
Characterization of Insulin-like Peptide (ILP) and Its Potential Role in Ovarian Development of the Cuttlefish Sepiella japonica

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
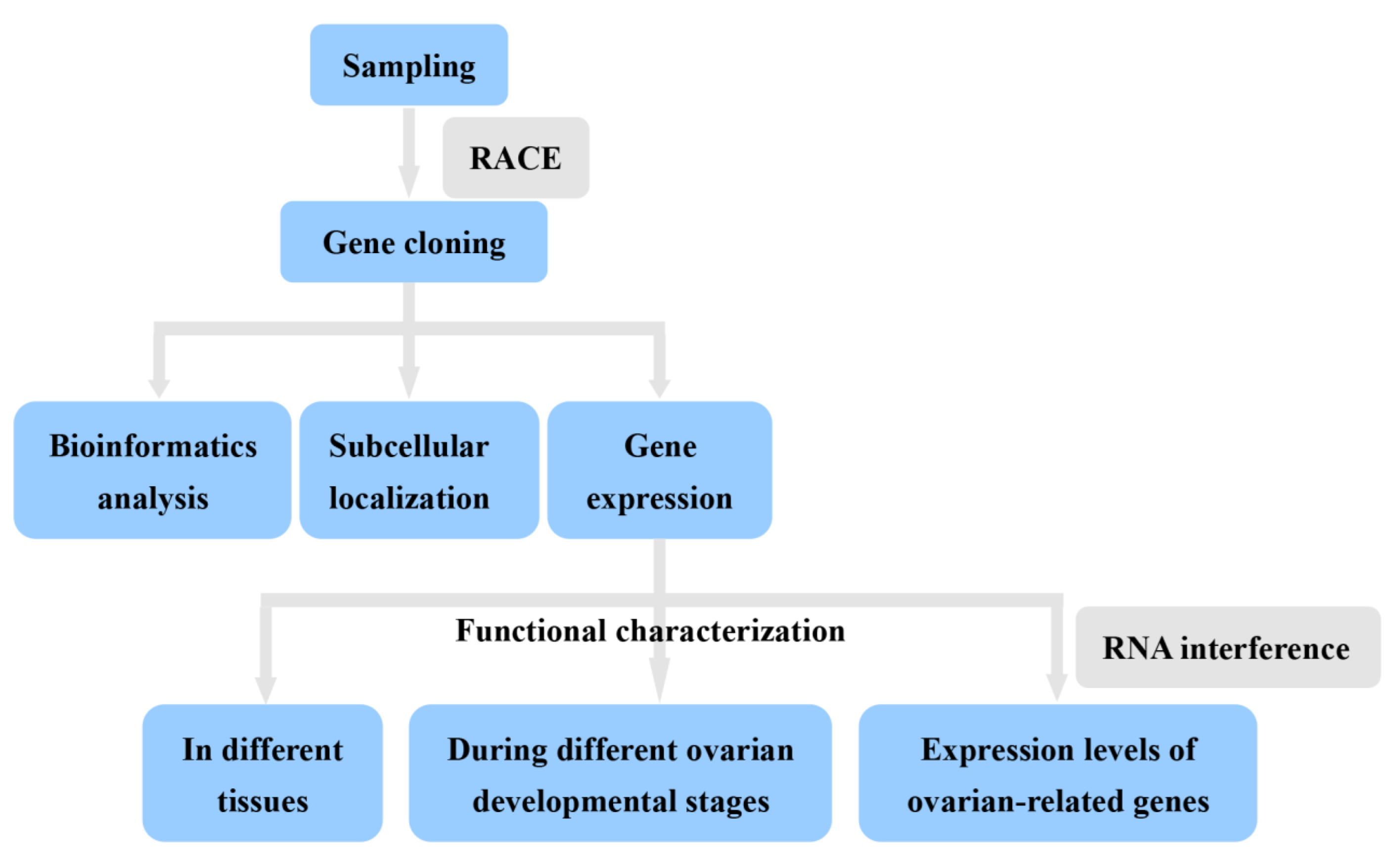
2. Materials and Methods
2.1. Animals
2.2. Cloning of SjILP
2.3. Bioinformatics Analysis
2.4. Subcellular Localization of SjILP
2.5. Expression of SjILP in Different Tissues
2.6. Expression of SjILP in Different Ovarian Development Stages
2.7. RNA Interference In Vivo
2.7.1. Synthesis and Injection of dsRNA
2.7.2. Functional Analysis of SjILP with Gene Silencing
2.8. Statistic Analysis
3. Results
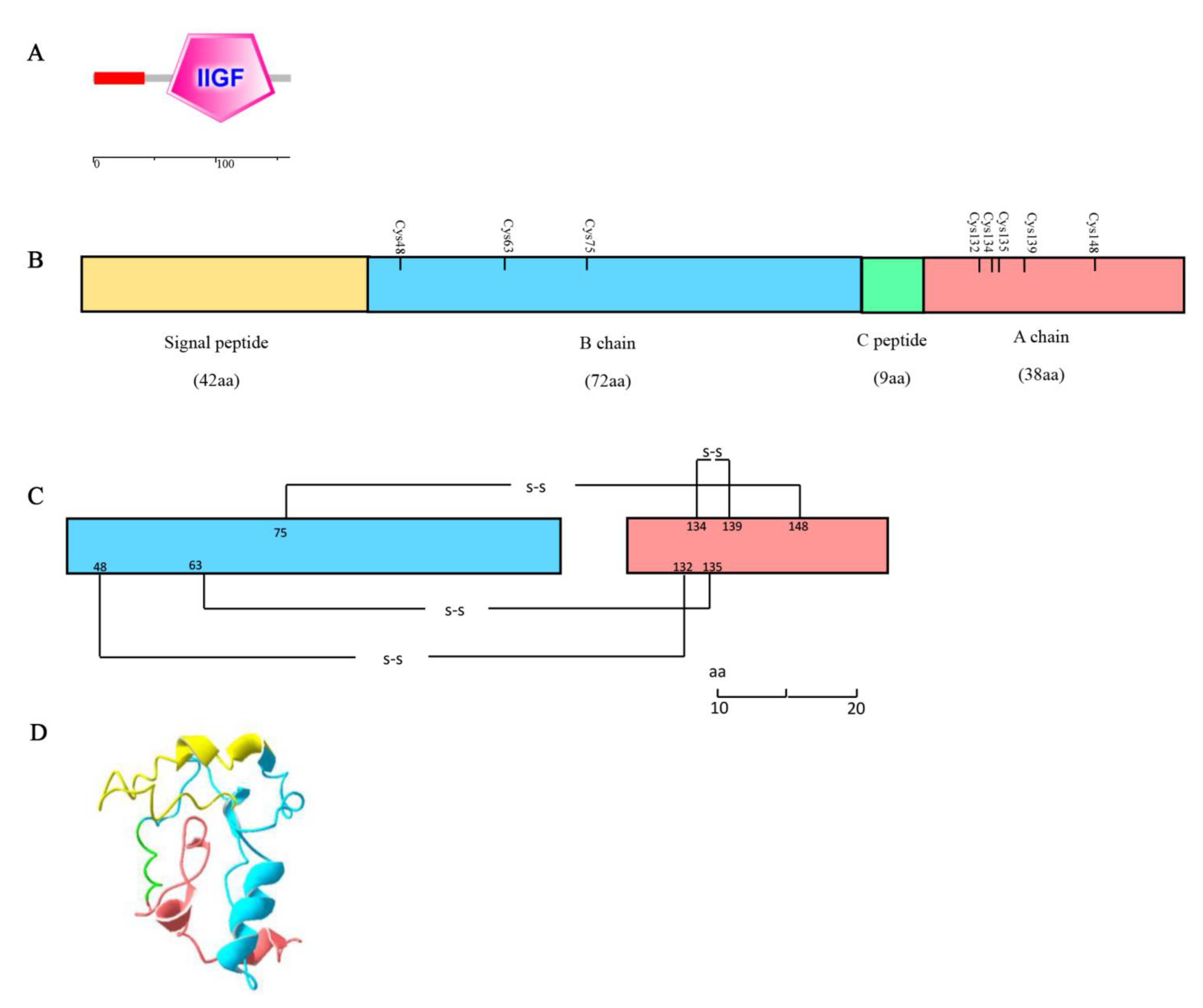
3.1. Characterization of SjILP Sequence
3.2. Multiple Alignment and Phylogenetic Analysis
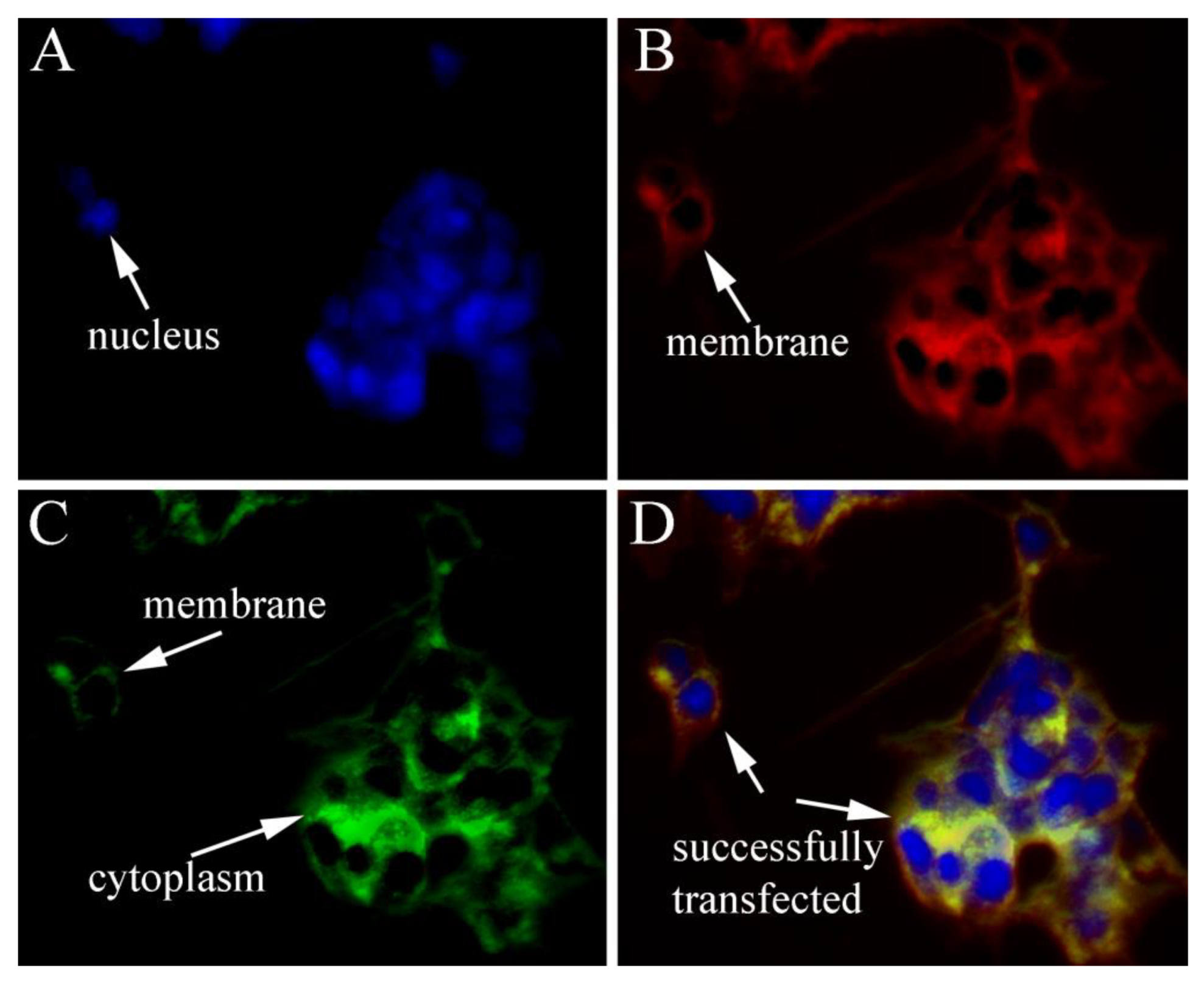
3.3. Subcellular Localization of SjILP
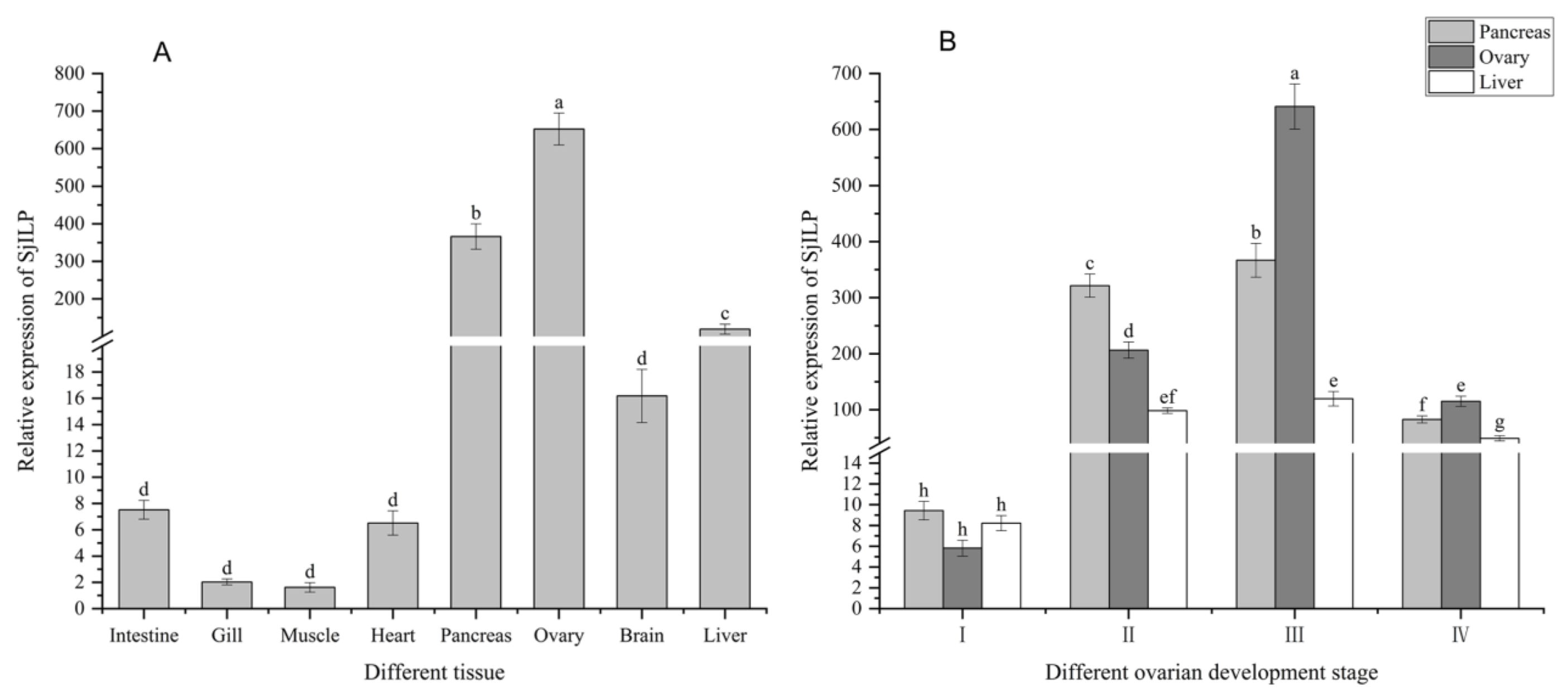
3.4. Expression Profile of SjILP Transcript in Different Tissues
3.5. Expression Profile of SjILP Transcript at Different Ovarian Developmental Stages
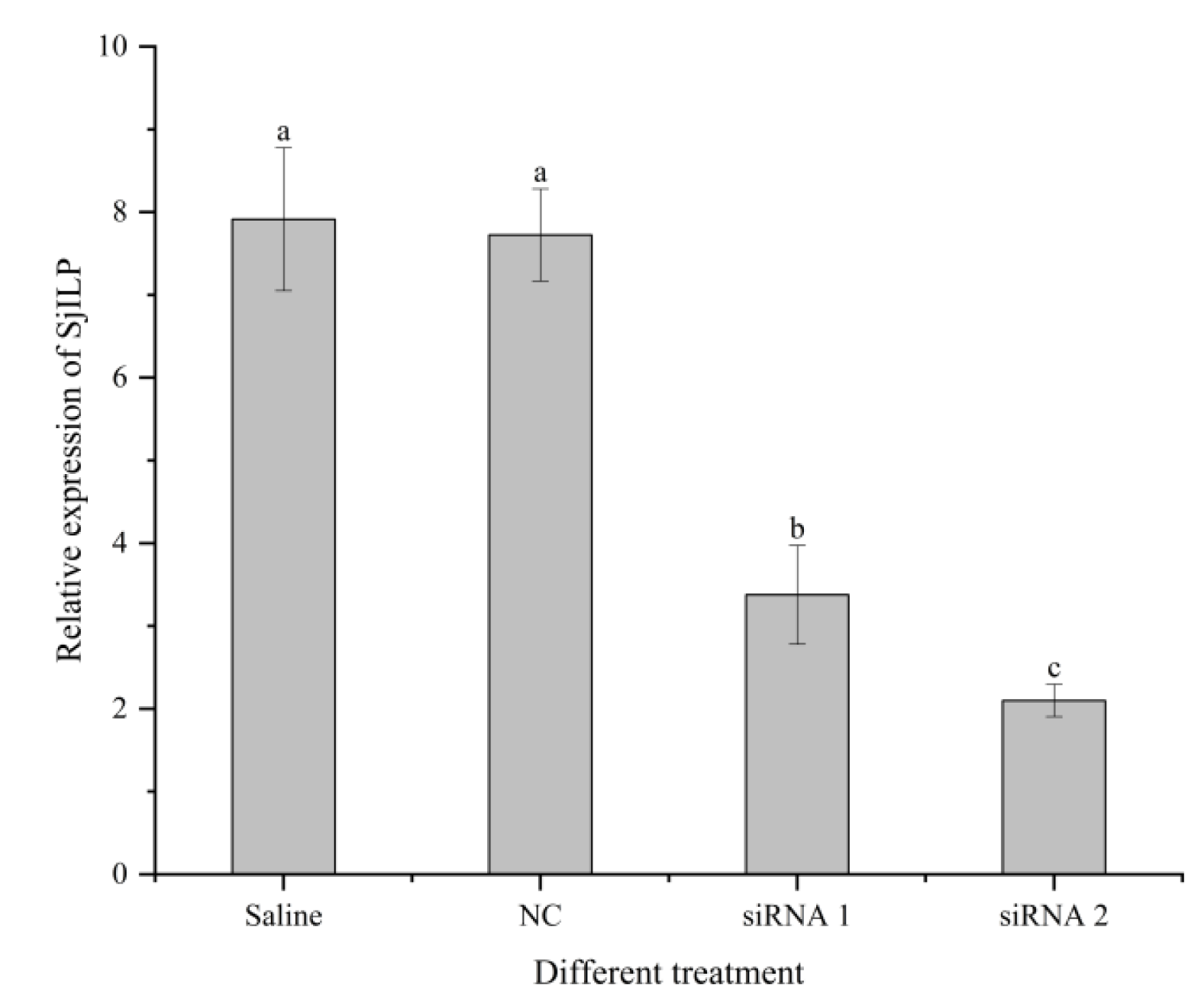
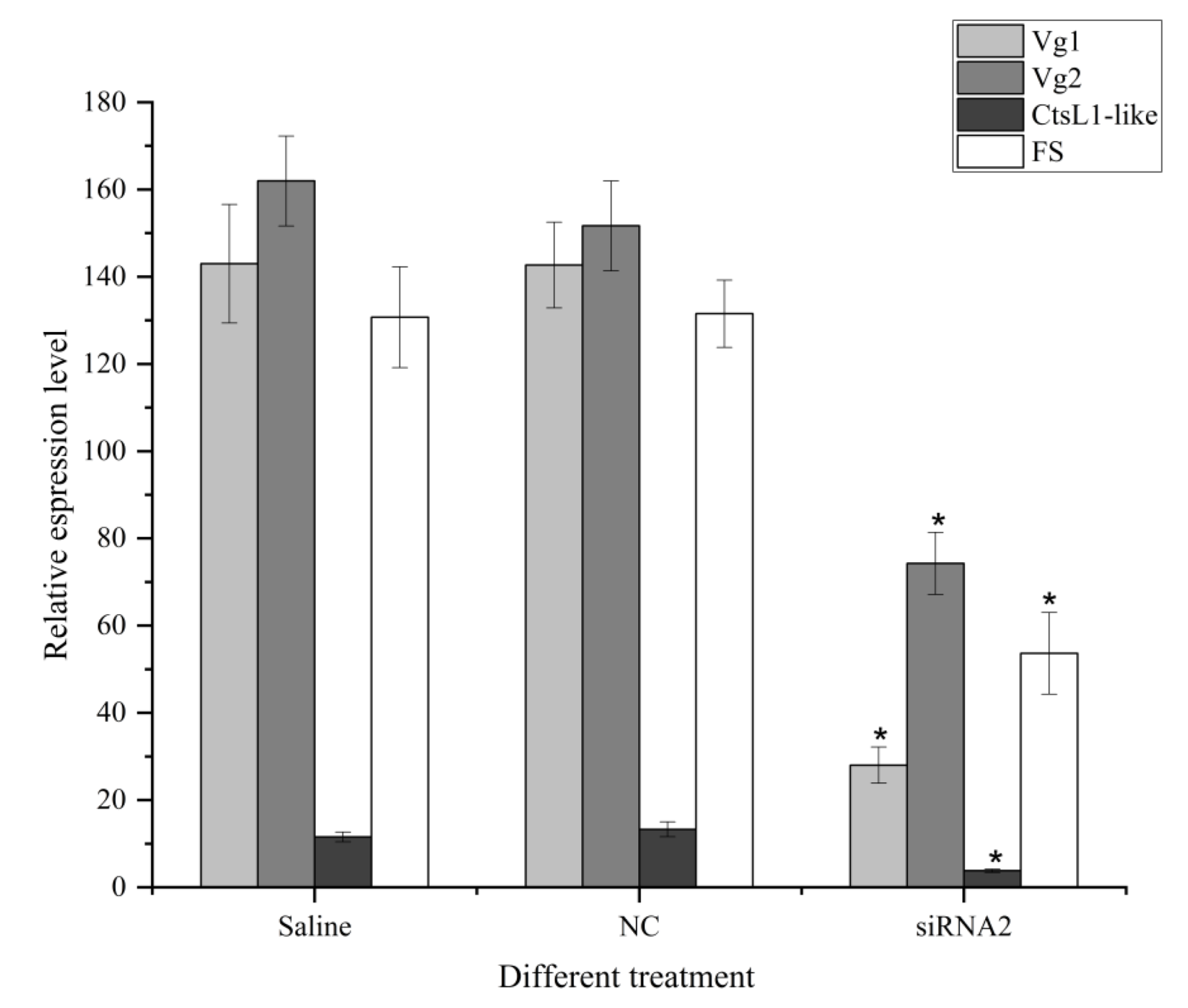
3.6. Effects of SjILP Silencing on Ovarian Development
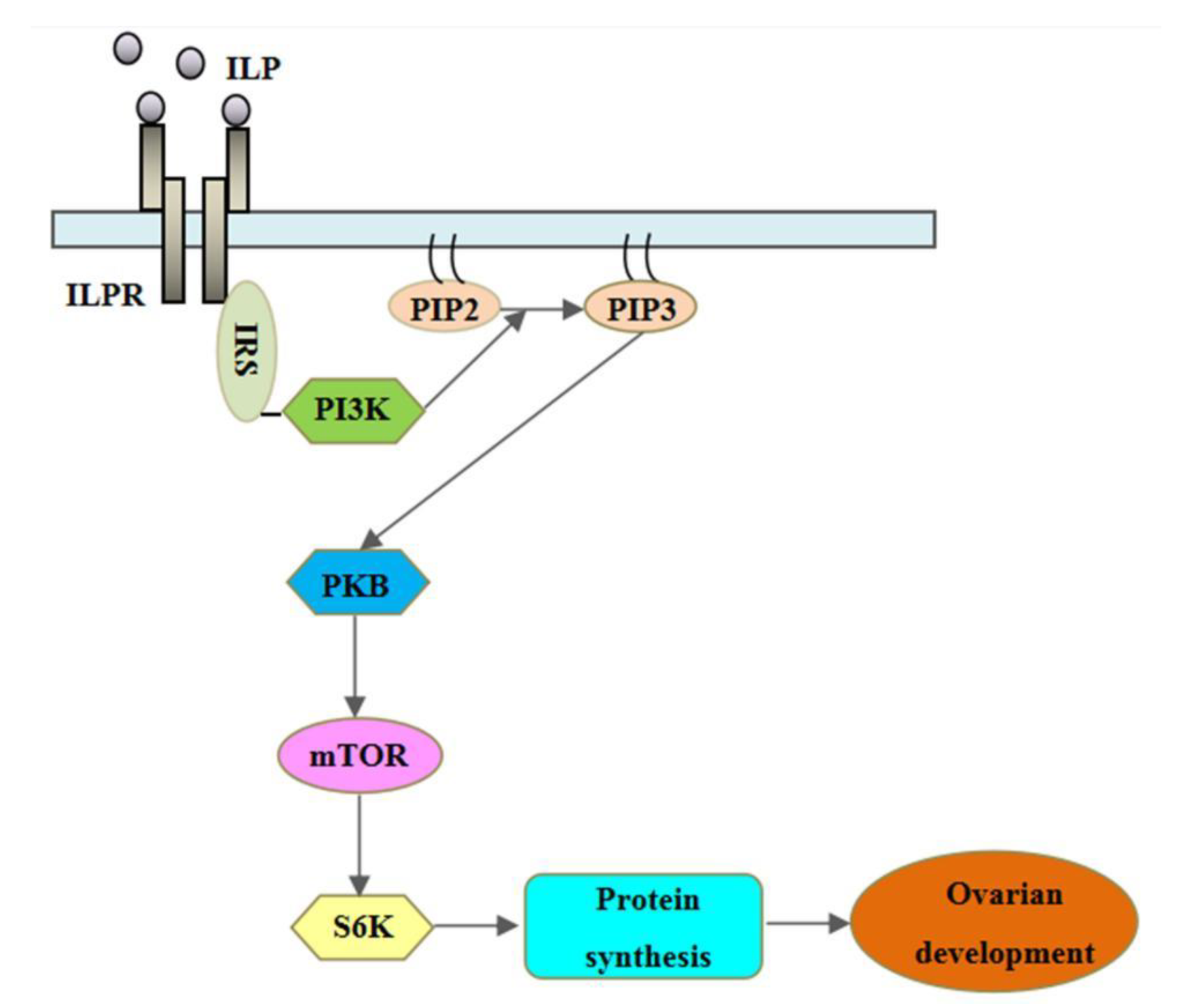
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okamoto, N. Molluscan insulin-related peptides. In Handbook of Hormones; Academic Press: Cambridge, MA, USA, 2021; pp. 681–683. [Google Scholar]
- Smit, A.B.; Vreugdenhil, E.; Ebberink, R.H.; Geraerts, W.P.; Klootwijk, J.; Joosse, J. Growth-controlling molluscan neurons produce the precursor of an insulin-related peptide. Nature 1988, 331, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Floyd, P.D.; Li, L.J.; Rubakhin, S.S.; Sweedler, J.V.; Horn, C.C.; Kupfermann, I.; Alexeeva, V.; Ellis, T.A.; Dembrow, N.C. Insulin prohormone processing, distribution, and relation to metabolism in Aplysia Californica. J. Neurosci. 1999, 19, 7732–7741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamano, K.; Awaji, M.; Usuki, H. cDNA structure of an insulin-related peptide in the Pacific oyster and seasonal changes in the gene expression. J. Endocrinol. 2005, 187, 55–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherif-Feildel, M.; Berthelin, C.H.; Adeline, B.; Rivière, G.; Favrel, P.; Kellner, K. Molecular evolution and functional characterisation of insulin related peptides in molluscs: Contributions of Crassostrea gigas genomic and transcriptomic-wide screening. Gen. Comp. Endocrinol. 2019, 271, 15–29. [Google Scholar] [CrossRef]
- Smit, A.B.; Spijker, S.; Van Minnen, J.; Burke, J.F.; De Winter, F.; Van Elk, R.; Geraerts, W.P. Expression and characterization of molluscan insulin-related peptide VII from the mollusc Lymnaea stagnalis. Neuroscience 1996, 70, 589–596. [Google Scholar] [CrossRef]
- Zhang, H.; He, M.X. The role of a new insulin-like peptide in the pearl oyster pinctada fucata martensii. Sci. Rep. 2020, 10, 433. [Google Scholar] [CrossRef] [Green Version]
- Plisetskaya, E.V.K.; Kazakov, V.K.; Soltitskaya, L.; Leibson, L.G. Insulin producing cells in the gut of freshwater bivalve Molluscs Anodonfa Cygnaea and Unio pictorum and the role of insulin in the regulation of their carbohydrate metabolism. Gen. Camp. Endocrinol. 1978, 35, 133–145. [Google Scholar] [CrossRef]
- Abdraba, A.M.; Saleuddin, A.S.M. Localization and immunological characterization of insulin-like peptide(s) in the land snail Otala lactea (Mollusca:Pulmonata). Can. J. Zool. 2000, 78, 1515–1526. [Google Scholar] [CrossRef]
- Li, Y.J.; Fu, H.R.; Zhang, F.Q.; Ren, L.T.; Tian, J.; Li, Q.; Liu, S.K. Identification, characterization, and expression profiles of insulin-like peptides suggest their critical roles in growth regulation of the pacific oyster, crassostrea gigas. Gene 2020, 769, 145244. [Google Scholar] [CrossRef]
- Niu, D.; Wang, F.; Zhao, H.; Wang, Z.; Xie, S.; Li, J. Identification, expression, and innate immune responses of two insulin-like peptide genes in the razor clam sinonovacula constricta. Fish Shellfish. Immun. 2016, 51, 401–404. [Google Scholar] [CrossRef]
- Zhang, L.H.; Wang, G.D. Insulin-like peptides in invertebrates and their signaling pathways-take insects. Biotechnol. Bull. 2014, 20, 33–42. [Google Scholar]
- Gricourt, L.; Mathieu, M.; Kellner, K. An insulin-like system involved in the control of pacific oyster Crassostrea gigas reproduction: hrIGF-1 effect on germinal cell proliferation and maturation associated with expression of an homologous insulin receptor-related receptor. Aquaculture 2006, 251, 85–98. [Google Scholar] [CrossRef]
- Sonetti, D.; Van Heumen, W.R.A.; Roubos, E.W. Light-and electro-microscopic immunocytochemistry of a molluscan insulin-related peptide in the central nervous system of Planorbarius corneus. Cell Tissue Res. 1992, 267, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Geraerts, W.P.M. The role of the lateral lobes in the control of growth and reproduction in the hermaphrodite freshwater snail Lymnaea stagnalis. Gen. Comp. Endocrinol 1976, 29, 97–108. [Google Scholar] [CrossRef]
- Monnier, Z.; Bride, M. In vitro effects of methionine-en-kephalin, somatostatin and insulin on cultured gonadal cells of the snail Helix aspersa. Experientia 1995, 51, 824–829. [Google Scholar] [CrossRef]
- Brown, M.R.; Clark, K.D.; Gulia, M.; Zhao, Z.W.; Garczynski, S.F.; Crim, J.W.; Suderman, R.J.; Strand, M.R. An insulin-like peptide regulates egg maturation and metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2008, 105, 5716–5721. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.Y.; Wu, C.W.; Ye, D.Y. A Fluorescent Method for Marking the Cuttlefish, Sepiella mainddroni de Rochebrune. J. Zhejiang Ocean. Univ. (Nat. Sci.) 2010, 29, 120–128. [Google Scholar]
- Lü, Z.M.; Liu, W.; Liu, L.Q.; Shi, H.L.; Ping, H.L.; Wang, T.M.; Chi, C.F.; Wu, C.W.; Chen, C.H.; Shen, K.N.; et al. De novo assembly and comparison of the ovarian transcriptomes of the common Chinese cuttlefish (Sepiella japonica) with different gonadal development. Genom. Data 2016, 7, 155–158. [Google Scholar] [CrossRef]
- Lü, Z.M.; Liu, W.; Liu, L.Q.; Wang, T.M.; Shi, H.L.; Ping, H.L.; Chi, C.F.; Yang, J.W.; Wu, C.W. Cloning, characterization, and expression profile of estrogen receptor in common Chinese cuttlefish, Sepiella japonica. J. Exp. Zool. Part A 2016, 325A, 181–193. [Google Scholar] [CrossRef]
- Pang, Z.; Lü, Z.M.; Wang, M.T.; Gong, L.; Liu, B.J.; Liu, L.Q. Characterization, relative abundances of mrna transcripts, and subcellular localization of two forms of membrane progestin receptors (MPRs) in the common chinese cuttlefish, sepiella japonica. Anim. Reprod. Sci. 2019, 208, 106107. [Google Scholar] [CrossRef]
- Lü, Z.M.; Zhu, K.H.; Pang, Z.; Liu, L.Q.; Jiang, H.; Liu, B.J.; Shi, H.L.; Ping, H.L.; Chi, C.F.; Gong, L. Identification, characterization and mrna transcript abundance profiles of estrogen related receptor (ERR) in Sepiella japonica imply its possible involvement in female reproduction. Anim. Reprod. Sci. 2019, 211, 106231. [Google Scholar] [CrossRef] [PubMed]
- Gomot, P. Studies on the control of spermatogenic DNA synthesis by the mesocerebrum in the snail Helix aspersa. Cell. Mol. Neurobiol. 1993, 13, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.X.; Freddolino, P.L.; Zhang, Y. COFACTOR: Improved protein function prediction by combining structure, sequence and protein-protein interaction information. Nucleic Acids Res. 2017, 45, 291–299. [Google Scholar] [CrossRef]
- Zheng, W.; Zhang, C.X.; Li, Y.; Pearce, R.; Bell, E.W.; Zhang, Y. Folding non-homology proteins by coupling deep-learning contact maps with I-TASSER assembly simulations. Cell Rep. Methods 2021, 1, 100014. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- Kumar, S.; Dudley, J.; Nei, M.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsvik, P.A.; Lie, K.K.; Jordal, A.E.O.; Nilsen, T.O.; Hordvik, I. Evaluation of potential reference genes in real-time RT-PCR studies of Atlantic salmon. BMC Mol. Biol. 2005, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gore, S.R.; Harms, C.A.; Kukanich, B.; Forsythe, J.; Lewbart, G.A.; Papich, M.G. Enrofloxacin pharmacokinetics in the European cuttlefish, Sepia officinalis, after a single iv injection and bath administration. J. Vet. Pharmacol. Ther. 2005, 28, 433–439. [Google Scholar] [CrossRef]
- Kimura-Kawakami, M.; Iwami, M.; Kawakami, A.; Nagasawa, H.; Suzuki, A.; Ishizaki, H. Structure and expression of bombyxin-related peptide genes of the moth Samia cynthia ricini. Gen. Comp. Endocr. 1992, 86, 257–268. [Google Scholar] [CrossRef]
- Krieger, M.J.B.; Jahan, N.; Riehle, M.A.; Cao, C.; Brown, M.R. Molecular characterization of insulin-like peptide genes and their expression in the african malaria mosquito, Anopheles gambiae. Insect Mol. Biol. 2010, 13, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Okuno, A.; Hasegawa, Y.; Nishiyama, M.; Ohira, T.; Ko, R.; Kurihara, M.; Matsumoto, S.; Nagasawa, H. Preparation of an active recombinant peptide of crustacean androgenic gland hormone. Peptides 2002, 23, 567–572. [Google Scholar] [CrossRef]
- Mareddy, V.R.; Rosen, O.; Thaggard, H.B.; Manor, R.; Kuballa, A.V.; Aflalo, E.D.; Sagi, A.; Paterson, B.; Elizur, A. Isolation and characterization of the complete cdna sequence encoding a putative insulin-like peptide from the androgenic gland of penaeus monodon. Aquaculture 2011, 318, 364–370. [Google Scholar] [CrossRef]
- Velde, S.; Badisco, L.; Claeys, I.; Verleyen, P.; Chen, X.; Bosch, L.V.; Broeck, J.V.; Smagghe, G. Insulin-like peptides in Spodoptera littoralis (Lepidoptera): Detection, localization and identification. Gen. Comp. Endocr. 2007, 153, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.Y.; Lin, J.Y.; Guo, S.Z.; Chen, Y.; Li, J.L.; Qiu, G.F. Molecular characterization and expression analysis of an insulin-like gene from the androgenic gland of the oriental river prawn, macrobrachium nipponense. Gen. Comp. Endocr. 2013, 185, 90–96. [Google Scholar] [CrossRef]
- Smit, A.B.; Kesteren, R.E.V.; Li, K.W.; Minnen, J.V.; Spijker, S.; Heerikhuizen, H.V.; Geraerts, W.P.M. Towards understanding the role of insulin in the brain: Lessons from insulin-related signaling systems in the invertebrate brain. Prog. Neurobiol. 1998, 54, 35–54. [Google Scholar] [CrossRef]
- Iwami, M.; Furuya, I.; Kataoka, H. Bombyxin-related peptides: cDNA structure and expression in the brain of the hornworm Agrius convolvuli. Insect. Biochem. Mol. Biol. 1996, 26, 25–32. [Google Scholar] [CrossRef]
- Orikasa, C.; Yamauchi, H.; Nagasawa, H.; Suzuki, A.; Nagata, M. Induction of oocyte-nurse cell differentiation in the ovary by the brain during the initial stage of oogenesis in the silkworm, Bombyx mori (Lepidoptera: Bombycidae). Appl. Entomol. Zool. 1993, 28, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Luo, X.N.; Zhang, S.H.; Yin, C.; Dou, Y.X.; Cai, X.P. Identification of putative insulin-like peptides and components of insulin signaling pathways in parasitic platyhelminths by the use of genome-wide screening. Febs J. 2014, 2818, 77–893. [Google Scholar] [CrossRef]
- Geraerts, W.P.M. Neurohormonal control of growth and carbohydrate metabolism by the light green cells in Lymnaea stagnalis. Gen. Comp. Endocrinol. 1992, 86, 433–444. [Google Scholar] [CrossRef]
- Meester, I.; Ramkema, M.D.; Van Minnen, J.; Boer, H.H. Differential expression of four genes encoding molluscan insu-lin-related peptides in the central nervous system of the pond snail Lymmea stagnalis. Cell Tissue Res. 1992, 269, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Kellner-Cousin, K.M.E.; Mathieu, M. Identification of insulin-like peptides in cerebral ganglia neurosecretory cells in the mussel Mytilus Edulis. Tissue Cell. 1994, 26, 891–899. [Google Scholar] [CrossRef]
- Danton, E.; Kiyomoto, M.; Komaru, A.; Wada, K.T.; Awaji, M.; Mathieu, M. Comparative analysis of storage tissue and insulin-like neurosecretion in diploid and triploid mussels Mytilus galloprovincialis LMK in relation to their gametogenesis cycle. Invertebr. Reprod. Dev. 1996, 29, 37–46. [Google Scholar] [CrossRef]
- Gómez-Mendikute, A.; Elizondo, M.; Venier, P.; Cajaraville, M.P. Characterization of mussel gill cells in vivo and in vitro. Cell. Tissue Res. 2005, 321, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.S.; Rybczynski, R.; Wilson, T.G.; Wang, Y.; Wayne, M.L.; Zhou, Y.; Partridge, L.; Harshman, L.G. Insulin signaling is necessary for vitellogenesis in Drosophila melanogaster independent of the roles of juvenile hormone and ecdysteroids: Female sterility of the chico1 insulin signaling mutation is autonomous to the ovary. J. Insect Physiol. 2005, 51, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Gricourt, L.; Bonnec, G.; Boujard, D.; Mathieu, M.; Kellner, K. Insulin-like system and growth regulation in the Pacific oyster Crassostrea gigas: hrIGF-1 effect on protein synthesis of mantle edge cells and expression of an homologous insulin receptor-related receptor. Gen. Comp. Endocrinol. 2003, 134, 44–56. [Google Scholar] [CrossRef]
- Chen, C.; Li, H.W.; Ku, W.L.; Lin, C.J.; Chang, C.F.; Wu, G.C. Two distinct vitellogenin genes are similar in function and expression in the bigfin reef squid sepioteuthis lessoniana. Biol. Reprod. 2018, 5, 5. [Google Scholar] [CrossRef]
- Sheng, Z.; Xu, J.; Bai, H.; Zhu, F.; Palli, S.R. Juvenile hormone regulates vitellogenin gene expression through insulin-like peptide signaling pathway in the red flour beetle, tribolium castaneum. J. Biol. Chem. 2011, 286, 41924–41936. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.T.; Mccarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Hao, L.; Cynthia, K. Genes that act downstream of daf-16 to influence the lifespan of caenorhabditis elegans. Nature 2003, 424, 277. [Google Scholar] [CrossRef]
- Wuertz, S.; Nitsche, A.; Jastroch, M.; Gessner, J.; Klingenspor, M.; Kirschbaum, F.; Kloas, W. The role of the IGF-i system for vitellogenesis in maturing female sterlet, 1758. Acipenser ruthenus linnaeus. Gen. Comp. Endocrinol. 2007, 150, 140–150. [Google Scholar] [CrossRef]
- Badisco, L.; Marchal, E.; Wielendaele, P.V.; Verlinden, H.; Vleugels, R.; Broeck, J.V. RNA interference of insulin-related peptide and neuroparsins affects vitellogenesis in the desert locust schistocerca gregaria. Peptides 2011, 32, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Tingaud-Sequeira, A.; Cerdà, J. Phylogenetic relationships and gene expression pattern of three different cathepsin L. (Ctsl) isoforms in zebrafish: Ctsla is the putative yolk processing enzyme. Gene 2007, 386, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Fu, H.; Qiao, H.; Jin, S.; Xiong, Y. Expression and functional analysis of cathepsin L1 in ovarian development of the oriental river prawn, macrobrachium nipponense. Aquacult. Rep. 2021, 20, 100724. [Google Scholar] [CrossRef]
- Britton, C.; Murray, L. Cathepsin l protease (CPL-1) is essential for yolk processing during embryogenesis in caenorhabditis elegans. J. Cell. Sci. 2004, 117, 5133–5143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashimada, K.; Pelosi, E.; Chen, H.; Schlessinger, D.; Wilhelm, D.; Koopman, P. FOXL2 and BMP2 act cooperatively to regulate follistatin gene expression during ovarian development. Endocrinology 2011, 152, 272–280. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lü, Z.; Yao, C.; Zhao, S.; Zhang, Y.; Gong, L.; Liu, B.; Liu, L. Characterization of Insulin-like Peptide (ILP) and Its Potential Role in Ovarian Development of the Cuttlefish Sepiella japonica. Curr. Issues Mol. Biol. 2022, 44, 2490-2504. https://doi.org/10.3390/cimb44060170
Lü Z, Yao C, Zhao S, Zhang Y, Gong L, Liu B, Liu L. Characterization of Insulin-like Peptide (ILP) and Its Potential Role in Ovarian Development of the Cuttlefish Sepiella japonica. Current Issues in Molecular Biology. 2022; 44(6):2490-2504. https://doi.org/10.3390/cimb44060170
Chicago/Turabian StyleLü, Zhenming, Chenghao Yao, Shijie Zhao, Yao Zhang, Li Gong, Bingjian Liu, and Liqin Liu. 2022. "Characterization of Insulin-like Peptide (ILP) and Its Potential Role in Ovarian Development of the Cuttlefish Sepiella japonica" Current Issues in Molecular Biology 44, no. 6: 2490-2504. https://doi.org/10.3390/cimb44060170
APA StyleLü, Z., Yao, C., Zhao, S., Zhang, Y., Gong, L., Liu, B., & Liu, L. (2022). Characterization of Insulin-like Peptide (ILP) and Its Potential Role in Ovarian Development of the Cuttlefish Sepiella japonica. Current Issues in Molecular Biology, 44(6), 2490-2504. https://doi.org/10.3390/cimb44060170