

Non-Peptide Agonists and Antagonists of the Prokineticin Receptors


Abstract
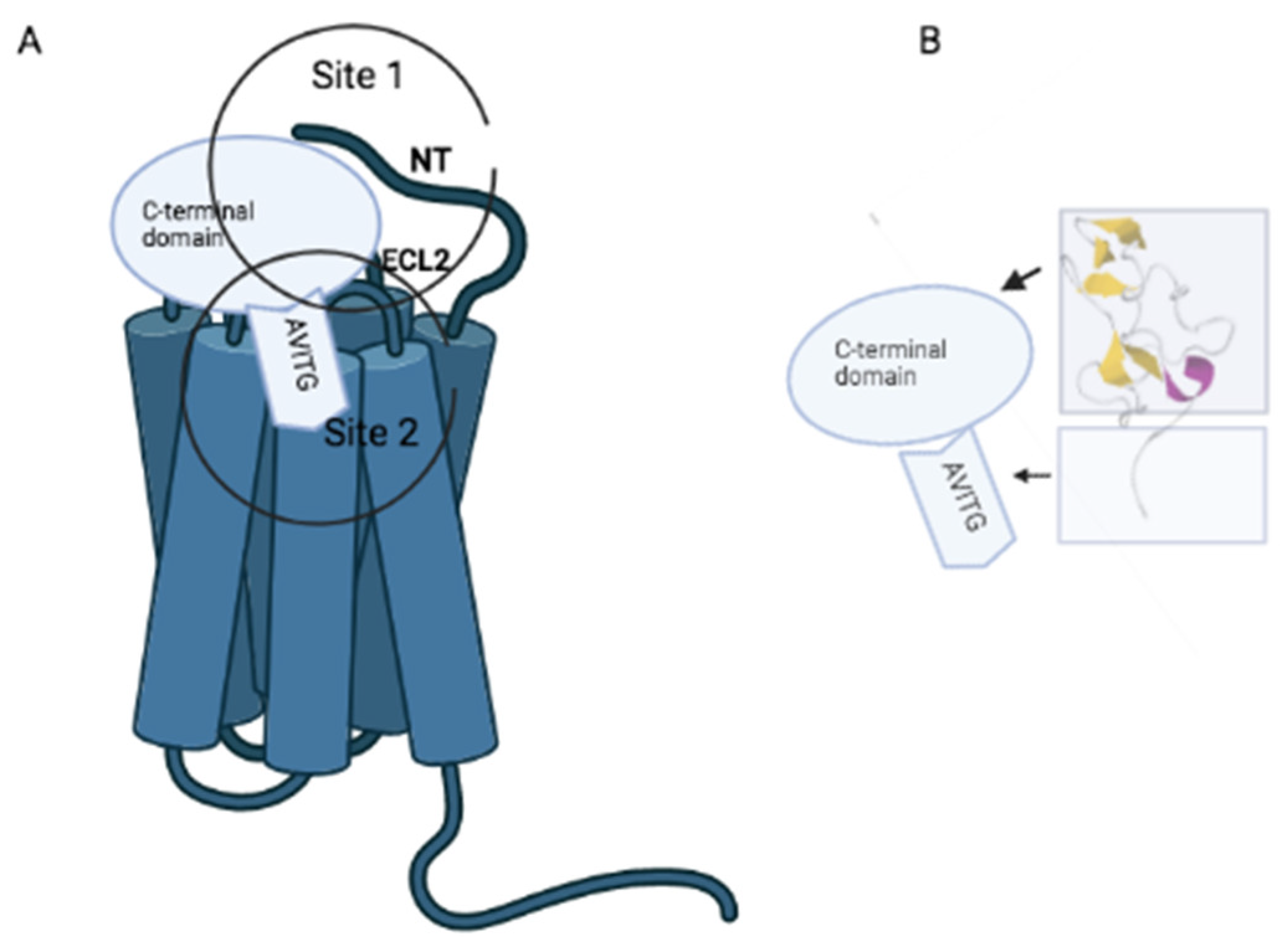
:1. Introduction
2. Receptor-Prokineticin Interactions
3. Non-Peptide Orthosteric Ligands of Prokineticin Receptors

{kind=link}
{kind=link}
| Drug | Biological System | Effect | References |
|---|---|---|---|
| PC1 | CHO cells | Inhibition of Bv8-induced intracellular Ca2+ mobilization | [41] |
| Primary cortical cultures | Inhibition of Aβ1-42 induced neuronal death; Reduction of PK2-induced increase of AMPA currents | [52,53] | |
| Tg2575 hippocampal slides | Rescue of LTP impairment | [52] | |
| Inflamed mice | Reduction of CFA-induced inflammatory pain | [44] | |
| Mice with neuropathy induced by nerve ligation | Reduction of CCI and SNI-induced neuropathic pain | [45,46,47] | |
| Mice with neuropathy induced by chemotherapeutics | Reduction of bortezomid and vincristine -induced neuropathic pain | [49,50,51] | |
| Mice with diabetic neuropathy | Reduction of streptozotocin -induced neuropathic pain | [48] | |
| Rats with intracerebroventricular infusion of Aβ1-42 | Reduction of Aβ1-42 -induced cognitive impairment | [54] | |
| PC1 and PC7 | Mice with autoimmune encephalomyelitis (EAE) | Efficacy in reducing EAE severity | [55] |
| PC7 | Primary murine cell cultures or rat organotypic hippocampal slices | Increasing of neuronal damage induced by oxygen-glucose deprivation (OGD) | [58] |
| PC7 | Porcine primary trophoblast cells | Abolishing of PK1-induced expressions of genes involved in angiogenesis | [57] |
| PKRA7 | Glioblastoma and pancreatic cancer | Inhibition of angiogenesis and macrophage infiltration | [59] |
| Arthritis | Inhibition of macrophage infiltration and production of inflammatory cytokines | [60] | |
| Cystitis | Modulation of inflammation-mediated voiding dysfunction and spontaneous visceral pain | [62] | |
| Orchitis | Inhibition inflammatory process | [61] | |
| Diabetic Cardimyocites | Inhibition of metformin effect | [63] | |
| Inhibition of trehalose effect | [64] | ||
| Pregnancy pathologies | Induction of pregnancy outcome enhancing feto-placental growth | [56] | |
| Parkinson’s Disease | Increase of the effects induced by MTPT | [65] |
4. Non-Peptides Allosteric Ligands of Prokineticin Receptors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mollay, C.; Wechselberger, C.; Mignogna, G.; Negri, L.; Melchiorri, P.; Barra, D.; Kreil, G. Bv8, a small protein from frog skin and its homologue from snake venom induce hyperalgesia in rats. Eur. J. Pharmacol. 1999, 374, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Schweitz, H.; Pacaud, P.; Diochot, S.; Moinier, D.; Lazdunski, M. MIT (1), a black mamba toxin with a new and highly potent activity on intestinal contraction. FEBS Lett. 1999, 461, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joubert, F.J.; Strydom, D.J. Snake venom. The amino acid sequence of protein A from Dendroaspis polylepis polylepis (black mamba) venom. Hoppe Seylers Z. Physiol. Chem. 1980, 361, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- LeCouter, J.; Kowalski, J.; Foster, J.; Hass, P.; Zhang, Z.; Dillard-Telm, L.; Frantz, G.; Rangell, L.; DeGuzman, L.; Keller, G.A.; et al. Identification of an angiogenic mitogen selective for endocrine gland endothelium. Nature 2001, 412, 877–884. [Google Scholar] [CrossRef] [PubMed]
- LeCouter, J.; Lin, R.; Tejada, M.; Frantz, G.; Peale, F.; Hillan, K.J.; Ferrara, N. The endocrine-gland-derived VEGF homologue Bv8 promotes angiogenesis in the testis: Localization of Bv8 receptors to endothelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 2685–2690. [Google Scholar] [CrossRef] [Green Version]
- Negri, L.; Ferrara, N. The Prokineticins: Neuromodulators and Mediators of Inflammation and Myeloid Cell-Dependent Angiogenesis. Physiol. Rev. 2018, 98, 1055–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattanzi, R.; Miele, R. Prokineticin-Receptor Network: Mechanisms of Regulation. Life 2022, 12, 172. [Google Scholar] [CrossRef]
- Kaser, A.; Winklmayr, M.; Lepperdinger, G.; Kreil, G. The AVIT protein family. Secreted cysteine rich vertebrate proteins with diverse functions. EMBO Rep. 2003, 4, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Marsango, S.; Bonaccorsi di Patti, M.C.; Barra, D.; Miele, R. Evidence that prokineticin receptor 2 exists as a dimer in vivo. Cell. Mol. Life Sci. 2011, 68, 2919–2929. [Google Scholar] [CrossRef]
- Sposini, S.; Caltabiano, G.; Hanyaloglu, A.C.; Miele, R. Identification of transmembrane domains that regulate spatial arrangements and activity of prokineticin receptor 2 dimers. Mol. Cell. Endocrinol. 2015, 399, 362–372. [Google Scholar] [CrossRef]
- Monnier, J.; Samson, M. Cytokine properties of prokineticins. FEBS J. 2008, 275, 4014–4021. [Google Scholar] [CrossRef]
- Lattanzi, R.; Severini, C.; Miele, R. Prokineticin 2 in cancer-related inflammation Cancer Lett. 2022, 546, 215838. Cancer Lett. 2022, 546, 215838. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.L.; Li, J.D.; Cheng, M.Y.; Leslie, F.M.; Lee, A.G.; Zhou, Q.Y. Dependence of olfactory bulb neurogenesis on prokineticin 2 signaling. Science 2005, 308, 1923–1927. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Yamazaki, C.; Masumoto, K.H.; Nagano, M.; Naito, M.; Soga, T.; Hiyama, H.; Matsumoto, M.; Takasaki, J.; Kamohara, M.; et al. Abnormal development of the olfactory bulb and reproductive system in mice lacking prokineticin receptor PKR2. Proc. Natl. Acad. Sci. USA 2006, 103, 4140–4145. [Google Scholar] [CrossRef] [Green Version]
- Maftei, D.; Lattanzi, R.; Vincenzi, M.; Squillace, S.; Fullone, M.R.; Miele, R. The balance of concentration between Prokineticin 2β and Prokineticin 2 modulates the food intake by STAT3 signaling. BBA Adv. 2021, 1, 100028. [Google Scholar] [CrossRef]
- Cheng, M.Y.; Bullock, C.M.; Li, C.; Lee, A.G.; Bermak, J.C.; Belluzzi, J.; Weaver, D.R.; Leslie, F.M.; Zhou, Q.Y. Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus. Nature 2002, 417, 405–410. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Cheng, M.Y. Prokineticin 2 and circadian clock output. FEBS J. 2005, 272, 5703–5709. [Google Scholar] [CrossRef]
- Negri, L.; Lattanzi, R.; Giannini, E.; De Felice, M.; Colucci, A.; Melchiorri, P. Bv8, the am- phibian homologue of the mammalian prokineticins, modulates ingestive behaviour in rats. Br. J. Pharmacol. 2004, 142, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Fullone, M.R.; Maftei, D.; Vincenzi, M.; Lattanzi, R.; Miele, R. Identification of Regions Involved in the Physical Interaction between Melanocortin Receptor Accessory Protein 2 and Prokineticin Receptor 2. Biomolecules 2022, 12, 474. [Google Scholar] [CrossRef] [PubMed]
- Fullone, M.R.; Maftei, D.; Vincenzi, M.; Lattanzi, R.; Miele, R. Arginine 125 Is an Essential Residue for the Function of MRAP2. Int. J. Mol. Sci. 2022, 23, 9853. [Google Scholar] [CrossRef] [PubMed]
- Amodeo, G.; Maftei, D.; Lattanzi, R.; Verduci, B.; Comi, L.; Galimberti, G.; Sacerdote, P.; Franchi, S. Controlling the activation of the prokineticin system as therapeutic approach to relief neuropathic pain and reduce neuroinflammation. Pharmadvances 2022, 4, 104–120. [Google Scholar] [CrossRef]
- Lattanzi, R.; Miele, R. Versatile Role of Prokineticins and Prokineticin Receptors in Neuroinflammation. Biomedicines 2021, 9, 1648. [Google Scholar] [CrossRef] [PubMed]
- Maftei, D.; Schirinzi, T.; Mercuri, N.B.; Lattanzi, R.; Severini, C. Potential Clinical Role of Prokineticin 2 (PK2) in Neurodegenerative Diseases. Curr. Neuropharmacol. 2022, 20, 2019–2023. [Google Scholar] [CrossRef] [PubMed]
- Clark-Lewis, I.; Kim, K.S.; Rajarathnam, K.; Gong, J.H.; Dewald, B.; Moser, B.; Baggiolini, M.; Sykes, B.D. Structure-activity relationships of chemokines. J. Leukoc. Biol. 1995, 57, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Boisbouvier, J.; Albrand, J.P.; Blackledge, M.; Jaquinod, M.; Schweitz, H.; Lazdunski, M.; Marion, D. A structural homologue of colipase in black mamba venom revealed by NMR floating disulphide bridge analysis. J. Mol. Biol. 1998, 283, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Morales, R.A.; Daly, N.L.; Vetter, I.; Mobli, M.; Napier, I.A.; Craik, D.J.; Lewis, R.J.; Christie, M.J.; King, G.F.; Alewood, P.F.; et al. Chemical synthesis and structure of the prokineticin Bv8. Chem. Biol. Chem. 2010, 11, 1882–1888. [Google Scholar] [CrossRef]
- Lattanzi, R.; Maftei, D.; Vincenzi, M.; Fullone, M.R.; Miele, R. Identification and Characterization of a New Splicing Variant of Prokineticin 2. Life 2022, 12, 248. [Google Scholar] [CrossRef]
- Lattanzi, R.; Maftei, D.; Negri, L.; Fusco, I.; Miele, R. PK2β ligand, a splice variant of prokineticin 2, is able to modulate and drive signaling through PKR1 receptor. Neuropeptides 2018, 71, 32–42. [Google Scholar] [CrossRef]
- Bullock, C.M.; Li, J.D.; Zhou, Q.Y. Structural determinants required for the bioactivities of prokineticins and identification of prokineticin receptor antagonists. Mol. Pharmacol. 2004, 65, 582–588. [Google Scholar] [CrossRef] [Green Version]
- Negri, L.; Lattanzi, R.; Giannini, E.; Colucci, M.; Grohovaz, F.; Codazzi, F.; Mignogna, G.; Barra, D.; Kaiser, A.; Kreil, G.; et al. Biological activity of Bv8 analogues. Br. J. Pharmacol. 2005, 146, 625–632. [Google Scholar] [CrossRef]
- Cole, L.W.; Sidis, Y.; Zhang, C.K.; Quinton, R.; Plummer, L.; Pignatelli, D.; Hughes, V.A.; Dwyer, A.A.; Raivio, T.; Hayes, F.J.; et al. Mutations in Prokineticin 2 and Prokineticin receptor 2 genes in Human Gonadotrophin-Releasing Hormone Deficiency: Molecular Genetics and Clinical Spectrum. J. Clin. Endocrinol. Metab. 2008, 93, 3551–3559. [Google Scholar] [CrossRef] [Green Version]
- Dodé, C.; Teixeira, L.; Levilliers, J.; Fouveaut, C.; Bouchard, P.; Kottler, M.-L.; Lespinasse, J.; Lienhardt-Roussie, A.; Mathieu, M.; Moerman, A.; et al. Kallmann syndrome: Mutations in the genes encoding prokineticin-2 and prokineticin receptor-2. PLoS Genet. 2006, 2, e175. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, R.; Sacerdote, P.; Franchi, S.; Canestrelli, M.; Miele, R.; Barra, D.; Visentin, S.; DeNuccio, C.; Porreca, F.; De Felice, M.; et al. Pharmacological activity of a Bv8 analogue modified in position 24. Br. J. Pharmacol. 2012, 166, 950–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miele, R.; Lattanzi, R.; Bonaccorsi di Patti, M.C.; Paiardini, A.; Negri, L.; Barra, D. Expression of Bv8 in Pichia pastoris to identify structural features for receptor binding. Protein Expr. Purif. 2010, 73, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Kufareva, I.; Salanga, C.L.; Handel, T.M. Chemokine and chemokine receptor structure and interactions: Implications for therapeutic strategies. Immunol. Cell Biol. 2015, 93, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levit, A.; Yarnitzky, T.; Wiener, A.; Meidan, R.; Niv, M.Y. Modeling of human prokineticin receptors: Interactions with novel small-molecule binders and potential off-target drugs. PLoS ONE 2011, 6, e27990. [Google Scholar] [CrossRef] [Green Version]
- Monnier, C.; Dode, C.; Fabre, L.; Teixeira, L.; Labesse, G.; Pin, J.P.; Hardelin, J.P.; Rondard, P. PROKR2 missense mutations associated with Kallmann syndrome impair receptor signalling activity. Hum. Mol. Genet. 2009, 18, 75–81. [Google Scholar] [CrossRef]
- Fullone, M.R.; Lattanzi, R.; Maftei, D.; Bonaccorsi, M.C.; Miele, R. Analysis of role of aromatic residues in extracellular loop 2 of Prokineticin receptor 2 in ligand binding probed with genetically encoded photo-crosslinkers. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183549. [Google Scholar] [CrossRef]
- Lattanzi, R.; Maftei, D.; Fullone, M.R.; Miele, R. Identification and characterization of Prokineticin receptor 2 splicing variant and its modulation in an animal model of Alzheimer’s disease. Neuropeptides 2019, 73, 49–56. [Google Scholar] [CrossRef]
- Nebigil, C.G.; Désaubry, L. The role of GPCR signaling in cardiac epithelial to mesenchymal transformation (EMT). Trends Cardiovasc. Med. 2019, 2, 200–204. [Google Scholar] [CrossRef]
- Balboni, G.; Lazzari, I.; Trapella, C.; Negri, L.; Lattanzi, R.; Giannini, E.; Nicotra, A.; Melchiorri, P.; Visentin, S.; De Nuccio, C.; et al. Triazine Compounds as Antagonists at Bv8-Prokineticin Receptors. J. Med. Chem. 2008, 51, 7635–7639. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, R.; Congiu, C.; Onnis, V.; Deplano, A.; Salvadori, S.; Marconi, V.; Maftei, D.; Francioso, A.; Ambrosio, C.; Casella, I.; et al. Halogenated triazinediones behave as antagonists of PKR1: In vitro and in vivo pharmacological characterization. Int. J. Pharm. Sci. Res. 2015, 10, 1033–1042. [Google Scholar]
- Congiu, C.; Onnis, V.; Deplano, A.; Salvadori, S.; Marconi, V.; Maftei, D.; Negri, L.; Lattanzi, R.; Balboni, G. A new convenient synthetic method and preliminary pharmacological characterization of triazinediones as prokineticin receptor antagonists. Eur. J. Med. Chem. 2014, 81, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Giannini, E.; Lattanzi, R.; Nicotra, A.; Campese, A.F.; Grazioli, P.; Screpanti, I.; Balboni, G.; Severo, S.; Sacerdote, P.; Negri, L. The chemokine Bv8/prokineticin 2 is up-regulated in inflammatory granulocytes and modulates inflammatory pain. Proc. Natl. Acad. Sci. USA 2009, 106, 14646–14651. [Google Scholar] [CrossRef] [Green Version]
- Maftei, D.; Marconi, V.; Florenzano, F.; Giancotti, L.A.; Castelli, M.; Moretti, S.; Borsani, E.; Rodella, L.F.; Balboni, G.; Luongo, L.; et al. Controlling the activation of the Bv8/prokineticin system reduces neuroinflammation and abolishes thermal and tactile hyperalgesia in neuropathic animals. Br. J. Pharmacol. 2014, 171, 4850–4865. [Google Scholar] [CrossRef] [Green Version]
- Lattanzi, R.; Maftei, D.; Marconi, V.; Florenzano, F.; Franchi, S.; Borsani, E.; Rodella, F.L.; Balboni, G.; Salvadori, S.; Sacerdote, P.; et al. Prokineticin 2 upregulation in the peripheral nervous system has a major role in triggering and maintaining neuropathic pain in the chronic constriction injury model. BioMed Res. Int. 2015, 2015, 301292. [Google Scholar] [CrossRef] [Green Version]
- Guida, F.; Lattanzi, R.; Boccella, S.; Maftei, D.; Romano, R.; Marconi, V.; Balboni, G.; Salvadori, S.; Scafuro, M.A.; De Novellis, V.; et al. PC1, a non-peptide PKR1-preferring antagonist, reduces pain behavior and spinal neuronal sensitization in neuropathic mice. Pharmacol. Res. 2015, 91, 36–46. [Google Scholar] [CrossRef]
- Castelli, M.; Amodeo, G.; Negri, L.; Lattanzi, R.; Maftei, D.; Gotti, C.; Pistillo, F.; Onnis, V.; Congiu, C.; Panerai, A.E.; et al. Antagonism of the Prokineticin System Prevents and Reverses Allodynia and Inflammation in a Mouse Model of Diabetes. PLoS ONE 2016, 11, e0146259. [Google Scholar] [CrossRef] [Green Version]
- Moschetti, G.; Kalpachidou, T.; Amodeo, G.; Lattanzi, R.; Sacerdote, P.; Kress, M.; Franchi, S. Prokineticin Receptor Inhibition With PC1 Protects Mouse Primary Sensory Neurons From Neurotoxic Effects of Chemotherapeutic Drugs in vitro. Front. Immunol. 2020, 11, 2119. [Google Scholar] [CrossRef]
- Moschetti, G.; Amodeo, G.; Paladini, M.S.; Raffaella Molteni, R.; Balboni, G.; Panerai, A.; Sacerdote, P.; Franchi, S. Prokineticin 2 promotes and sustains neuroinflammation in vincristine treated mice: Focus on pain and emotional like behavior. Brain Behav. Immun. 2019, 82, 422–431. [Google Scholar] [CrossRef]
- Moschetti, G.; Amodeo, G.; Maftei, D.; Lattanzi, R.; Procacci, P.; Sartori, P.; Balboni, G.; Onnis, V.; Conte, V.; Panerai, A.; et al. Targeting prokineticin system counteracts hypersensitivity, neuroinflammation, and tissue damage in a mouse model of bortezomib-induced peripheral neuropathy. Neuroinflammation 2019, 16, 89. [Google Scholar] [CrossRef] [PubMed]
- Severini, C.; Lattanzi, R.; Maftei, D.; Marconi, V.; Ciotti, M.T.; Petrocchi Passeri, P.; Florenzano, F.; Del Duca, E.; Caioli, S.; Zona, C.; et al. Bv8/prokineticin 2 is involved in Aβ-induced neurotoxicity. Sci. Rep. 2015, 19, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caioli, S.; Severini, C.; Ciotti, T.; Florenzano, F.; Pimpinella, D.; Petrocchi Passeri, P.; Balboni, G.; Polisca, P.; Lattanzi, R.; Nisticò, R.; et al. Prokineticin system modulation as a new target to counteract the amyloid beta toxicity induced by glutamatergic alterations in an in vitro model of Alzheimer’s disease. Neuropharmacology 2017, 116, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Maftei, D.; Ratano, P.; Fusco, I.; Marconi, V.; Squillace, S.; Negri, L.; Severini, C.; Balboni, G.; Steardo, L.; Bronzuoli, M.R.; et al. The prokineticin receptor antagonist PC1 rescues memory impairment induced by Amyloid administration through the modulation of prokineticin system. Neuropharmacology 2019, 158, 107739. [Google Scholar] [CrossRef]
- Abou-hamdan, M.; Costanza, M.; Fontana, E.; Di Dario, M.; Musio, S.; Congiu, C.; Onnis, V.; Lattanzi, R.; Radaelli, M.; Martinelli, V.; et al. Critical role for prokineticin 2 in central nervous system autoimmunity. Neurol. Neuroimmunol. Neuroinflamm. 2015, 2, e95. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, D.; Sergent, F.; Abi Nahed, R.; Traboulsi, W.; Collet, C.; Marquette, C.; Hoffmann, P.; Balboni, G.; Zhou, Q.-Y.; Murthi, P.; et al. Evidence-Based View of Safety and Effectiveness of Prokineticin Receptors Antagonists during Pregnancy. Biomedicines 2021, 9, 309. [Google Scholar] [CrossRef]
- Goryszewska-Szczurek, E.; Baryla, M.; Kaczynski, P.; Waclawik, A. Prokineticin1–prokineticin receptor1signalingin trophoblast promotes embryo implantation and placenta development. Sci. Rep. 2019, 11, 13715. [Google Scholar] [CrossRef]
- Landucci, E.; Lattanzi, R.; Gerace, E.; Scartabelli, T.; Balboni, G.; Negri, L.; Pellegrini-Giampietro, D.E. Prokineticins are neuroprotective in models of cerebral ischemia and ischemic tolerance in vitro. Neuropharmacology 2016, 108, 39–48. [Google Scholar] [CrossRef]
- Curtis, V.F.; Wang, H.; Yang, P.; McLendon, R.E.; Li, X.; Zhou, Q.Y.; Wang, X.F. A PK2/Bv8/PROK2 Antagonist Suppresses Tumorigenic Processes by Inhibiting Angiogenesis in Glioma and Blocking Myeloid Cell Infiltration in Pancreatic Cancer. PLoS ONE 2013, 8, e54916. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Noda, K.; Yoshida, K.; Otani, K.; Yoshiga, M.; Oto, Y.; Saito, S.; Kurosaka, D. Prokineticin2 antagonist, PKRA7suppresses arthritis in mice with collagen-induced arthritis. BMC Musculoskelet. Disord. 2016, 17, 387. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Su, Y.; Zhou, T.; Hu, Z.; Wei, J.; Wang, W.; Liu, C.; Zhang, H.; Zhao, K. ActivationoftheNLRP3InflammasomePathwayby Prokineticin 2 in Testicular Macrophages of Uropathogenic Escherichia coli—Induced Orchitis. Front. Immunol. 2019, 14, 1872. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhang, H.; Liu, L.; Wang, J.; Ye, Z. PK2/PKR1 Signaling Regulates Bladder Function and Sensation in Rats with Cyclophosphamide-Induced Cystitis. Mediat. Inflamm. 2015, 2015, 289519. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, M.; Zhang, Y.; Cai, F.; Jiang, B.; Zha, W.; Yu, W. Metformin Ameliorates Diabetic Cardiomyopathy by Activating the PK2/PKR Pathway. Front. Physiol. 2020, 11, 425. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, S.; Zhao, Q.; Yang, Z.; Yan, X.; Li, C.; Zha, W.; Yu, W. Trehalose Ameliorates Diabetic Cardiomyopathy: Role of the PK2/PKR Pathway Oxidative. Med. Cell. Longev. 2021, 2021, 6779559. [Google Scholar] [CrossRef]
- Gordon, R.; Neal, M.L.; Luo, J.; Langley, M.R.; Harischandra, D.S.; Panicker, N.; Charli, A.; Jin, H.; Anantharam, V.; Woodruff, T.M.; et al. Prokineticin-2 upregulation during neuronal injury mediates a compensatory protective response against dopaminergic neuronal degeneration. Nat. Commun. 2016, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Schirinzi, T.; Maftei, D.; Pieri, M.; Bernardini, S.; Mercuri, N.B.; Lattanzi, R.; Severini, C. Increase Of Prokineticin-2 in Serum of Patients with Parkinson’s Disease. Mov. Disord. 2021, 36, 1031–1036. [Google Scholar] [CrossRef]
- Gasser, A.; Brogi, S.; Urayama, K.; Nishi, T.; Kurose, H.; Tafi, A.; Ribeiro, N.; Désaubry, L.; Nebigil, C.G. Discovery and cardioprotective effects of the first non-Peptide agonists of the G protein-coupled prokineticin receptor-1. PLoS ONE 2015, 10, e0121027. [Google Scholar] [CrossRef] [Green Version]
- Gasser, A.; Chen, Y.W.; Audebrand, A.; Daglayan, A.; Charavin, M.; Escoubet, B.; Karpov, P.; Tetko, I.; Chan, M.W.; Cardinale, D.; et al. Prokineticin receptor-1 signaling inhibits dose- and time dependent anthracycline-induced cardiovascular toxicity via myocardial and vascular protection. Cardio Oncol. 2019, 1, 84–102. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lattanzi, R.; Miele, R. Non-Peptide Agonists and Antagonists of the Prokineticin Receptors. Curr. Issues Mol. Biol. 2022, 44, 6323-6332. https://doi.org/10.3390/cimb44120431
Lattanzi R, Miele R. Non-Peptide Agonists and Antagonists of the Prokineticin Receptors. Current Issues in Molecular Biology. 2022; 44(12):6323-6332. https://doi.org/10.3390/cimb44120431
Chicago/Turabian StyleLattanzi, Roberta, and Rossella Miele. 2022. "Non-Peptide Agonists and Antagonists of the Prokineticin Receptors" Current Issues in Molecular Biology 44, no. 12: 6323-6332. https://doi.org/10.3390/cimb44120431
APA StyleLattanzi, R., & Miele, R. (2022). Non-Peptide Agonists and Antagonists of the Prokineticin Receptors. Current Issues in Molecular Biology, 44(12), 6323-6332. https://doi.org/10.3390/cimb44120431