
Propensity Score-Matched Analysis to Identify Pathways Associated with Loss of Sodium Iodide Symporter in Papillary Thyroid Cancer


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Acquisition
2.2. Propensity Score Matching
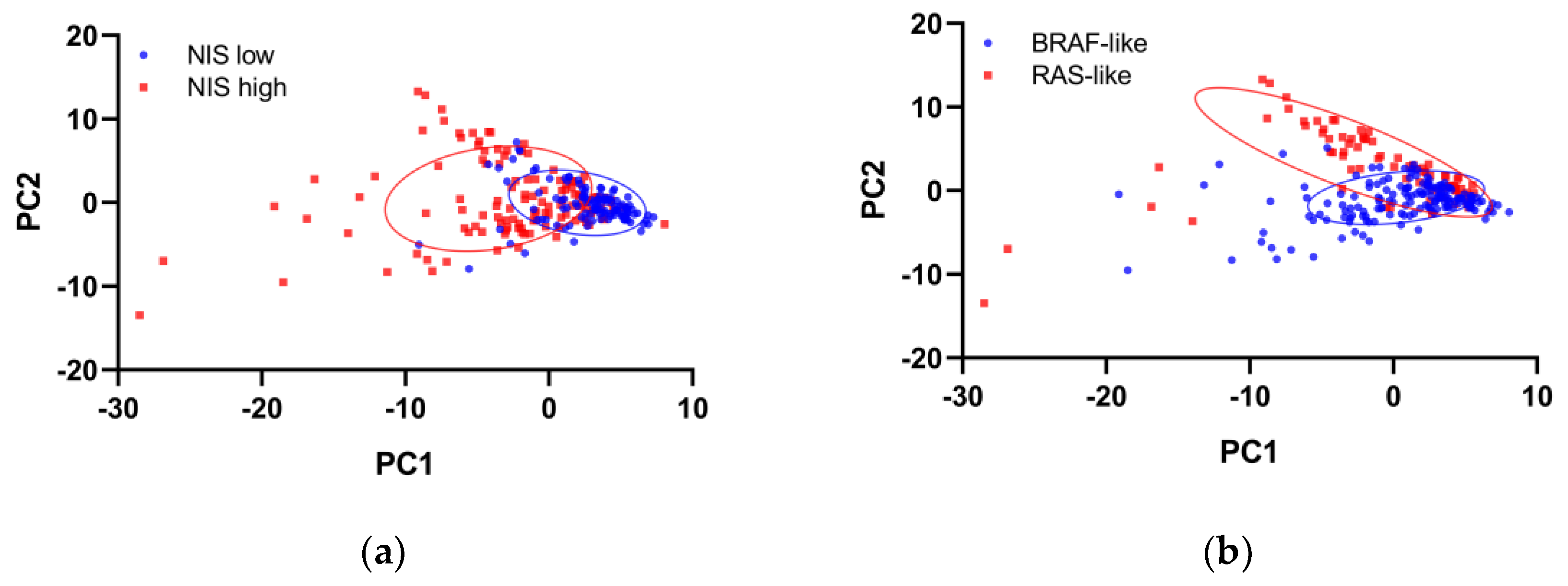
2.3. Principal Component Analysis
2.4. Pathway Analysis
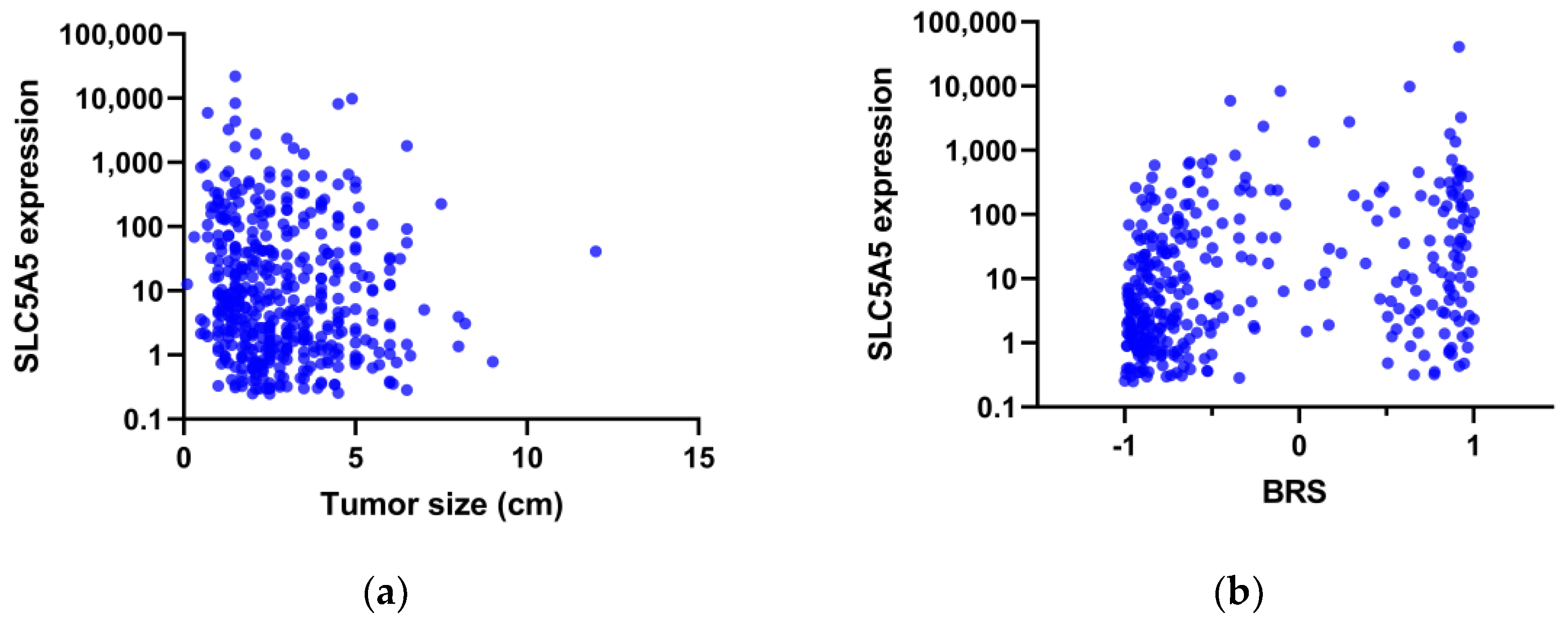
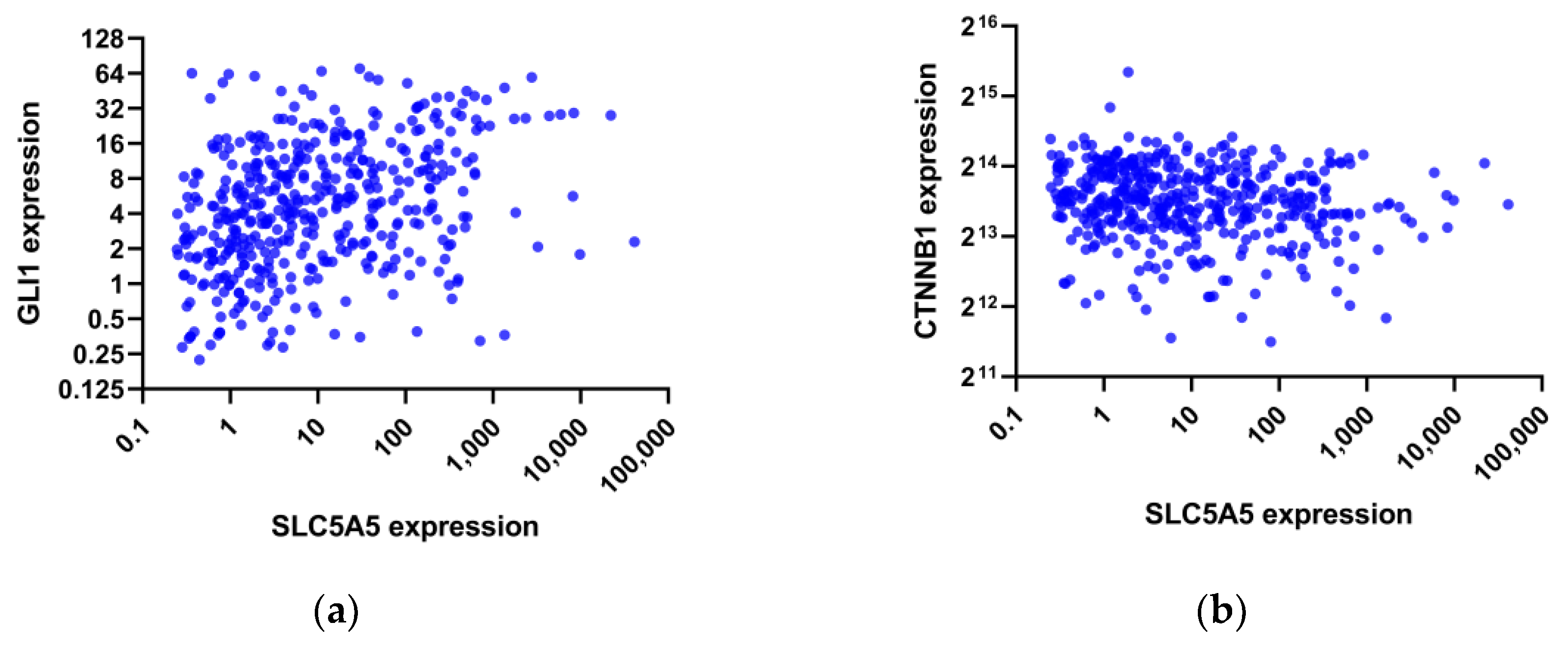
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Portulano, C.; Paroder-Belenitsky, M.; Carrasco, N. The Na+/I− symporter (NIS): Mechanism and medical impact. Endocr. Rev. 2014, 35, 106–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orosco, R.K.; Hussain, T.; Noel, J.E.; Chang, D.C.; Dosiou, C.; Mittra, E.; Divi, V.; Orloff, L.A. Radioactive iodine in differentiated thyroid cancer: A national database perspective. Endocr. Relat. Cancer 2019, 26, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Fugazzola, L.; Elisei, R.; Fuhrer, D.; Jarzab, B.; Leboulleux, S.; Newbold, K.; Smit, J. 2019 European Thyroid Association Guidelines for the Treatment and Follow-Up of Advanced Radioiodine-Refractory Thyroid Cancer. Eur. Thyroid J. 2019, 8, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Buffet, C.; Wassermann, J.; Hecht, F.; Leenhardt, L.; Dupuy, C.; Groussin, L.; Lussey-Lepoutre, C. Redifferentiation of radioiodine-refractory thyroid cancers. Endocr. Relat. Cancer 2020, 27, R113–R132. [Google Scholar] [CrossRef]
- Oh, J.M.; Ahn, B.C. Molecular mechanisms of radioactive iodine refractoriness in differentiated thyroid cancer: Impaired sodium iodide symporter (NIS) expression owing to altered signaling pathway activity and intracellular localization of NIS. Theranostics 2021, 11, 6251–6277. [Google Scholar] [CrossRef] [PubMed]
- de Morais, R.M.; Sobrinho, A.B.; de Souza Silva, C.M.; de Oliveira, J.R.; da Silva, I.C.R.; de Toledo Nobrega, O. The role of the NIS (SLC5A5) gene in papillary thyroid cancer: A systematic review. Int. J. Endocrinol. 2018, 2018, 9128754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothenberg, S.M.; McFadden, D.G.; Palmer, E.L.; Daniels, G.H.; Wirth, L.J. Redifferentiation of iodine-refractory BRAF V600E-mutant metastatic papillary thyroid cancer with dabrafenib. Clin. Cancer Res. 2015, 21, 1028–1035. [Google Scholar] [CrossRef] [Green Version]
- Chien, M.N.; Yang, P.S.; Hsu, Y.C.; Liu, T.P.; Lee, J.J.; Cheng, S.P. Transcriptome analysis of papillary thyroid cancer harboring telomerase reverse transcriptase promoter mutation. Head Neck 2018, 40, 2528–2537. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Integrated genomic characterization of papillary thyroid carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.P.; Lee, J.J.; Chang, Y.C.; Lin, C.H.; Li, Y.S.; Liu, C.L. Overexpression of chitinase-3-like protein 1 is associated with structural recurrence in patients with differentiated thyroid cancer. J. Pathol. 2020, 252, 114–124. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Lee, J.J.; Chien, M.N.; Chen, M.J.; Leung, C.H.; Cheng, S.P. Is papillary thyroid microcarcinoma a biologically different disease? A propensity score-matched analysis. J. Surg. Oncol. 2019, 120, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.H.; Lien, C.W.; Wu, W.C.; Lee, L.S.; Shaw, J.S. Artificial intelligence of neuropsychological tests for the prediction and verification of decline in gait parameters in patients with mild cognitive impairment. Int. J. Gerontol. 2020, 14, 277–283. [Google Scholar]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Jiang, H.; Xu, W.; Wang, X.; Ma, B.; Liao, T.; Wang, Y. Clinical, pathological, and molecular characteristics correlating to the occurrence of radioiodine refractory differentiated thyroid carcinoma: A systematic review and meta-analysis. Front. Oncol. 2020, 10, 549882. [Google Scholar] [CrossRef]
- Tavares, C.; Coelho, M.J.; Eloy, C.; Melo, M.; da Rocha, A.G.; Pestana, A.; Batista, R.; Ferreira, L.B.; Rios, E.; Selmi-Ruby, S.; et al. NIS expression in thyroid tumors, relation with prognosis clinicopathological and molecular features. Endocr. Connect. 2018, 7, 78–90. [Google Scholar] [CrossRef]
- Faggiano, A.; Coulot, J.; Bellon, N.; Talbot, M.; Caillou, B.; Ricard, M.; Bidart, J.M.; Schlumberger, M. Age-dependent variation of follicular size and expression of iodine transporters in human thyroid tissue. J. Nucl. Med. 2004, 45, 232–237. [Google Scholar]
- Mihailovic, J.; Stefanovic, L.; Malesevic, M.; Markoski, B. The importance of age over radioiodine avidity as a prognostic factor in differentiated thyroid carcinoma with distant metastases. Thyroid 2009, 19, 227–232. [Google Scholar] [CrossRef]
- Espadinha, C.; Santos, J.R.; Sobrinho, L.G.; Bugalho, M.J. Expression of iodine metabolism genes in human thyroid tissues: Evidence for age and BRAFV600E mutation dependency. Clin. Endocrinol. 2009, 70, 629–635. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Liu, C.L.; Yang, P.S.; Tsai, C.H.; Lee, J.J.; Cheng, S.P. Interaction of age at diagnosis with transcriptional profiling in papillary thyroid cancer. World J. Surg. 2016, 40, 2922–2929. [Google Scholar] [CrossRef]
- Chien, M.N.; Yang, P.S.; Lee, J.J.; Wang, T.Y.; Hsu, Y.C.; Cheng, S.P. Recurrence-associated genes in papillary thyroid cancer: An analysis of data from The Cancer Genome Atlas. Surgery 2017, 161, 1642–1650. [Google Scholar] [CrossRef]
- Ricarte-Filho, J.C.; Ryder, M.; Chitale, D.A.; Rivera, M.; Heguy, A.; Ladanyi, M.; Janakiraman, M.; Solit, D.; Knauf, J.A.; Tuttle, R.M.; et al. Mutational profile of advanced primary and metastatic radioactive iodine-refractory thyroid cancers reveals distinct pathogenetic roles for BRAF, PIK3CA, and AKT1. Cancer Res. 2009, 69, 4885–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiden, K.B.; Williamson, A.J.; Doscas, M.E.; Ye, J.; Wang, Y.; Liu, D.; Xing, M.; Prinz, R.A.; Xu, X. The sonic hedgehog signaling pathway maintains the cancer stem cell self-renewal of anaplastic thyroid cancer by inducing snail expression. J. Clin. Endocrinol. Metab. 2014, 99, E2178–E2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Jeong, S.; Lee, C.R.; Ku, C.R.; Kang, S.W.; Jeong, J.J.; Nam, K.H.; Shin, D.Y.; Chung, W.Y.; Lee, E.J.; et al. GLI1 transcription factor affects tumor aggressiveness in patients with papillary thyroid cancers. Medicine 2015, 94, e998. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.S.; Lee, J.J.; Li, Y.S.; Cheng, S.P. Ethacridine induces apoptosis and differentiation in thyroid cancer cells in vitro. Anticancer Res. 2019, 39, 4095–4100. [Google Scholar] [CrossRef]
- Romitti, M.; Wajner, S.M.; Ceolin, L.; Ferreira, C.V.; Ribeiro, R.V.; Rohenkohl, H.C.; Weber Sde, S.; Lopez, P.L.; Fuziwara, C.S.; Kimura, E.T.; et al. MAPK and SHH pathways modulate type 3 deiodinase expression in papillary thyroid carcinoma. Endocr. Relat. Cancer 2016, 23, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Lan, L.; Basourakos, S.; Cui, D.; Zuo, X.; Deng, W.; Huo, L.; Chen, L.; Zhang, G.; Deng, L.; Shi, B.; et al. Inhibiting beta-catenin expression promotes efficiency of radioiodine treatment in aggressive follicular thyroid cancer cells probably through mediating NIS localization. Oncol. Rep. 2017, 37, 426–434. [Google Scholar] [CrossRef]
- Sastre-Perona, A.; Santisteban, P. Wnt-independent role of beta-catenin in thyroid cell proliferation and differentiation. Mol. Endocrinol. 2014, 28, 681–695. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, P.M.; Adam, B.L.; Gourin, C.G.; Moretz WH 3rd Bollag, R.J.; Wang, B.Y.; Liu, Z.; Lee, J.R.; Terris, D.J. Association of nuclear, cytoplasmic expression of galectin-3 with beta-catenin/Wnt-pathway activation in thyroid carcinoma. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 503–510. [Google Scholar] [CrossRef]
- Lee, J.J.; Hsu, Y.C.; Li, Y.S.; Cheng, S.P. Galectin-3 inhibitors suppress anoikis resistance and invasive capacity in thyroid cancer cells. Int. J. Endocrinol. 2021, 2021, 5583491. [Google Scholar] [CrossRef]
- Feng, F.; Yehia, L.; Ni, Y.; Chang, Y.S.; Jhiang, S.M.; Eng, C. A nonpump function of sodium iodide symporter in thyroid cancer via cross-talk with PTEN signaling. Cancer Res. 2018, 78, 6121–6133. [Google Scholar] [CrossRef] [Green Version]
- Rakhsh-Khorshid, H.; Samimi, H.; Torabi, S.; Sajjadi-Jazi, S.M.; Samadi, H.; Ghafouri, F.; Asgari, Y.; Haghpanah, V. Network analysis reveals essential proteins that regulate sodium-iodide symporter expression in anaplastic thyroid carcinoma. Sci. Rep. 2020, 10, 21440. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wu, X.; Zhang, X.; Hua, W.; Zhang, Y.; Maimaiti, Y.; Gao, Z.; Zhang, Y. Inhibition of BRD4 suppresses tumor growth and enhances iodine uptake in thyroid cancer. Biochem. Biophys. Res. Commun. 2016, 469, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Lyckesvard, M.N.; Kapoor, N.; Ingeson-Carlsson, C.; Carlsson, T.; Karlsson, J.O.; Postgard, P.; Himmelman, J.; Forssell-Aronsson, E.; Hammarsten, O.; Nilsson, M. Linking loss of sodium-iodide symporter expression to DNA damage. Exp. Cell Res. 2016, 344, 120–131. [Google Scholar] [CrossRef]
- Guo, Y.; Li, H.; Guan, H.; Ke, W.; Liang, W.; Xiao, H.; Li, Y. Dermatopontin inhibits papillary thyroid cancer cell proliferation through MYC repression. Mol. Cell. Endocrinol. 2019, 480, 122–132. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Low NIS (n = 250) | High NIS (n = 250) | p Value | |
|---|---|---|---|
| Age (years) | 46 (35–58) | 46 (35–58) | 0.971 |
| Female | 180 (72%) | 185 (74%) | 0.614 |
| Subtype | 0.281 | ||
| Classic | 179 (72%) | 175 (70%) | |
| Follicular variant | 48 (19%) | 55 (22%) | |
| Tall cell variant | 21 (8%) | 14 (6%) | |
| Other variants | 2 (1%) | 6 (2%) | |
| Tumor size (cm) a | 2.7 (2.0–4.0) | 2.3 (1.5–3.7) | 0.002 |
| Multifocality a | 104 (42%) | 122 (50%) | 0.072 |
| Extrathyroidal extension a,b | 0.001 | ||
| None | 149 (62%) | 181 (75%) | |
| Minimal (T3) | 78 (32%) | 55 (23%) | |
| Advanced (T4) | 15 (6%) | 4 (2%) | |
| Lymph node metastasis | 122 (49%) | 102 (41%) | 0.072 |
| Stage b | 0.058 | ||
| Stage I | 131 (52%) | 149 (60%) | |
| Stage II | 27 (11%) | 22 (9%) | |
| Stage III | 52 (21%) | 57 (23%) | |
| Stage IV | 40 (16%) | 22 (9%) | |
| Risk of recurrence a,b | 0.001 | ||
| Low | 74 (32%) | 97 (44%) | |
| Intermediate | 136 (59%) | 121 (55%) | |
| High | 20 (9%) | 4 (2%) | |
| BRAF V600E mutation a | 146 (59%) | 88 (36%) | <0.001 |
| RAS mutation a | 23 (9%) | 29 (12%) | 0.387 |
| TERT promoter mutation a | 28 (14%) | 11 (6%) | 0.006 |
| Univariate | Multivariate | |||||
|---|---|---|---|---|---|---|
| Hazard Ratio | 95% CI | p Value | Hazard Ratio | 95% CI | p Value | |
| Age at diagnosis | 1.012 | 0.994–1.031 | 0.180 | |||
| Tumor size | 1.171 | 1.000–1.372 | 0.050 | 0.999 | 0.806–1.237 | 0.989 |
| Multifocality | 1.090 | 0.606–1.961 | 0.774 | |||
| Extrathyroidal extension | 1.817 | 1.010–3.266 | 0.046 | 0.796 | 0.360–1.758 | 0.572 |
| Lymph node metastasis | 1.917 | 1.060–3.466 | 0.031 | 1.339 | 0.909–1.972 | 0.139 |
| AJCC stage | 1.443 | 1.135–1.834 | 0.003 | 1.344 | 0.954–1.895 | 0.091 |
| BRAF V600E mutation | 1.450 | 0.797–2.638 | 0.223 | |||
| RAS mutation | 1.223 | 0.482–3.104 | 0.672 | |||
| TERT promoter mutation | 2.806 | 1.217–6.471 | 0.015 | 1.611 | 0.643–4.033 | 0.309 |
| Low NIS expression | 3.610 | 1.791–7.276 | <0.001 | 3.136 | 1.255–7.832 | 0.014 |
| Pathway | Enrichment Score | Adjusted p |
|---|---|---|
| KEGG_HEDGEHOG_SIGNALING_PATHWAY | −0.547 | 0.001 |
| KEGG_INTESTINAL_IMMUNE_NETWORK_FOR_IGA_PRODUCTION | −0.760 | 0.004 |
| KEGG_PRIMARY_IMMUNODEFICIENCY | −0.773 | 0.006 |
| KEGG_ASTHMA | −0.753 | 0.010 |
| KEGG_CYTOKINE_CYTOKINE_RECEPTOR_INTERACTION | −0.513 | 0.011 |
| KEGG_HEMATOPOIETIC_CELL_LINEAGE | −0.601 | 0.019 |
| KEGG_CHEMOKINE_SIGNALING_PATHWAY | −0.494 | 0.021 |
| KEGG_T_CELL_RECEPTOR_SIGNALING_PATHWAY | −0.484 | 0.021 |
| KEGG_CELL_ADHESION_MOLECULES_CAMS | −0.533 | 0.022 |
| KEGG_B_CELL_RECEPTOR_SIGNALING_PATHWAY | −0.493 | 0.024 |
| KEGG_COMPLEMENT_AND_COAGULATION_CASCADES | −0.498 | 0.031 |
| KEGG_GLYCOSAMINOGLYCAN_BIOSYNTHESIS_CHONDROITIN_SULFATE | −0.570 | 0.031 |
| KEGG_RIBOFLAVIN_METABOLISM | +0.542 | 0.035 |
| KEGG_ONE_CARBON_POOL_BY_FOLATE | +0.580 | 0.037 |
| KEGG_TYPE_I_DIABETES_MELLITUS | −0.690 | 0.040 |
| KEGG_AUTOIMMUNE_THYROID_DISEASE | −0.630 | 0.041 |
| KEGG_GRAFT_VERSUS_HOST_DISEASE | −0.734 | 0.047 |
| KEGG_NUCLEOTIDE_EXCISION_REPAIR | +0.458 | 0.047 |
| KEGG_ALLOGRAFT_REJECTION | −0.747 | 0.048 |
| KEGG_DNA_REPLICATION | +0.624 | 0.048 |
| Pathway | Enrichment Score | Adjusted p |
|---|---|---|
| PID_TCR_CALCIUM_PATHWAY | −0.668 | 0.001 |
| PID_MYC_PATHWAY | +0.521 | 0.004 |
| PID_CD8_TCR_PATHWAY | −0.644 | 0.018 |
| PID_IL12_STAT4_PATHWAY | −0.676 | 0.018 |
| PID_ALPHA_SYNUCLEIN_PATHWAY | −0.454 | 0.027 |
| PID_PI3KCI_PATHWAY | −0.524 | 0.027 |
| PID_LKB1_PATHWAY | +0.397 | 0.029 |
| PID_HIV_NEF_PATHWAY | −0.557 | 0.031 |
| PID_FRA_PATHWAY | −0.574 | 0.031 |
| PID_IL23_PATHWAY | −0.659 | 0.032 |
| PID_CD8_TCR_DOWNSTREAM_PATHWAY | −0.523 | 0.034 |
| PID_TCR_PATHWAY | −0.616 | 0.038 |
| PID_WNT_CANONICAL_PATHWAY | +0.519 | 0.047 |
| PID_BETA_CATENIN_DEG_PATHWAY | +0.502 | 0.048 |
| PID_ILK_PATHWAY | −0.420 | 0.050 |
| PID_CD40_PATHWAY | −0.493 | 0.053 |
| PID_NFAT_TFPATHWAY | −0.510 | 0.053 |
| PID_AP1_PATHWAY | −0.452 | 0.056 |
| PID_ERBB_NETWORK_PATHWAY | +0.572 | 0.056 |
| PID_THROMBIN_PAR4_PATHWAY | −0.577 | 0.062 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, F.; Kuo, C.-Y.; Tsai, C.-H.; Cheng, S.-P. Propensity Score-Matched Analysis to Identify Pathways Associated with Loss of Sodium Iodide Symporter in Papillary Thyroid Cancer. Curr. Issues Mol. Biol. 2022, 44, 1488-1496. https://doi.org/10.3390/cimb44040101
Lee F, Kuo C-Y, Tsai C-H, Cheng S-P. Propensity Score-Matched Analysis to Identify Pathways Associated with Loss of Sodium Iodide Symporter in Papillary Thyroid Cancer. Current Issues in Molecular Biology. 2022; 44(4):1488-1496. https://doi.org/10.3390/cimb44040101
Chicago/Turabian StyleLee, Fang, Chi-Yu Kuo, Chung-Hsin Tsai, and Shih-Ping Cheng. 2022. "Propensity Score-Matched Analysis to Identify Pathways Associated with Loss of Sodium Iodide Symporter in Papillary Thyroid Cancer" Current Issues in Molecular Biology 44, no. 4: 1488-1496. https://doi.org/10.3390/cimb44040101
APA StyleLee, F., Kuo, C.-Y., Tsai, C.-H., & Cheng, S.-P. (2022). Propensity Score-Matched Analysis to Identify Pathways Associated with Loss of Sodium Iodide Symporter in Papillary Thyroid Cancer. Current Issues in Molecular Biology, 44(4), 1488-1496. https://doi.org/10.3390/cimb44040101