoS ONE 2016, 11, e0157845. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A. Genome-wide association study for carcass traits in a composite beef cattle breed. Livest. Sci. 2018, 213, 35–43. [Google Scholar]
- Schenkel, F.; Miller, S.; Jiang, Z.; Mandell, I.; Ye, X.; Li, H.; Wilton, J. Association of a single nucleotide polymorphism in the calpastatin gene with carcass and meat quality traits of beef cattle. J. Anim. Sci. 2006, 84, 291–299. [Google Scholar] [CrossRef]
- Chmurzyńska, A. The multigene family of fatty acid-binding proteins (FABPs): Function, structure and polymorphism. J. Appl. Genet. 2006, 47, 39–48. [Google Scholar] [CrossRef]
- Veerkamp, J.H.; Maatman, R.G. Cytoplasmic fatty acid-binding proteins: Their structure and genes. Prog. Lipid Res. 1995, 34, 17–52. [Google Scholar] [CrossRef]
- Roy, R.; Calvo, J.; Hayes, H.; Rodellar, C.; Eggen, A. Fine mapping of the bovine heart fatty acid-binding protein gene (FABP3) to BTA2q45 by fluorescence in situ hybridization and radiation hybrid mapping. Anim. Genet. 2003, 34, 466–467. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.; Marcos, S.; Jurado, J.; Serrano, M. Association of the heart fatty acid-binding protein (FABP3) gene with milk traits in Manchega breed sheep. Anim. Genet. 2004, 35, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Qu, G.; Ding, S.; Dong, X. The Research Progress of Beef Cattle Intramuscular Fat Deposition Related Genes. In Proceedings of the 7th International Conference on Education, Management, Computer and Society (EMCS 2017), Shenyang, China, 17–19 March 2017; pp. 956–959. [Google Scholar]
- Shimano, H.; Horton, J.D.; Hammer, R.E.; Shimomura, I.; Brown, M.S.; Goldstein, J.L. Overproduction of cholesterol and fatty acids causes massive liver enlargement in transgenic mice expressing truncated SREBP-1a. J. Clin. Investig. 1996, 98, 1575–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.; Chai, H.-H.; Lee, S.-H.; Cho, Y.-M.; Choi, J.-W.; Kim, N.-K. Gene expression patterns associated with peroxisome proliferator-activated receptor (PPAR) signaling in the longissimus dorsi of Hanwoo (Korean cattle). Asian-Australas. J. Anim. Sci. 2015, 28, 1075. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Park, T.-S.; Yoon, D.-H.; Cheong, H.-S.; Namgoong, S.; Park, B.-L.; Lee, H.-W.; Han, C.-S.; Kim, E.-M.; Cheong, I.-C. Identification of genetic polymorphisms in FABP3 and FABP4 and putative association with back fat thickness in Korean native cattle. BMB Rep. 2008, 41, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Matarese, V.; Bernlohr, D. Purification of murine adipocyte lipid-binding protein. Characterization as a fatty acid-and retinoic acid-binding protein. J. Biol. Chem. 1988, 263, 14544–14551. [Google Scholar] [CrossRef]
- Casas, E.; Shackelford, S.; Keele, J.; Koohmaraie, M.; Smith, T.; Stone, R. Detection of quantitative trait loci for growth and carcass composition in cattle. J. Anim. Sci. 2003, 81, 2976–2983. [Google Scholar] [CrossRef] [Green Version]
- Maharani, D.; Jung, Y.; Jung, W.; Jo, C.; Ryoo, S.; Lee, S.; Yeon, S.; Lee, J. Association of five candidate genes with fatty acid composition in Korean cattle. Mol. Biol. Rep. 2012, 39, 6113–6121. [Google Scholar] [CrossRef]
- Shin, S.-C.; Heo, J.-P.; Chung, E.-R. Genetic variants of the FABP4 gene are associated with marbling scores and meat quality grades in Hanwoo (Korean cattle). Mol. Biol. Rep. 2012, 39, 5323–5330. [Google Scholar] [CrossRef]
- Lee, H.J.; Chung, Y.J.; Jang, S.; Seo, D.W.; Lee, H.K.; Yoon, D.; Lim, D.; Lee, S.H. Genome-wide identification of major genes and genomic prediction using high-density and text-mined gene-based SNP panels in Hanwoo (Korean cattle). PLoS ONE 2020, 15, e0241848. [Google Scholar] [CrossRef]
- Lee, S.H.; Van Der Werf, J.; Lee, S.; Park, E.; Oh, S.; Gibson, J.; Thompson, J.M. Genetic polymorphisms of the bovine fatty acid binding protein 4 gene are significantly associated with marbling and carcass weight in Hanwoo (Korean Cattle). Anim. Genet. 2010, 41, 442–444. [Google Scholar]
- Chakravarty, B.; Gu, Z.; Chirala, S.S.; Wakil, S.J.; Quiocho, F.A. Human fatty acid synthase: Structure and substrate selectivity of the thioesterase domain. Proc. Natl. Acad. Sci. USA 2004, 101, 15567–15572. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.; Kwon, E.; Im, S.; Seo, K.; Baik, M. Expression of fat deposition and fat removal genes is associated with intramuscular fat content in longissimus dorsi muscle of Korean cattle steers. J. Anim. Sci. 2012, 90, 2044–2053. [Google Scholar] [CrossRef]
- Mazzucco, J.P.; Goszczynski, D.E.; Ripoli, M.V.; Melucci, L.M.; Pardo, A.M.; Colatto, E.; Rogberg-Muñoz, A.; Mezzadra, C.; Depetris, G.; Giovambattista, G. Growth, carcass and meat quality traits in beef from Angus, Hereford and cross-breed grazing steers, and their association with SNPs in genes related to fat deposition metabolism. Meat Sci. 2016, 114, 121–129. [Google Scholar] [CrossRef]
- Oh, D.; Lee, Y.; La, B.; Yeo, J.; Chung, E.; Kim, Y.; Lee, C. Fatty acid composition of beef is associated with exonic nucleotide variants of the gene encoding FASN. Mol. Biol. Rep. 2012, 39, 4083–4090. [Google Scholar] [CrossRef]
- Yeon, S.; Lee, S.; Choi, B.; Lee, H.; Jang, G.; Lee, K.; Kim, K.; Lee, J.; Chung, H. Genetic variation of FASN is associated with fatty acid composition of Hanwoo. Meat Sci. 2013, 94, 133–138. [Google Scholar] [CrossRef]
- Bhuiyan, M.S.; Kim, Y.K.; Kim, H.J.; Lee, D.H.; Lee, S.H.; Yoon, H.B.; Lee, S.H. Genome-wide association study and prediction of genomic breeding values for fatty-acid composition in Korean Hanwoo cattle using a high-density single-nucleotide polymorphism array. J. Anim. Sci. 2018, 96, 4063–4075. [Google Scholar] [CrossRef] [PubMed]
- Cases, S.; Smith, S.J.; Zheng, Y.-W.; Myers, H.M.; Lear, S.R.; Sande, E.; Novak, S.; Collins, C.; Welch, C.B.; Lusis, A.J. Identification of a gene encoding an acyl CoA: Diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13018–13023. [Google Scholar] [CrossRef] [Green Version]
- Grisart, B.; Coppieters, W.; Farnir, F.; Karim, L.; Ford, C.; Berzi, P.; Cambisano, N.; Mni, M.; Reid, S.; Simon, P. Positional candidate cloning of a QTL in dairy cattle: Identification of a missense mutation in the bovine DGAT1 gene with major effect on milk yield and composition. Genome Res. 2002, 12, 222–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, A.; Krämer, W.; Werner, F.A.; Kollers, S.; Kata, S.; Durstewitz, G.; Buitkamp, J.; Womack, J.E.; Thaller, G.; Fries, R. Association of a lysine-232/alanine polymorphism in a bovine gene encoding acyl-CoA: Diacylglycerol acyltransferase (DGAT1) with variation at a quantitative trait locus for milk fat content. Proc. Natl. Acad. Sci. USA 2002, 99, 9300–9305. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.; Oh, J.; Lee, J.; Yoon, D.; Choi, Y.; Cho, B.; Lee, H.; Jeon, G. Association of sequence variations in DGAT 1 gene with economic traits in Hanwoo (Korea cattle). Asian-Australas. J. Anim. Sci. 2007, 20, 817–820. [Google Scholar] [CrossRef]
- Li, X.; Ekerljung, M.; Lundström, K.; Lundén, A. Association of polymorphisms at DGAT1, leptin, SCD1, CAPN1 and CAST genes with color, marbling and water holding capacity in meat from beef cattle populations in Sweden. Meat Sci. 2013, 94, 153–158. [Google Scholar] [CrossRef]
- Tait, R., Jr.; Shackelford, S.; Wheeler, T.; King, D.; Keele, J.; Casas, E.; Smith, T.; Bennett, G. CAPN1, CAST, and DGAT1 genetic effects on preweaning performance, carcass quality traits, and residual variance of tenderness in a beef cattle population selected for haplotype and allele equalization. J. Anim. Sci. 2014, 92, 5382–5393. [Google Scholar] [CrossRef]
- Thaller, G.; Kühn, C.; Winter, A.; Ewald, G.; Bellmann, O.; Wegner, J.; Zühlke, H.; Fries, R. DGAT1, a new positional and functional candidate gene for intramuscular fat deposition in cattle. Anim. Genet. 2003, 34, 354–357. [Google Scholar] [CrossRef]
- Wu, X.-L.; MacNeil, M.D.; De, S.; Xiao, Q.-J.; Michal, J.J.; Gaskins, C.T.; Reeves, J.J.; Busboom, J.R.; Wright, R.W.; Jiang, Z. Evaluation of candidate gene effects for beef backfat via Bayesian model selection. Genetica 2005, 125, 103–113. [Google Scholar] [CrossRef]
- Joo, S.-T.; Hwang, Y.-H.; Frank, D. Characteristics of Hanwoo cattle and health implications of consuming highly marbled Hanwoo beef. Meat Sci. 2017, 132, 45–51. [Google Scholar] [CrossRef]
- Beak, S.-H.; Lee, Y.; Lee, E.B.; Kim, K.H.; Kim, J.G.; Bok, J.D.; Kang, S.-K. Study on the fatty acid profile of phospholipid and neutral lipid in Hanwoo beef and their relationship to genetic variation. J. Anim. Sci. Technol. 2019, 61, 69. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Sharma, A.; Lee, S.H.; Lee, D.H.; Cho, Y.M.; Yang, B.S.; Lee, S.H. Genetic association of PLAG1, SCD, CYP7B1 and FASN SNPs and their effects on carcass weight, intramuscular fat and fatty acid composition in Hanwoo steers (Korean cattle). Anim. Genet. 2017, 48, 251–252. [Google Scholar] [CrossRef]
- Lee, S.-H.; Cho, Y.-M.; Lee, S.-H.; Kim, B.-S.; Kim, N.-K.; Choy, Y.-H.; Kim, K.-H.; Yoon, D.-H.; Im, S.-K.; Oh, S.-J. Identification of marbling-related candidate genes in M. longissimus dorsi of high-and low marbled Hanwoo (Korean Native Cattle) steers. BMB Rep. 2008, 41, 846–851. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.-y.; Lee, Y.-s.; Yeo, J.-s. Identification of the SNP (single necleotide polymorphism) of the stearoyl-CoA desaturase (SCD) associated with unsaturated fatty acid in Hanwoo (Korean cattle). Asian-Australas. J. Anim. Sci. 2011, 24, 757–765. [Google Scholar] [CrossRef]
- Shin, S.; Heo, J.; Chung, K.; Chung, E. Validation of molecular diagnostic markers for carcass traits in commercial Hanwoo steers. In Proceedings of the 56th International Congress of Meat Science and Technology (56th ICoMST), Jeju, Korea, 15–20 August 2010; pp. 15–20. [Google Scholar]
- Rincon, G.; Farber, E.; Farber, C.; Nkrumah, J.; Medrano, J. Polymorphisms in the STAT6 gene and their association with carcass traits in feedlot cattle. Anim. Genet. 2009, 40, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Maffei, Á.; Halaas, J.; Ravussin, E.; Pratley, R.; Lee, G.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.; Fei, H.; Lee, G.-H.; Dani, C.; Leroy, P.; Zhang, Y.; Proenca, R.; Negrel, R.; Ailhaud, G.; Friedman, J.M. Increased expression in adipocytes of ob RNA in mice with lesions of the hypothalamus and with mutations at the db locus. Proc. Natl. Acad. Sci. USA 1995, 92, 6957–6960. [Google Scholar] [CrossRef] [PubMed]
- Foka, P.; Karamichali, E.; Dalagiorgou, G.; Serti, E.; Doumba, P.P.; Pissas, G.; Kakkanas, A.; Kazazi, D.; Kochlios, E.; Gaitanou, M. Hepatitis C virus modulates lipid regulatory factor Angiopoietin-like 3 gene expression by repressing HNF-1α activity. J. Hepatol. 2014, 60, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Arca, M.; Minicocci, I.; Maranghi, M. The angiopoietin-like protein 3: A hepatokine with expanding role in metabolism. Curr. Opin. Lipidol. 2013, 24, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.-F.; Ortigues-Marty, I.; Pethick, D.; Herpin, P.; Fernandez, X. Nutritional and hormonal regulation of energy metabolism in skeletal muscles of meat-producing animals. Livest. Prod. Sci. 1998, 56, 115–143. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Byrne, K.A.; Reverter, A.; Harper, G.S.; Taniguchi, M.; McWilliam, S.M.; Mannen, H.; Oyama, K.; Lehnert, S.A. Transcriptional profiling of skeletal muscle tissue from two breeds of cattle. Mamm. Genome 2005, 16, 201–210. [Google Scholar] [CrossRef]
- Ryu, J.; Kim, Y.; Kim, C.; Kim, J.; Lee, C. Association of bovine carcass phenotypes with genes in an adaptive thermogenesis pathway. Mol. Biol. Rep. 2012, 39, 1441–1445. [Google Scholar] [CrossRef]
- Lemos, M.V.; Chiaia, H.L.J.; Berton, M.P.; Feitosa, F.L.; Aboujaoud, C.; Camargo, G.M.; Pereira, A.S.; Albuquerque, L.G.; Ferrinho, A.M.; Mueller, L.F. Genome-wide association between single nucleotide polymorphisms with beef fatty acid profile in Nellore cattle using the single step procedure. BMC Genom. 2016, 17, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Van Der Werf, J.; Kim, N.K.; Lee, S.H.; Gondro, C.; Park, E.W.; Oh, S.J.; Gibson, J.P.; Thompson, J.M. QTL and gene expression analyses identify genes affecting carcass weight and marbling on BTA14 in Hanwoo (Korean Cattle). Mamm. Genome 2011, 22, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-Y.; Lee, J.-Y.; Jang, J.-E.; Lee, S.-U. Genetic effects of sterol regulatory element binding proteins and fatty acid-binding protein4 on the fatty acid composition of Korean cattle (Hanwoo). Asian-Australas. J. Anim. Sci. 2017, 30, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priatno, W.; Jo, Y.-H.; Nejad, J.G.; Lee, J.-S.; Moon, J.-O.; Lee, H.-G. “Dietary supplementation of L-tryptophan” increases muscle development, adipose tissue catabolism and fatty acid transportation in the muscles of Hanwoo steers. J. Anim. Sci. Technol. 2020, 62, 595. [Google Scholar] [CrossRef]
- Irshad, A.; Kandeepan, G.; Kumar, S.; Ashish, K.; Vishnuraj, M.; Shukla, V. Factors influencing carcass composition of livestock: A review. J. Anim. Prod. Adv. 2013, 3, 177–186. [Google Scholar]
- Lam, C.K.; Chen, J.; Cao, Y.; Yang, L.; Wong, Y.M.; Yeung, S.Y.V.; Yao, X.; Huang, Y.; Chen, Z.-Y. Conjugated and non-conjugated octadecaenoic acids affect differently intestinal acyl coenzyme A: Cholesterol acyltransferase activity. Atherosclerosis 2008, 198, 85–93. [Google Scholar] [CrossRef]
- Monte, M.J.; Marin, J.J.; Antelo, A.; Vazquez-Tato, J. Bile acids: Chemistry, physiology, and pathophysiology. World J. Gastroenterol. WJG 2009, 15, 804. [Google Scholar] [CrossRef]
- Brunes, L.C.; Baldi, F.; Lopes, F.B.; Lôbo, R.B.; Espigolan, R.; Costa, M.F.; Stafuzza, N.B.; Magnabosco, C.U. Weighted single-step genome-wide association study and pathway analyses for feed efficiency traits in Nellore cattle. J. Anim. Breed. Genet. 2021, 138, 23–44. [Google Scholar] [CrossRef]
- Seabury, C.M.; Oldeschulte, D.L.; Saatchi, M.; Beever, J.E.; Decker, J.E.; Halley, Y.A.; Bhattarai, E.K.; Molaei, M.; Freetly, H.C.; Hansen, S.L. Genome-wide association study for feed efficiency and growth traits in US beef cattle. BMC Genom. 2017, 18, 386. [Google Scholar] [CrossRef] [Green Version]
- Terakado, A.; Costa, R.; De Camargo, G.; Irano, N.; Bresolin, T.; Takada, L.; Carvalho, C.; Oliveira, H.; Carvalheiro, R.; Baldi, F. Genome-wide association study for growth traits in Nelore cattle. Animal 2018, 12, 1358–1362. [Google Scholar] [CrossRef] [Green Version]
- Lindholm-Perry, A.; Kuehn, L.; Smith, T.; Ferrell, C.; Jenkins, T.; Freetly, H.; Snelling, W. A region on BTA14 that includes the positional candidate genes LYPLA1, XKR4 and TMEM68 is associated with feed intake and growth phenotypes in cattle 1. Anim. Genet. 2012, 43, 216–219. [Google Scholar] [CrossRef]
- Fortes, M.; Lehnert, S.; Bolormaa, S.; Reich, C.; Fordyce, G.; Corbet, N.; Whan, V.; Hawken, R.; Reverter, A. Finding genes for economically important traits: Brahman cattle puberty. Anim. Prod. Sci. 2012, 52, 143–150. [Google Scholar] [CrossRef]
- de Las Heras-Saldana, S.; Lopez, B.I.; Moghaddar, N.; Park, W.; Park, J.-E.; Chung, K.Y.; Lim, D.; Lee, S.H.; Shin, D.; van der Werf, J.H. Use of gene expression and whole-genome sequence information to improve the accuracy of genomic prediction for carcass traits in Hanwoo cattle. Genet. Sel. Evol. 2020, 52, 54. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kim, J.-J. Multiple linkage disequilibrium mapping methods to validate additive quantitative trait loci in Korean native cattle (Hanwoo). Asian-Australas. J. Anim. Sci. 2015, 28, 926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Srikanth, K.; Won, S.; Son, J.-H.; Park, J.-E.; Park, W.; Chai, H.-H.; Lim, D. Haplotype-based genome-wide association study and identification of candidate genes associated with carcass traits in Hanwoo cattle. Genes 2020, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, M.; Liu, D.; Lan, X.; Lei, C.; Chen, H. The novel coding region SNPs of PPARGC1A gene and their associations with growth traits in Chinese native cattle. Mol. Biol. Rep. 2014, 41, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Puigserver, P. Tissue-specific regulation of metabolic pathways through the transcriptional coactivator PGC1-α. Int. J. Obes. 2005, 29, S5–S9. [Google Scholar] [CrossRef] [Green Version]
- Weikard, R.; Kühn, C.; Goldammer, T.; Freyer, G.; Schwerin, M. The bovine PPARGC1A gene: Molecular characterization and association of an SNP with variation of milk fat synthesis. Physiol. Genom. 2005, 21, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Chung, E. Novel SNPs in the bovine ADIPOQ and PPARGC1A genes are associated with carcass traits in Hanwoo (Korean cattle). Mol. Biol. Rep. 2013, 40, 4651–4660. [Google Scholar] [CrossRef]
- Jiang, H.; Lucy, M.C. Variants of the 5′-untranslated region of the bovine growth hormone receptor mRNA: Isolation, expression and effects on translational efficiency. Gene 2001, 265, 45–53. [Google Scholar] [CrossRef]
- Lin, B.Z.; Sasazaki, S.; Lee, J.H.; Mannen, H. Genetic diversity of growth hormone receptor gene in cattle. Anim. Sci. J. 2009, 80, 528–531. [Google Scholar] [CrossRef]
- Di Stasio, L.; Destefanis, G.; Brugiapaglia, A.; Albera, A.; Rolando, A. Polymorphism of the GHR gene in cattle and relationships with meat production and quality. Anim. Genet. 2005, 36, 138–140. [Google Scholar] [CrossRef] [PubMed]
- Putra, W.; Agung, P.; Anwar, S.; Said, S. Polymorphism of bovine growth hormone receptor gene (g. 3338A> G) and its association with body measurements and body weight in Pasundan cows. Trop. Anim. Sci. J. 2019, 42, 90–96. [Google Scholar] [CrossRef]
- Phillips, J., III. Inherited defects in growth hormone sumthesis and action. Metab. Mol. Bases Inherit. Dis. 1995, 7, 3023–3044. [Google Scholar]
- Ardicli, S.; Dincel, D.; Samli, H.; Balci, F. Effects of polymorphisms at LEP, CAST, CAPN1, GHR, FABP4 and DGAT1 genes on fattening performance and carcass traits in Simmental bulls. Arch. Anim. Breed. 2017, 60, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Sherman, E.; Nkrumah, J.; Murdoch, B.; Li, C.; Wang, Z.; Fu, A.; Moore, S. Polymorphisms and haplotypes in the bovine neuropeptide Y, growth hormone receptor, ghrelin, insulin-like growth factor 2, and uncoupling proteins 2 and 3 genes and their associations with measures of growth, performance, feed efficiency, and carcass merit in beef cattle. J. Anim. Sci. 2008, 86, 1–16. [Google Scholar]
- Bergen, R.; Miller, S.; Wilton, J. Genetic correlations among indicator traits for carcass composition measured in yearling beef bulls and finished feedlot steers. Can. J. Anim. Sci. 2005, 85, 463–473. [Google Scholar] [CrossRef]
- Corva, P.; Soria, L.; Schor, A.; Villarreal, E.; Cenci, M.P.; Motter, M.; Mezzadra, C.; Melucci, L.; Miquel, C.; Paván, E. Association of CAPN1 and CAST gene polymorphisms with meat tenderness in Bos taurus beef cattle from Argentina. Genet. Mol. Biol. 2007, 30, 1064–1069. [Google Scholar] [CrossRef] [Green Version]
- Curi, R.A.; Chardulo, L.A.L.; Mason, M.; Arrigoni, M.; Silveira, A.C.; De Oliveira, H. Effect of single nucleotide polymorphisms of CAPN1 and CAST genes on meat traits in Nellore beef cattle (Bos indicus) and in their crosses with Bos taurus. Anim. Genet. 2009, 40, 456–462. [Google Scholar] [CrossRef]
- Liu, X.; Usman, T.; Wang, Y.; Wang, Z.; Xu, X.; Wu, M.; Zhang, Y.; Zhang, X.; Li, Q.; Liu, L. Polymorphisms in epigenetic and meat quality related genes in fourteen cattle breeds and association with beef quality and carcass traits. Asian-Australas. J. Anim. Sci. 2015, 28, 467. [Google Scholar] [CrossRef] [Green Version]
- White, S.; Casas, E.; Wheeler, T.; Shackelford, S.; Koohmaraie, M.; Riley, D.; Chase, C., Jr.; Johnson, D.; Keele, J.; Smith, T. A new single nucleotide polymorphism in CAPN1 extends the current tenderness marker test to include cattle of Bos indicus, Bos taurus, and crossbred descent. J. Anim. Sci. 2005, 83, 2001–2008. [Google Scholar] [CrossRef]
- Koohmaraie, M. Biochemical factors regulating the toughening and tenderization processes of meat. Meat Sci. 1996, 43, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Page, B.; Casas, E.; Quaas, R.; Thallman, R.; Wheeler, T.; Shackelford, S.; Koohmaraie, M.; White, S.; Bennett, G.; Keele, J. Association of markers in the bovine CAPN1 gene with meat tenderness in large crossbred populations that sample influential industry sires. J. Anim. Sci. 2004, 82, 3474–3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koohmaraie, M. The role of Ca2+-dependent proteases (calpains) in post mortem proteolysis and meat tenderness. Biochimie 1992, 74, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Kulig, H.; Kmieć, M. Association between leptin gene polymorphisms and growth traits in Limousin cattle. Russ. J. Genet. 2009, 45, 738–741. [Google Scholar] [CrossRef]
- Lusk, J. Association of single nucleotide polymorphisms in the leptin gene with body weight and backfat growth curve parameters for beef cattle. J. Anim. Sci. 2007, 85, 1865–1872. [Google Scholar] [CrossRef]
- Corva, P.M.; Fernandez Macedo, G.; Soria, L.; Papaleo Mazzucco, J.; Motter, M.; Villarreal, E.L.; Schor, A.; Mezzadra, C.A.; Melucci, L.M.; Miquel, M.C. Effect of leptin gene polymorphisms on growth, slaughter and meat quality traits of grazing Brangus steers. Genet. Mol. Res 2009, 8, 105–116. [Google Scholar] [CrossRef]
- Shin, S.; Chung, E. Association of SNP marker in the leptin gene with carcass and meat quality traits in Korean cattle. Asian-Australas. J. Anim. Sci. 2006, 20, 1–6. [Google Scholar] [CrossRef]
- Perez-Montarelo, D.; Fernandez, A.; Barragan, C.; Noguera, J.L.; Folch, J.M.; Rodríguez, M.C.; Ovilo, C.; Silio, L.; Fernandez, A.I. Transcriptional characterization of porcine leptin and leptin receptor genes. PLoS ONE 2013, 8, e66398. [Google Scholar] [CrossRef] [Green Version]
- Barb, C.; Hausman, G.; Houseknecht, K. Biology of leptin in the pig. Domest. Anim. Endocrinol. 2001, 21, 297–317. [Google Scholar] [CrossRef]
- Raza, S.H.A.; Liu, G.-Y.; Zhou, L.; Gui, L.-s.; Khan, R.; Jinmeng, Y.; Chugang, M.; Schreurs, N.M.; Ji, R.; Zan, L. Detection of polymorphisms in the bovine leptin receptor gene affects fat deposition in two Chinese beef cattle breeds. Gene 2020, 758, 144957. [Google Scholar] [CrossRef]
- Kononoff, P.; Defoor, P.; Engler, M.; Swingle, R.; Gleghorn, J.; James, S.; Marquess, F. Impacts of a leptin SNP on growth performance and carcass characters in finishing steers studied over time. J. Anim. Sci. 2017, 95, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Nkrumah, J.; Li, C.; Basarab, J.; Guercio, S.; Meng, Y.; Murdoch, B.; Hansen, C.; Moore, S. Association of a single nucleotide polymorphism in the bovine leptin gene with feed intake, feed efficiency, growth, feeding behaviour, carcass quality and body composition. Can. J. Anim. Sci. 2004, 84, 211–219. [Google Scholar] [CrossRef]
- Souza, F.; Mercadante, M.; Fonseca, L.; Ferreira, L.; Regatieri, I.; Ayres, D.; Tonhati, H.; Silva, S.; Razook, A.; Albuquerque, L.G.d. Assessment of DGAT1 and LEP gene polymorphisms in three Nelore (Bos indicus) lines selected for growth and their relationship with growth and carcass traits. J. Anim. Sci. 2010, 88, 435–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woronuk, G.; Marquess, F.; James, S.; Palmer, J.; Berryere, T.; Deobald, H.; Howie, S.; Kononoff, P. Association of leptin genotypes with beef cattle characteristics. Anim. Genet. 2012, 43, 608–610. [Google Scholar] [CrossRef]
- Kong, H.; Oh, J.; Lee, S.; Hong, Y.; Song, W.; Lee, S.; Kim, H.; Yoo, B.; Lee, H.; Jeon, G. Association of polymorphisms in the bovine Leptin gene with ultrasound measurements for improving in Korean cattle. Asian-Australas. J. Anim. Sci. 2006, 19, 1691–1695. [Google Scholar] [CrossRef]
- Li, C.; Basarab, J.; Snelling, W.; Benkel, B.; Murdoch, B.; Hansen, C.; Moore, S. Assessment of positional candidate genes myf 5 and igf 1 for growth on bovine chromosome 5 in commercial lines of Bos taurus. J. Anim. Sci. 2004, 82, 1–7. [Google Scholar] [CrossRef]
- Van Laere, A.-S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Baxter, R.C. The insulin-like growth factors and their binding proteins. Comp. Biochem. Physiol. Part B Comp. Biochem. 1988, 91, 229–235. [Google Scholar] [CrossRef]
- Etherton, T.D. Somatotropic function: The somatomedin hypothesis revisited. J. Anim. Sci. 2004, 82, E239–E244. [Google Scholar]
- Hirai, S.; Matsumoto, H.; Hino, N.; Kawachi, H.; Matsui, T.; Yano, H. Myostatin inhibits differentiation of bovine preadipocyte. Domest. Anim. Endocrinol. 2007, 32, 1–14. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.-J. Regulation of skeletal muscle mass in mice by a new TGF-p superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Fiems, L.O. Double muscling in cattle: Genes, husbandry, carcasses and meat. Animals 2012, 2, 472–506. [Google Scholar] [CrossRef]
- Grobet, L.; Royo Martin, L.J.; Poncelet, D.; Pirottin, D.; Brouwers, B.; Riquet, J.; Schoeberlein, A.; Dunner, S.; Ménissier, F.; Massabanda, J. A deletion in the bovine myostatin gene causes the double–muscled phenotype in cattle. Nat. Genet. 1997, 17, 71–74. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lee, S.-J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.S.; Yoon, D.-H.; Park, B.L.; Kim, L.H.; Bae, J.S.; Namgoong, S.; Lee, H.W.; Han, C.S.; Kim, J.O.; Cheong, I.-C. A single nucleotide polymorphism in CAPN1 associated with marbling score in Korean cattle. BMC Genet. 2008, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Dang, C.; Cho, S.; Sharma, A.; Kim, H.; Jeon, G.; Yeon, S.; Hong, S.; Park, B.; Kang, H.; Lee, S. Genome-wide association study for Warner-Bratzler shear force and sensory traits in Hanwoo (Korean cattle). Asian-Australas. J. Anim. Sci. 2014, 27, 1328. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.; Lee, S.H.; Gondro, C.; Choi, B.H.; Chai, H.H.; Strucken, E.; Cho, Y.M. Identification of selection footprints in a brown Hanwoo (Korean cattle) population for production traits. In Proceedings of the 10th World Congress on Genetics Applied to Livestock Production (WCGALP), Vancouver, BC, Canada, 17–22 August 2014. [Google Scholar]
- Yang, Y.B.; Pandurangan, M.; Jeong, D.; Hwang, I. The effect of troglitazone on lipid accumulation and related gene expression in Hanwoo muscle satellite cell. J. Physiol. Biochem. 2013, 69, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Davis, M. Effects of genetic variants for the calpastatin gene on calpastatin activity and meat tenderness in Hanwoo (Korean cattle). Meat Sci. 2012, 90, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Kim, S.-C.; Chai, H.-H.; Cho, S.-H.; Kim, H.-C.; Lim, D.; Choi, B.-H.; Dang, C.-G.; Sharma, A.; Gondro, C. Mutations in calpastatin and μ-calpain are associated with meat tenderness, flavor and juiciness in Hanwoo (Korean cattle): Molecular modeling of the effects of substitutions in the calpastatin/μ-calpain complex. Meat Sci. 2014, 96, 1501–1508. [Google Scholar] [CrossRef]
- Shin, Y.; Jung, H.-j.; Jung, M.; Yoo, S.; Subramaniyam, S.; Markkandan, K.; Kang, J.-M.; Rai, R.; Park, J.; Kim, J.-J. Discovery of gene sources for economic traits in Hanwoo by whole-genome resequencing. Asian-Australas. J. Anim. Sci. 2016, 29, 1353. [Google Scholar] [CrossRef] [Green Version]
- Seong, J.; Yoon, H.; Kong, H.S. Identification of microRNA and target gene associated with marbling score in Korean cattle (Hanwoo). Genes Genom. 2016, 38, 529–538. [Google Scholar] [CrossRef]
- Han, S.-H.; Cho, I.-C.; Ko, M.-S.; Kim, E.-Y.; Park, S.-P.; Lee, S.-S.; Oh, H.-S. A promoter polymorphism of MSTN g.− 371T> A and its associations with carcass traits in Korean cattle. Mol. Biol. Rep. 2012, 39, 3767–3772. [Google Scholar] [CrossRef] [PubMed]
- Mehrban, H.; Naserkheil, M.; Lee, D.H.; Ibáñez-Escriche, N. Genetic parameters and correlations of related feed efficiency, growth, and carcass traits in Hanwoo beef cattle. Anim. Biosci. 2021, 34, 824. [Google Scholar] [CrossRef]
- Fink, T.; Tiplady, K.; Lopdell, T.; Johnson, T.; Snell, R.G.; Spelman, R.J.; Davis, S.R.; Littlejohn, M.D. Functional confirmation of PLAG1 as the candidate causative gene underlying major pleiotropic effects on body weight and milk characteristics. Sci. Rep. 2017, 7, 44793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindholm-Perry, A.K.; Kuehn, L.A.; Oliver, W.T.; Sexten, A.K.; Miles, J.R.; Rempel, L.A.; Cushman, R.A.; Freetly, H.C. Adipose and muscle tissue gene expression of two genes (NCAPG and LCORL) located in a chromosomal region associated with cattle feed intake and gain. PLoS ONE 2013, 8, e80882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Littlejohn, M.; Grala, T.; Sanders, K.; Walker, C.; Waghorn, G.; Macdonald, K.; Coppieters, W.; Georges, M.; Spelman, R.; Hillerton, E. Genetic variation in PLAG1 associates with early life body weight and peripubertal weight and growth in Bos taurus. Anim. Genet. 2012, 43, 591–594. [Google Scholar] [CrossRef]
- Setoguchi, K.; Furuta, M.; Hirano, T.; Nagao, T.; Watanabe, T.; Sugimoto, Y.; Takasuga, A. Cross-breed comparisons identified a critical 591-kb region for bovine carcass weight QTL (CW-2) on chromosome 6 and the Ile-442-Met substitution in NCAPG as a positional candidate. BMC Genet. 2009, 10, 43. [Google Scholar] [CrossRef]
- Setoguchi, K.; Watanabe, T.; Weikard, R.; Albrecht, E.; Kühn, C.; Kinoshita, A.; Sugimoto, Y.; Takasuga, A. The SNP c. 1326T> G in the non-SMC condensin I complex, subunit G (NCAPG) gene encoding a p. Ile442Met variant is associated with an increase in body frame size at puberty in cattle. Anim. Genet. 2011, 42, 650–655. [Google Scholar] [CrossRef]
- Snelling, W.; Allan, M.; Keele, J.; Kuehn, L.; McDaneld, T.; Smith, T.; Sonstegard, T.; Thallman, R.; Bennett, G. Genome-wide association study of growth in crossbred beef cattle. J. Anim. Sci. 2010, 88, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Utsunomiya, Y.T.; Do Carmo, A.S.; Carvalheiro, R.; Neves, H.H.; Matos, M.C.; Zavarez, L.B.; Pérez O’Brien, A.M.; Sölkner, J.; McEwan, J.C.; Cole, J.B. Genome-wide association study for birth weight in Nellore cattle points to previously described orthologous genes affecting human and bovine height. BMC Genet. 2013, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Weng, Z.; Su, H.; Saatchi, M.; Lee, J.; Thomas, M.G.; Dunkelberger, J.R.; Garrick, D.J. Genome-wide association study of growth and body composition traits in Brangus beef cattle. Livest. Sci. 2016, 183, 4–11. [Google Scholar] [CrossRef]
- Naserkheil, M.; Mehrban, H.; Lee, D.; Park, M.N. Genome-wide association study for carcass primal cut yields using single-step Bayesian approach in Hanwoo cattle. Front. Genet. 2021, 12, 2343. [Google Scholar] [CrossRef] [PubMed]
- Eberlein, A.; Takasuga, A.; Setoguchi, K.; Pfuhl, R.; Flisikowski, K.; Fries, R.; Klopp, N.; Fürbass, R.; Weikard, R.; Kühn, C. Dissection of genetic factors modulating fetal growth in cattle indicates a substantial role of the non-SMC condensin I complex, subunit G (NCAPG) gene. Genetics 2009, 183, 951–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dej, K.J.; Ahn, C.; Orr-Weaver, T.L. Mutations in the Drosophila condensin subunit dCAP-G: Defining the role of condensin for chromosome condensation in mitosis and gene expression in interphase. Genetics 2004, 168, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Weikard, R.; Altmaier, E.; Suhre, K.; Weinberger, K.M.; Hammon, H.M.; Albrecht, E.; Setoguchi, K.; Takasuga, A.; Kühn, C. Metabolomic profiles indicate distinct physiological pathways affected by two loci with major divergent effect on Bos taurus growth and lipid deposition. Physiol. Genom. 2010, 42, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Yao, K.; Yin, Y.-L.; Chu, W.; Liu, Z.; Deng, D.; Li, T.; Huang, R.; Zhang, J.; Tan, B.; Wang, W. Dietary arginine supplementation increases mTOR signaling activity in skeletal muscle of neonatal pigs. J. Nutr. 2008, 138, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Hoshiba, H.; Setoguchi, K.; Watanabe, T.; Kinoshita, A.; Mizoshita, K.; Sugimoto, Y.; Takasuga, A. Comparison of the effects explained by variations in the bovine PLAG1 and NCAPG genes on daily body weight gain, linear skeletal measurements and carcass traits in Japanese Black steers from a progeny testing program. Anim. Sci. J. 2013, 84, 529–534. [Google Scholar] [CrossRef]
- McClure, M.; Morsci, N.; Schnabel, R.; Kim, J.; Yao, P.; Rolf, M.; McKay, S.; Gregg, S.; Chapple, R.; Northcutt, S. A genome scan for quantitative trait loci influencing carcass, post-natal growth and reproductive traits in commercial Angus cattle. Anim. Genet. 2010, 41, 597–607. [Google Scholar] [CrossRef]
- Pryce, J.E.; Hayes, B.J.; Bolormaa, S.; Goddard, M.E. Polymorphic regions affecting human height also control stature in cattle. Genetics 2011, 187, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, I.A.; Khatkar, M.S.; Thomson, P.C.; Raadsma, H.W. Composite selection signals for complex traits exemplified through bovine stature using multibreed cohorts of European and African Bos taurus. G3 Genes Genomes Genet. 2015, 5, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Bennett, E.J.; Huttlin, E.L.; Guo, A.; Li, J.; Possemato, A.; Sowa, M.E.; Rad, R.; Rush, J.; Comb, M.J. Systematic and quantitative assessment of the ubiquitin-modified proteome. Mol. Cell 2011, 44, 325–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindholm-Perry, A.K.; Sexten, A.K.; Kuehn, L.A.; Smith, T.P.; King, D.A.; Shackelford, S.D.; Wheeler, T.L.; Ferrell, C.L.; Jenkins, T.G.; Snelling, W.M. Association, effects and validation of polymorphisms within the NCAPG-LCORL locus located on BTA6 with feed intake, gain, meat and carcass traits in beef cattle. BMC Genet. 2011, 12, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, L.; Takeda, H.; Lin, L.; Druet, T.; Arias, J.A.; Baurain, D.; Cambisano, N.; Davis, S.R.; Farnir, F.; Grisart, B. Variants modulating the expression of a chromosome domain encompassing PLAG1 influence bovine stature. Nat. Genet. 2011, 43, 405–413. [Google Scholar] [CrossRef]
- Li, Z.; Wu, M.; Zhao, H.; Fan, L.; Zhang, Y.; Yuan, T.; He, S.; Wang, P.; Zhang, Y.; Sun, X. The PLAG1 mRNA expression analysis among genetic variants and relevance to growth traits in Chinese cattle. Anim. Biotechnol. 2020, 31, 504–511. [Google Scholar] [CrossRef]
- Nishimura, S.; Watanabe, T.; Mizoshita, K.; Tatsuda, K.; Fujita, T.; Watanabe, N.; Sugimoto, Y.; Takasuga, A. Genome-wide association study identified three major QTL for carcass weight including the PLAG1-CHCHD7 QTN for stature in Japanese Black cattle. BMC Genet. 2012, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Zatkova, A.; Rouillard, J.M.; Hartmann, W.; Lamb, B.J.; Kuick, R.; Eckart, M.; Von Schweinitz, D.; Koch, A.; Fonatsch, C.; Pietsch, T. Amplification and overexpression of the IGF2 regulator PLAG1 in hepatoblastoma. Genes Chromosom. Cancer 2004, 39, 126–137. [Google Scholar] [CrossRef]
- Song, Y.; Xu, L.; Chen, Y.; Zhang, L.; Gao, H.; Zhu, B.; Niu, H.; Zhang, W.; Xia, J.; Gao, X. Genome-wide association study reveals the PLAG1 gene for knuckle, biceps and shank weight in Simmental beef cattle. PLoS ONE 2016, 11, e0168316. [Google Scholar] [CrossRef] [Green Version]
- Turnay, J.; Lecona, E.; Fernández-Lizarbe, S.; Guzmán-Aránguez, A.; Fernández, M.P.; Olmo, N.; Lizarbe, M.A. Structure–function relationship in annexin A13, the founder member of the vertebrate family of annexins. Biochem. J. 2005, 389, 899–911. [Google Scholar] [CrossRef] [Green Version]
- Rosie, R.; Thomson, E.; Blum, M.; Roberts, J.; Fink, G. Oestrogen positive feedback reduces arcuate proopiomelanocortin messenger ribonucleic acid. J. Neuroendocrinol. 1992, 4, 625–630. [Google Scholar] [CrossRef]
- Taylor, J.A.; Goubillon, M.-L.; Broad, K.D.; Robinson, J.E. Steroid control of gonadotropin-releasing hormone secretion: Associated changes in pro-opiomelanocortin and preproenkephalin messenger RNA expression in the ovine hypothalamus. Biol. Reprod. 2007, 76, 524–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widmann, P.; Reverter, A.; Fortes, M.R.; Weikard, R.; Suhre, K.; Hammon, H.; Albrecht, E.; Kuehn, C. A systems biology approach using metabolomic data reveals genes and pathways interacting to modulate divergent growth in cattle. BMC Genom. 2013, 14, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollander, M.; Do, T.; Will, T.; Helms, V. Detecting rewiring events in protein-protein interaction networks based on transcriptomic data. Front. Bioinform. 2021, 2021, 724297. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; You, Z.-H.; Li, L.-P.; Huang, W.-Z.; Guo, J.-X.; Yu, C.-Q.; Wang, L.-P.; Zhao, Z.-Y. DWPPI: A Deep Learning Approach for Predicting Protein–Protein Interactions in Plants Based on Multi-Source Information With a Large-Scale Biological Network. Front. Bioeng. Biotechnol. 2022, 10, 807522. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Chr | Position (bp) | Reference |
|---|---|---|---|---|
| FABP3 | fatty acid-binding protein 3 | 2 | 122287099–122294639 | [35,41] |
| FABP4 | fatty acid-binding protein 4 | 14 | 44676559–44680947 | [39,40,41,42,58,60,62,72,73] |
| FASN | fatty acid synthase | 19 | 50776167–50794939 | [41,46,47,48,58] |
| DGAT1 | diacylglycerol O-acyltransferase 1 | 14 | 603981–614153 | [41,52] |
| SCD | stearoyl-CoA desaturase | 26 | 21263976–21279185 | [41,58,59,60,61,62] |
| STAT6 | signal transducer and activator of transcription 6 | 5 | 56325240–56340447 | [41] |
| ANGPTL3 | Angiopoietin-like 3 | 3 | 82994334–83003679 | [41] |
| LPL | lipoprotein lipase | 8 | 66989665–67016694 | [41,74] |
| MAP2K6 | mitogen-activated protein kinase kinase 6 | 19 | 61130409–61236793 | [11,41,70] |
| Gene Symbol | Gene Name | Chr | Position (bp) | Reference |
|---|---|---|---|---|
| SDCBP | syndecan-binding protein | 14 | 24710921–24764191 | [4,10] |
| CYP7A1 | cytochrome P450, family 7, subfamily A, polypeptide 1 | 14 | 24664833–24675169 | [4,9,10] |
| TMEM68 | transmembrane protein 68 | 14 | 23034273–23070127 | [12,85] |
| TOX | thymocyte selection-associated high-mobility group box | 14 | 24946881–25258596 | [4,10,11,83,84,85] |
| FAM110B | family with sequence similarity 110 member B | 14 | 24291600–24433683 | [4,11,12,84,85] |
| UBXN2B | UBX domain protein 2B | 14 | 24587679–24624435 | [9,11,83,84] |
| PPARGC1A | PPARG coactivator 1 alpha | 6 | 43380463–43903480 | [10,12,41,90] |
| GHR | growth hormone receptor | 20 | 31869704–32043372 | [41] |
| Gene Symbol | Gene Name | Chr | Position (bp) | Reference |
|---|---|---|---|---|
| CAPN1 | calpain 1 | 29 | 43399871–43427419 | [25,41,127,128,129,130] |
| CAST | calpastatin | 7 | 96034228–96183530 | [25,41,128,131,132,133] |
| LEP | leptin | 4 | 92436837–92453660 | [41,117] |
| LEPR | leptin receptor | 3 | 79741204–79838014 | [41] |
| IGF1 | insulin-like growth factor 1 | 5 | 66185922–66264134 | [41,133,134] |
| IGF2 | insulin-like growth factor 2 | 29 | 49395153–49422469 | [41] |
| MSTN | myostatin | 2 | 6278864–6285491 | [41,135] |
| Gene Symbol | Gene Name | Chr | Position (bp) | Reference |
|---|---|---|---|---|
| ABCG2 | ATP-binding cassette subfamily G member 2 | 6 | 36475491–36603215 | [12,145] |
| NCAPG | non-SMC condensin I complex subunit G | 6 | 37301610–37378129 | [10,41] |
| LCORL | Ligand-dependent nuclear receptor corepressor-like | 6 | 37377806–37557145 | [10,41] |
| CHCHD7 | coiled-coil-helix-coiled-coil-helix domain-containing 7 | 14 | 23376149–23383137 | [9,12] |
| PLAG1 | PLAG1 zinc finger | 14 | 23323709–23375661 | [10,12,59] |
| ANXA13 | annexin A13 | 14 | 16143324–16159206 | [9,12] |
| PENK | proenkephalin | 14 | 23542355–23546761 | [4,12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naserkheil, M.; Manzari, Z.; Dang, C.G.; Lee, S.S.; Park, M.N. Exploring and Identifying Candidate Genes and Genomic Regions Related to Economically Important Traits in Hanwoo Cattle. Curr. Issues Mol. Biol. 2022, 44, 6075-6092. https://doi.org/10.3390/cimb44120414
Naserkheil M, Manzari Z, Dang CG, Lee SS, Park MN. Exploring and Identifying Candidate Genes and Genomic Regions Related to Economically Important Traits in Hanwoo Cattle. Current Issues in Molecular Biology. 2022; 44(12):6075-6092. https://doi.org/10.3390/cimb44120414
Chicago/Turabian StyleNaserkheil, Masoumeh, Zeinab Manzari, Chang Gwon Dang, Seung Soo Lee, and Mi Na Park. 2022. "Exploring and Identifying Candidate Genes and Genomic Regions Related to Economically Important Traits in Hanwoo Cattle" Current Issues in Molecular Biology 44, no. 12: 6075-6092. https://doi.org/10.3390/cimb44120414
APA StyleNaserkheil, M., Manzari, Z., Dang, C. G., Lee, S. S., & Park, M. N. (2022). Exploring and Identifying Candidate Genes and Genomic Regions Related to Economically Important Traits in Hanwoo Cattle. Current Issues in Molecular Biology, 44(12), 6075-6092. https://doi.org/10.3390/cimb44120414