
Calcium–Permeable Channels and Endothelial Dysfunction in Acute Lung Injury

Abstract
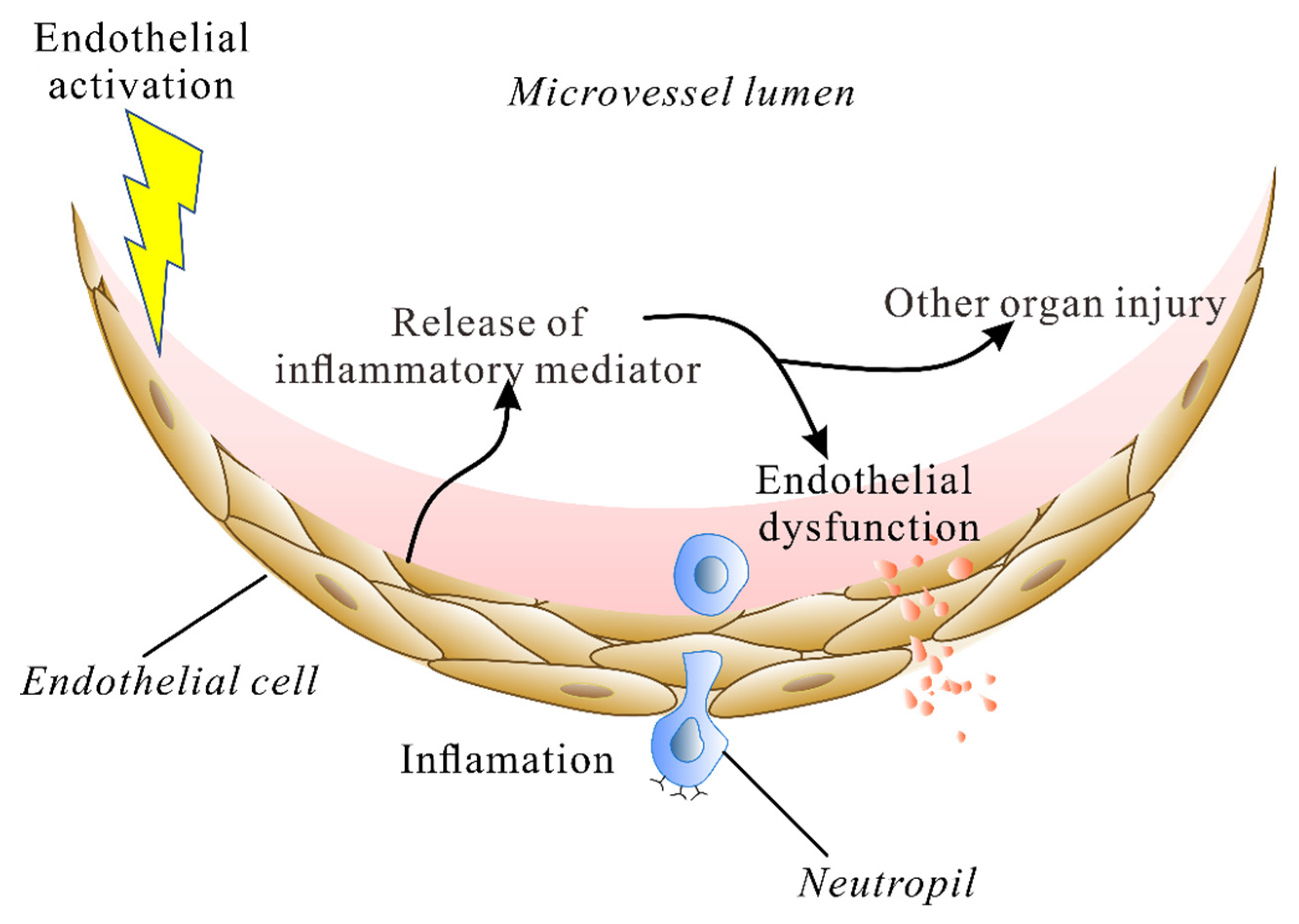
:1. Introduction
2. The Endothelium during Lung Injury
3. Calcium Influx into ECs Mediates Lung Injury
4. Calcium Machinery
4.1. Voltage-Gated Ca2+ Channels
4.2. Store-Operated Calcium (SOC) Channel
4.3. TRP Channels
4.4. Piezo Channel
4.5. Calcium Flux: A New Target for Therapy
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mowery, N.T.; Terzian, W.T.H.; Nelson, A.C. Acute lung injury. Curr. Probl. Surg. 2020, 57, 100777. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.W. Current and Future Direct-Acting Antivirals against COVID-19. Front. Microbiol. 2020, 11, 587944. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Cao, C.; Gao, Y.; Zhang, W.; Xie, Y.; Duan, Y.; Kong, S.; You, M.; Ma, R.; Jiang, L.; et al. Prognostic value of bedside lung ultrasound score in patients with COVID-19. Crit. Care 2020, 24, 700. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Y.; Zhang, C.; Huang, F.; Wang, F.; Yuan, J.; Wang, Z.; Li, J.; Li, J.; Feng, C.; et al. Clinical and biochemical indexes from 2019-nCoV infected patients linked to viral loads and lung injury. Sci. China Life Sci. 2020, 63, 364–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Escobar, A.; Vera-Vera, S.; Jurado-Roman, A.; Jimenez-Valero, S.; Galeote, G.; Moreno, R. Calcium Signaling Pathway Is Involved in the Shedding of ACE2 Catalytic Ectodomain: New Insights for Clinical and Therapeutic Applications of ACE2 for COVID-19. Biomolecules 2022, 12, 76. [Google Scholar] [CrossRef]
- Pirahanchi, Y.; Marsidi, J.L.; Brown, K.N. Physiology, Endothelial Derived Relaxation Factor; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Townsley, M.I. Permeability and calcium signaling in lung endothelium: Unpack the box. Pulm. Circ. 2018, 8, 2045893217738218. [Google Scholar] [CrossRef] [Green Version]
- Haywood, N.; Ta, H.Q.; Zhang, A.; Charles, E.J.; Rotar, E.; Noona, S.t.; Salmon, M.; Daneva, Z.; Sonkusare, S.K.; Laubach, V.E. Endothelial Transient Receptor Potential Vanilloid 4 Channels Mediate Lung Ischemia-Reperfusion Injury. Ann. Thorac. Surg. 2022, 113, 1256–1264. [Google Scholar] [CrossRef]
- Li, M.; Fang, X.Z.; Zheng, Y.F.; Xie, Y.B.; Ma, X.D.; Liu, X.T.; Xia, Y.; Shao, D.H. Transient receptor potential vanilloid 4 is a critical mediator in LPS mediated inflammation by mediating calcineurin/NFATc3 signaling. Biochem. Biophys. Res. Commun. 2019, 513, 1005–1012. [Google Scholar] [CrossRef]
- Liu, L.; Guo, M.; Lv, X.; Wang, Z.; Yang, J.; Li, Y.; Yu, F.; Wen, X.; Feng, L.; Zhou, T. Role of Transient Receptor Potential Vanilloid 4 in Vascular Function. Front. Mol. Biosci. 2021, 8, 677661. [Google Scholar] [CrossRef]
- Esquivel-Ruiz, S.; Gonzalez-Rodriguez, P.; Lorente, J.A.; Perez-Vizcaino, F.; Herrero, R.; Moreno, L. Extracellular Vesicles and Alveolar Epithelial-Capillary Barrier Disruption in Acute Respiratory Distress Syndrome: Pathophysiological Role and Therapeutic Potential. Front. Physiol. 2021, 12, 752287. [Google Scholar] [CrossRef]
- Birukova, A.A.; Tian, X.; Cokic, I.; Beckham, Y.; Gardel, M.L.; Birukov, K.G. Endothelial barrier disruption and recovery is controlled by substrate stiffness. Microvasc. Res. 2013, 87, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komarova, Y.; Malik, A.B. Regulation of endothelial permeability via paracellular and transcellular transport pathways. Annu. Rev. Physiol. 2010, 72, 463–493. [Google Scholar] [CrossRef] [PubMed]
- Grimsley-Myers, C.M.; Isaacson, R.H.; Cadwell, C.M.; Campos, J.; Hernandes, M.S.; Myers, K.R.; Seo, T.; Giang, W.; Griendling, K.K.; Kowalczyk, A.P. VE-cadherin endocytosis controls vascular integrity and patterning during development. J. Cell Biol. 2020, 219, e201909081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotoli, B.M.; Barilli, A.; Visigalli, R.; Ferrari, F.; Dall’Asta, V. Endothelial Cell Activation by SARS-CoV-2 Spike S1 Protein: A Crosstalk between Endothelium and Innate Immune Cells. Biomedicines 2021, 9, 1220. [Google Scholar] [CrossRef]
- Lee, W.; Choi, H.J.; Sim, H.; Choo, S.; Song, G.Y.; Bae, J.S. Barrier protective functions of hederacolchiside-E against HMGB1-mediated septic responses. Pharmacol. Res. 2021, 163, 105318. [Google Scholar] [CrossRef]
- Andonegui, G.; Zhou, H.; Bullard, D.; Kelly, M.M.; Mullaly, S.C.; McDonald, B.; Long, E.M.; Robbins, S.M.; Kubes, P. Mice that exclusively express TLR4 on endothelial cells can efficiently clear a lethal systemic Gram-negative bacterial infection. J. Clin. Investig. 2009, 119, 1921–1930. [Google Scholar] [CrossRef] [Green Version]
- Pirianov, G.; Waddington, S.N.; Lindstrom, T.M.; Terzidou, V.; Mehmet, H.; Bennett, P.R. The cyclopentenone 15-deoxy-delta 12,14-prostaglandin J(2) delays lipopolysaccharide-induced preterm delivery and reduces mortality in the newborn mouse. Endocrinology 2009, 150, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Kuldo, J.M.; Westra, J.; Asgeirsdottir, S.A.; Kok, R.J.; Oosterhuis, K.; Rots, M.G.; Schouten, J.P.; Limburg, P.C.; Molema, G. Differential effects of NF-κB and p38 MAPK inhibitors and combinations thereof on TNF-α- and IL-1β-induced proinflammatory status of endothelial cells in vitro. Am. J. Physiol. Cell Physiol. 2005, 289, C1229–C1239. [Google Scholar] [CrossRef]
- Rayees, S.; Joshi, J.C.; Tauseef, M.; Anwar, M.; Baweja, S.; Rochford, I.; Joshi, B.; Hollenberg, M.D.; Reddy, S.P.; Mehta, D. PAR2-Mediated cAMP Generation Suppresses TRPV4-Dependent Ca(2+) Signaling in Alveolar Macrophages to Resolve TLR4-Induced Inflammation. Cell Rep. 2019, 27, 793–805.e4. [Google Scholar] [CrossRef] [Green Version]
- Escue, R.; Kandasamy, K.; Parthasarathi, K. Thrombin Induces Inositol Trisphosphate-Mediated Spatially Extensive Responses in Lung Microvessels. Am. J. Pathol. 2017, 187, 921–935. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Ding, J.; Zhou, X.; Chen, G.; Liu, S.F. Divergent roles of endothelial NF-kappaB in multiple organ injury and bacterial clearance in mouse models of sepsis. J. Exp. Med. 2008, 205, 1303–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdecchia, P.; Cavallini, C.; Spanevello, A.; Angeli, F. COVID-19: ACE2centric Infective Disease? Hypertension 2020, 76, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Kar, M. Vascular Dysfunction and Its Cardiovascular Consequences During and after COVID-19 Infection: A Narrative Review. Vasc. Health Risk Manag. 2022, 18, 105–112. [Google Scholar] [CrossRef]
- Oxley, T.J.; Mocco, J.; Majidi, S.; Kellner, C.P.; Shoirah, H.; Singh, I.P.; De Leacy, R.A.; Shigematsu, T.; Ladner, T.R.; Yaeger, K.A.; et al. Large-Vessel Stroke as a Presenting Feature of COVID-19 in the Young. N. Engl. J. Med. 2020, 382, e60. [Google Scholar] [CrossRef] [PubMed]
- Colas-Algora, N.; Millan, J. How many cadherins do human endothelial cells express? Cell. Mol. Life Sci. 2019, 76, 1299–1317. [Google Scholar] [CrossRef] [PubMed]
- Duong, C.N.; Vestweber, D. Mechanisms Ensuring Endothelial Junction Integrity beyond VE-Cadherin. Front. Physiol. 2020, 11, 519. [Google Scholar] [CrossRef]
- Ichimura, H.; Parthasarathi, K.; Quadri, S.; Issekutz, A.C.; Bhattacharya, J. Mechano-oxidative coupling by mitochondria induces proinflammatory responses in lung venular capillaries. J. Clin. Investig. 2003, 111, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Beckett, T.; Bagher, P.; Garland, C.J.; Dora, K.A. VEGF-A inhibits agonist-mediated Ca(2+) responses and activation of IKCa channels in mouse resistance artery endothelial cells. J. Physiol. 2018, 596, 3553–3566. [Google Scholar] [CrossRef]
- White, L.E.; Cui, Y.; Shelak, C.M.; Lie, M.L.; Hassoun, H.T. Lung endothelial cell apoptosis during ischemic acute kidney injury. Shock 2012, 38, 320–327. [Google Scholar] [CrossRef]
- Richter, F.; Williams, S.K.; John, K.; Huber, C.; Vaslin, C.; Zanker, H.; Fairless, R.; Pichi, K.; Marhenke, S.; Vogel, A.; et al. The TNFR1 Antagonist Atrosimab Is Therapeutic in Mouse Models of Acute and Chronic Inflammation. Front. Immunol. 2021, 12, 705485. [Google Scholar] [CrossRef]
- Rohrmann, S.; Garmo, H.; Malmstrom, H.; Hammar, N.; Jungner, I.; Walldius, G.; Van Hemelrijck, M. Association between serum calcium concentration and risk of incident and fatal cardiovascular disease in the prospective AMORIS study. Atherosclerosis 2016, 251, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Kang, K. Serum calcium and phosphate concentrations and intracranial atherosclerosis. Atherosclerosis 2014, 232, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Voltage-gated calcium channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Haynes, J., Jr.; Taylor, J.T.; Obiako, B.O.; Stubbs, J.R.; Li, M.; Stevens, T. Cav3.1 (alpha1G) T-type Ca2+ channels mediate vaso-occlusion of sickled erythrocytes in lung microcirculation. Circ. Res. 2003, 93, 346–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Wu, S. T-type calcium channels in pulmonary vascular endothelium. Microcirculation 2006, 13, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Wang, X.; Wang, Y.; King, J.A.C.; Xie, P.; Wu, S. CaMK4 is a downstream effector of the alpha1G T-type calcium channel to determine the angiogenic potential of pulmonary microvascular endothelial cells. Am. J. Physiol. Cell Physiol. 2021, 321, C964–C977. [Google Scholar] [CrossRef]
- Wei, Z.; Manevich, Y.; Al-Mehdi, A.B.; Chatterjee, S.; Fisher, A.B. Ca2+ flux through voltage-gated channels with flow cessation in pulmonary microvascular endothelial cells. Microcirculation 2004, 11, 517–526. [Google Scholar] [CrossRef]
- Gilbert, G.; Courtois, A.; Dubois, M.; Cussac, L.A.; Ducret, T.; Lory, P.; Marthan, R.; Savineau, J.P.; Quignard, J.F. T-type voltage gated calcium channels are involved in endothelium-dependent relaxation of mice pulmonary artery. Biochem. Pharmacol. 2017, 138, 61–72. [Google Scholar] [CrossRef]
- Zheng, Z.; Chen, H.; Xie, P.; Dickerson, C.A.; King, J.A.C.; Alexeyev, M.F.; Wu, S. alpha1G T-type calcium channel determines the angiogenic potential of pulmonary microvascular endothelial cells. Am. J. Physiol. Cell. Physiol. 2019, 316, C353–C364. [Google Scholar] [CrossRef]
- Zhou, C.; Chen, H.; King, J.A.; Sellak, H.; Kuebler, W.M.; Yin, J.; Townsley, M.I.; Shin, H.S.; Wu, S. Alpha1G T-type calcium channel selectively regulates P-selectin surface expression in pulmonary capillary endothelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2010, 299, L86–L97. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Wu, W.; Jiang, S.; Wan, S.; Meng, D.; Wang, Z.; Zhang, J.; Wei, L.; Yu, P. Mibefradil and Flunarizine, Two T-Type Calcium Channel Inhibitors, Protect Mice against Lipopolysaccharide-Induced Acute Lung Injury. Mediat. Inflamm. 2020, 2020, 3691701. [Google Scholar] [CrossRef] [PubMed]
- Hogan, P.G.; Lewis, R.S.; Rao, A. Molecular basis of calcium signaling in lymphocytes: STIM and ORAI. Annu. Rev. Immunol. 2010, 28, 491–533. [Google Scholar] [CrossRef] [Green Version]
- Nwokonko, R.M.; Cai, X.; Loktionova, N.A.; Wang, Y.; Zhou, Y.; Gill, D.L. The STIM-Orai Pathway: Conformational Coupling between STIM and Orai in the Activation of Store-Operated Ca(2+) Entry. Adv. Exp. Med. Biol. 2017, 993, 83–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhirajan, R.K.; Meng, S.; Chandramoorthy, H.C.; Mallilankaraman, K.; Mancarella, S.; Gao, H.; Razmpour, R.; Yang, X.F.; Houser, S.R.; Chen, J.; et al. Blockade of NOX2 and STIM1 signaling limits lipopolysaccharide-induced vascular inflammation. J. Clin. Investig. 2013, 123, 887–902. [Google Scholar] [CrossRef]
- Seeley, E.J.; Rosenberg, P.; Matthay, M.A. Calcium flux and endothelial dysfunction during acute lung injury: A STIMulating target for therapy. J. Clin. Investig. 2013, 123, 1015–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, X.; Dong, K.; Sun, R. STIM1 Regulates Endothelial Calcium Overload and Cytokine Upregulation during Sepsis. J. Surg. Res. 2021, 263, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Liang, X.; Li, H.; Sun, R. LPS-induced vein endothelial cell injury and acute lung injury have Btk and Orai 1 to regulate SOC-mediated calcium influx. Int. Immunopharmacol. 2021, 90, 107039. [Google Scholar] [CrossRef]
- Shinde, A.V.; Motiani, R.K.; Zhang, X.; Abdullaev, I.F.; Adam, A.P.; Gonzalez-Cobos, J.C.; Zhang, W.; Matrougui, K.; Vincent, P.A.; Trebak, M. STIM1 controls endothelial barrier function independently of Orai1 and Ca2+ entry. Sci. Signal. 2013, 6, ra18. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Zhang, P.; Yang, J.; Zhang, Z.; Wang, H.; Guo, Y.; Liu, M. The protective effect of the flavonoid fraction of Abutilon theophrasti Medic. leaves on LPS-induced acute lung injury in mice via the NF-kappaB and MAPK signalling pathways. Biomed. Pharmacother. 2019, 109, 1024–1031. [Google Scholar] [CrossRef]
- Lin, T.; Luo, W.; Li, Z.; Zhang, L.; Zheng, X.; Mai, L.; Yang, W.; Guan, G.; Su, Z.; Liu, P.; et al. Rhamnocitrin extracted from Nervilia fordii inhibited vascular endothelial activation via miR-185/STIM-1/SOCE/NFATc3. Phytomedicine 2020, 79, 153350. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Song, X.; Liu, R.; Zhang, J.; Li, Z. The structure of TRPC ion channels. Cell Calcium 2019, 80, 25–28. [Google Scholar] [CrossRef]
- Hicks, G.A. TRP channels as therapeutic targets: Hot property, or time to cool down? Neurogastroenterol. Motil. 2006, 18, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sooch, G.; Demaree, I.S.; White, F.A.; Obukhov, A.G. Transient Receptor Potential Canonical (TRPC) Channels: Then and Now. Cells 2020, 9, 1983. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cao, R.; Zhong, W. Host Calcium Channels and Pumps in Viral Infections. Cells 2019, 9, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achanta, S.; Jordt, S.E. Transient receptor potential channels in pulmonary chemical injuries and as countermeasure targets. Ann. N. Y. Acad. Sci. 2020, 1480, 73–103. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Steinritz, D.; Gudermann, T. Transient receptor potential (TRP) channels as molecular targets in lung toxicology and associated diseases. Cell Calcium 2017, 67, 123–137. [Google Scholar] [CrossRef]
- Mehta, D.; Malik, A.B. Signaling mechanisms regulating endothelial permeability. Physiol. Rev. 2006, 86, 279–367. [Google Scholar] [CrossRef]
- Tauseef, M.; Farazuddin, M.; Sukriti, S.; Rajput, C.; Meyer, J.O.; Ramasamy, S.K.; Mehta, D. Transient receptor potential channel 1 maintains adherens junction plasticity by suppressing sphingosine kinase 1 expression to induce endothelial hyperpermeability. FASEB J. 2016, 30, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Weissmann, N.; Sydykov, A.; Kalwa, H.; Storch, U.; Fuchs, B.; Mederos y Schnitzler, M.; Brandes, R.P.; Grimminger, F.; Meissner, M.; Freichel, M.; et al. Activation of TRPC6 channels is essential for lung ischaemia-reperfusion induced oedema in mice. Nat. Commun. 2012, 3, 649. [Google Scholar] [CrossRef]
- Tauseef, M.; Knezevic, N.; Chava, K.R.; Smith, M.; Sukriti, S.; Gianaris, N.; Obukhov, A.G.; Vogel, S.M.; Schraufnagel, D.E.; Dietrich, A.; et al. TLR4 activation of TRPC6-dependent calcium signaling mediates endotoxin-induced lung vascular permeability and inflammation. J. Exp. Med. 2012, 209, 1953–1968. [Google Scholar] [CrossRef]
- Rajan, S.; Schremmer, C.; Weber, J.; Alt, P.; Geiger, F.; Dietrich, A. Ca(2+) Signaling by TRPV4 Channels in Respiratory Function and Disease. Cells 2021, 10, 822. [Google Scholar] [CrossRef] [PubMed]
- Jian, M.Y.; King, J.A.; Al-Mehdi, A.B.; Liedtke, W.; Townsley, M.I. High vascular pressure-induced lung injury requires P450 epoxygenase-dependent activation of TRPV4. Am. J. Respir. Cell Mol. Biol. 2008, 38, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Michalick, L.; Tang, C.; Tabuchi, A.; Goldenberg, N.; Dan, Q.; Awwad, K.; Wang, L.; Erfinanda, L.; Nouailles, G.; et al. Role of Transient Receptor Potential Vanilloid 4 in Neutrophil Activation and Acute Lung Injury. Am. J. Respir. Cell Mol. Biol. 2016, 54, 370–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T.; Nilius, B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 2003, 424, 434–438. [Google Scholar] [CrossRef]
- Watanabe, H.; Vriens, J.; Janssens, A.; Wondergem, R.; Droogmans, G.; Nilius, B. Modulation of TRPV4 gating by intra- and extracellular Ca2+. Cell Calcium 2003, 33, 489–495. [Google Scholar] [CrossRef]
- Willette, R.N.; Bao, W.; Nerurkar, S.; Yue, T.L.; Doe, C.P.; Stankus, G.; Turner, G.H.; Ju, H.; Thomas, H.; Fishman, C.E.; et al. Systemic activation of the transient receptor potential vanilloid subtype 4 channel causes endothelial failure and circulatory collapse: Part 2. J. Pharmacol. Exp. Ther. 2008, 326, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, D.F.; King, J.A.; Weber, D.; Addison, E.; Liedtke, W.; Townsley, M.I. Transient receptor potential vanilloid 4-mediated disruption of the alveolar septal barrier: A novel mechanism of acute lung injury. Circ. Res. 2006, 99, 988–995. [Google Scholar] [CrossRef]
- Baratchi, S.; Keov, P.; Darby, W.G.; Lai, A.; Khoshmanesh, K.; Thurgood, P.; Vahidi, P.; Ejendal, K.; McIntyre, P. The TRPV4 Agonist GSK1016790A Regulates the Membrane Expression of TRPV4 Channels. Front. Pharmacol. 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Bihari, S.; Dixon, D.L.; Lawrence, M.D.; De Bellis, D.; Bonder, C.S.; Dimasi, D.P.; Bersten, A.D. Fluid-induced lung injury-role of TRPV4 channels. Pflugers Arch. 2017, 469, 1121–1134. [Google Scholar] [CrossRef]
- Suresh, K.; Servinsky, L.; Jiang, H.; Bigham, Z.; Yun, X.; Kliment, C.; Huetsch, J.; Damarla, M.; Shimoda, L.A. Reactive oxygen species induced Ca(2+) influx via TRPV4 and microvascular endothelial dysfunction in the SU5416/hypoxia model of pulmonary arterial hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L893–L907. [Google Scholar] [CrossRef] [Green Version]
- Balakrishna, S.; Song, W.; Achanta, S.; Doran, S.F.; Liu, B.; Kaelberer, M.M.; Yu, Z.; Sui, A.; Cheung, M.; Leishman, E.; et al. TRPV4 inhibition counteracts edema and inflammation and improves pulmonary function and oxygen saturation in chemically induced acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L158–L172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swain, S.M.; Liddle, R.A. Piezo1 acts upstream of TRPV4 to induce pathological changes in endothelial cells due to shear stress. J. Biol. Chem. 2021, 296, 100171. [Google Scholar] [CrossRef] [PubMed]
- Lawhorn, B.G.; Brnardic, E.J.; Behm, D.J. TRPV4 antagonists: A patent review (2015–2020). Expert Opin. Ther. Pat. 2021, 31, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Pairet, N.; Mang, S.; Fois, G.; Keck, M.; Kuhnbach, M.; Gindele, J.; Frick, M.; Dietl, P.; Lamb, D.J. TRPV4 inhibition attenuates stretch-induced inflammatory cellular responses and lung barrier dysfunction during mechanical ventilation. PLoS ONE 2018, 13, e0196055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorneloe, K.S.; Cheung, M.; Bao, W.; Alsaid, H.; Lenhard, S.; Jian, M.Y.; Costell, M.; Maniscalco-Hauk, K.; Krawiec, J.A.; Olzinski, A.; et al. An orally active TRPV4 channel blocker prevents and resolves pulmonary edema induced by heart failure. Sci. Transl. Med. 2012, 4, 159ra148. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, A.; Desjardins, J.; Shabbir, W.; Roy, A.; Filion, D.; Sauve, R.; Berthiaume, Y. Loss of barrier integrity in alveolar epithelial cells downregulates ENaC expression and activity via Ca(2+) and TRPV4 activation. Pflugers Arch. 2018, 470, 1615–1631. [Google Scholar] [CrossRef]
- Weber, J.; Rajan, S.; Schremmer, C.; Chao, Y.K.; Krasteva-Christ, G.; Kannler, M.; Yildirim, A.O.; Brosien, M.; Schredelseker, J.; Weissmann, N.; et al. TRPV4 channels are essential for alveolar epithelial barrier function as protection from lung edema. JCI Insight 2020, 5, e134464. [Google Scholar] [CrossRef]
- Kuebler, W.M.; Jordt, S.E.; Liedtke, W.B. Urgent reconsideration of lung edema as a preventable outcome in COVID-19: Inhibition of TRPV4 represents a promising and feasible approach. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L1239–L1243. [Google Scholar] [CrossRef]
- Cheung, M.; Bao, W.; Behm, D.J.; Brooks, C.A.; Bury, M.J.; Dowdell, S.E.; Eidam, H.S.; Fox, R.M.; Goodman, K.B.; Holt, D.A.; et al. Discovery of GSK2193874: An Orally Active, Potent, and Selective Blocker of Transient Receptor Potential Vanilloid 4. ACS Med. Chem. Lett. 2017, 8, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Brooks, C.A.; Barton, L.S.; Behm, D.J.; Eidam, H.S.; Fox, R.M.; Hammond, M.; Hoang, T.H.; Holt, D.A.; Hilfiker, M.A.; Lawhorn, B.G.; et al. Discovery of GSK2798745: A Clinical Candidate for Inhibition of Transient Receptor Potential Vanilloid 4 (TRPV4). ACS Med. Chem. Lett. 2019, 10, 1228–1233. [Google Scholar] [CrossRef]
- Goyal, N.; Skrdla, P.; Schroyer, R.; Kumar, S.; Fernando, D.; Oughton, A.; Norton, N.; Sprecher, D.L.; Cheriyan, J. Clinical Pharmacokinetics, Safety, and Tolerability of a Novel, First-in-Class TRPV4 Ion Channel Inhibitor, GSK2798745, in Healthy and Heart Failure Subjects. Am. J. Cardiovasc. Drugs 2019, 19, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mole, S.; Harry, A.; Fowler, A.; Hotee, S.; Warburton, J.; Waite, S.; Beerahee, M.; Behm, D.J.; Badorrek, P.; Muller, M.; et al. Investigating the effect of TRPV4 inhibition on pulmonary-vascular barrier permeability following segmental endotoxin challenge. Pulm. Pharmacol. Ther. 2020, 64, 101977. [Google Scholar] [CrossRef] [PubMed]
- Hecquet, C.M.; Ahmmed, G.U.; Malik, A.B. TRPM2 channel regulates endothelial barrier function. Adv. Exp. Med. Biol. 2010, 661, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Hecquet, C.M.; Zhang, M.; Mittal, M.; Vogel, S.M.; Di, A.; Gao, X.; Bonini, M.G.; Malik, A.B. Cooperative interaction of trp melastatin channel transient receptor potential (TRPM2) with its splice variant TRPM2 short variant is essential for endothelial cell apoptosis. Circ. Res. 2014, 114, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wang, L.; Moreno-Vinasco, L.; Lang, G.D.; Siegler, J.H.; Mathew, B.; Usatyuk, P.V.; Samet, J.M.; Geyh, A.S.; Breysse, P.N.; et al. Particulate matter air pollution disrupts endothelial cell barrier via calpain-mediated tight junction protein degradation. Part Fibre Toxicol. 2012, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; He, T.; Chen, S.; Yang, D.; Yi, W.; Cao, H.; Xiao, G. Roles of mechanosensitive channel Piezo1/2 proteins in skeleton and other tissues. Bone Res. 2021, 9, 44. [Google Scholar] [CrossRef]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, E.E.; Hong, Z.; Xiong, S.; Zhong, M.; Di, A.; Rehman, J.; Komarova, Y.A.; Malik, A.B. Endothelial cell Piezo1 mediates pressure-induced lung vascular hyperpermeability via disruption of adherens junctions. Proc. Natl. Acad. Sci. USA 2019, 116, 12980–12985. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jiang, L.; Huang, T.; Lu, D.; Song, Y.; Wang, L.; Gao, J. Mechanosensitive cation channel Piezo1 contributes to ventilator-induced lung injury by activating RhoA/ROCK1 in rats. Respir. Res. 2021, 22, 250. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, Y.; Lu, D.; Huang, T.; Yan, K.; Yang, W.; Gao, J. Mechanosensitive Piezo1 channel activation promotes ventilator-induced lung injury via disruption of endothelial junctions in ARDS rats. Biochem. Biophys. Res. Commun. 2021, 556, 79–86. [Google Scholar] [CrossRef]
- Zhong, M.; Wu, W.; Kang, H.; Hong, Z.; Xiong, S.; Gao, X.; Rehman, J.; Komarova, Y.A.; Malik, A.B. Alveolar Stretch Activation of Endothelial Piezo1 Protects Adherens Junctions and Lung Vascular Barrier. Am. J. Respir. Cell. Mol. Biol. 2020, 62, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.W.; Zhang, H.; Huang, J.Q.; Wang, S.N.; Lu, Y.; Cheng, B.; Dong, S.H.; Wang, Y.Y.; Li, F.S.; Li, Y.W. PIEZO1 Ion Channel Mediates Ionizing Radiation-Induced Pulmonary Endothelial Cell Ferroptosis via Ca(2+)/Calpain/VE-Cadherin Signaling. Front. Mol. Biosci. 2021, 8, 725274. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Target Channel | Compounds | Application | Reference |
|---|---|---|---|
| CaV3.1 | Mibefradil | LPS-induced lung injury | [36,42] |
| Flunarizine | LPS-induced lung injury | [36,42] | |
| SOCs | Rhamnocitrin | A potent inhibitor of endothelial activation | [51] |
| TRPV4 | GSK2798745 | LPS-induced lung inflammation * | [77,84] |
| GSK2193874 | High-pulmonary-venous-pressure (PVP)-induced edema | [77] | |
| HC-067047 | LPS-induced mouse lung injury | [9] | |
| Piezo1 | GSMTx4 | Ionizing-radiation (IR)-induced lung injury | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Y.; Wang, Z.; Frimpong, F.; Chen, X. Calcium–Permeable Channels and Endothelial Dysfunction in Acute Lung Injury. Curr. Issues Mol. Biol. 2022, 44, 2217-2229. https://doi.org/10.3390/cimb44050150
Hao Y, Wang Z, Frimpong F, Chen X. Calcium–Permeable Channels and Endothelial Dysfunction in Acute Lung Injury. Current Issues in Molecular Biology. 2022; 44(5):2217-2229. https://doi.org/10.3390/cimb44050150
Chicago/Turabian StyleHao, Ying, Zhuang Wang, Francis Frimpong, and Xingjuan Chen. 2022. "Calcium–Permeable Channels and Endothelial Dysfunction in Acute Lung Injury" Current Issues in Molecular Biology 44, no. 5: 2217-2229. https://doi.org/10.3390/cimb44050150