
Comparative Transcriptome Analysis of Softening and Ripening-Related Genes in Kiwifruit Cultivars Treated with Ethylene


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Ethylene Treatment
2.2. Transcriptome Sequencing
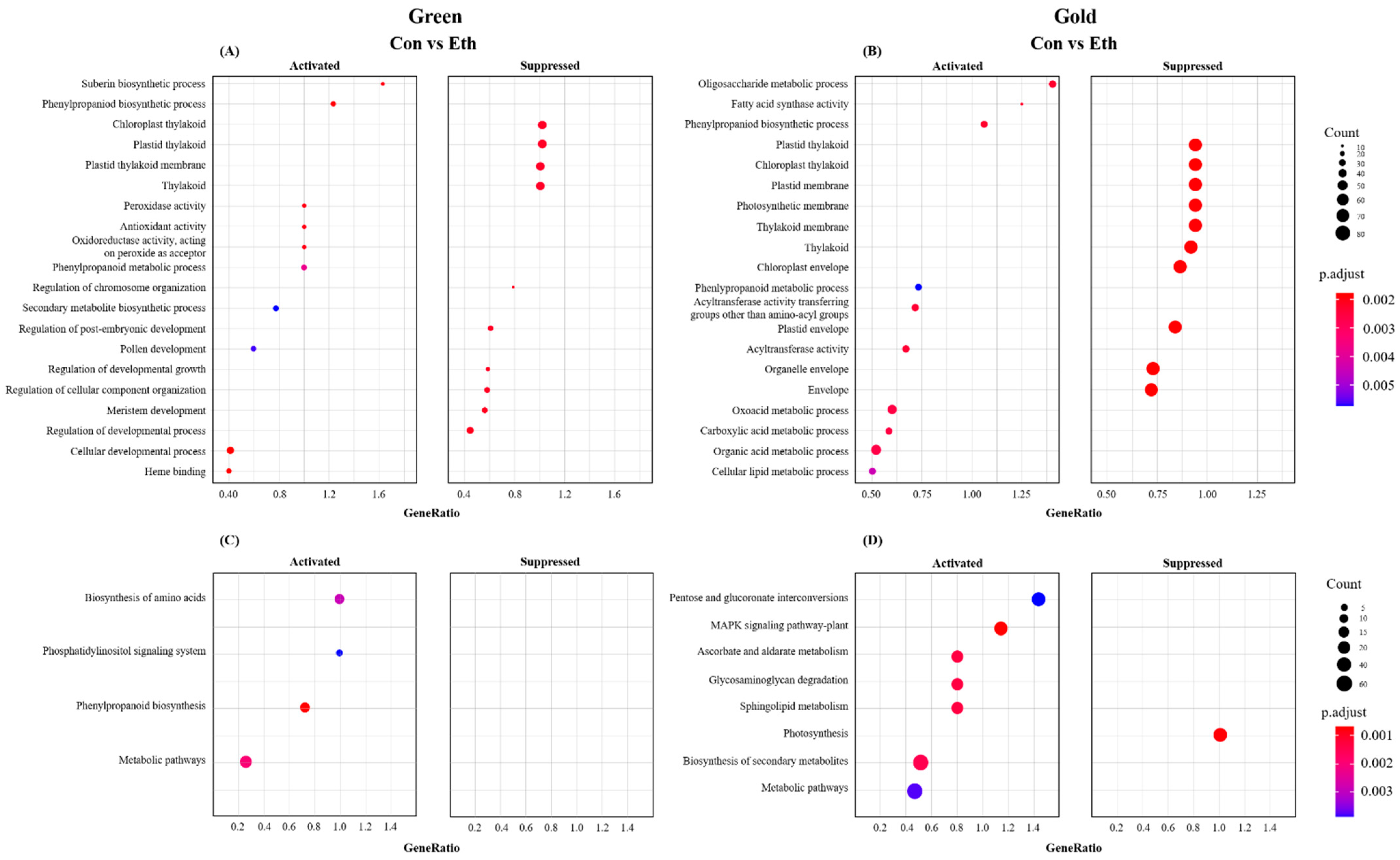
2.3. Identification of DEGs and Functional Enrichment Analysis
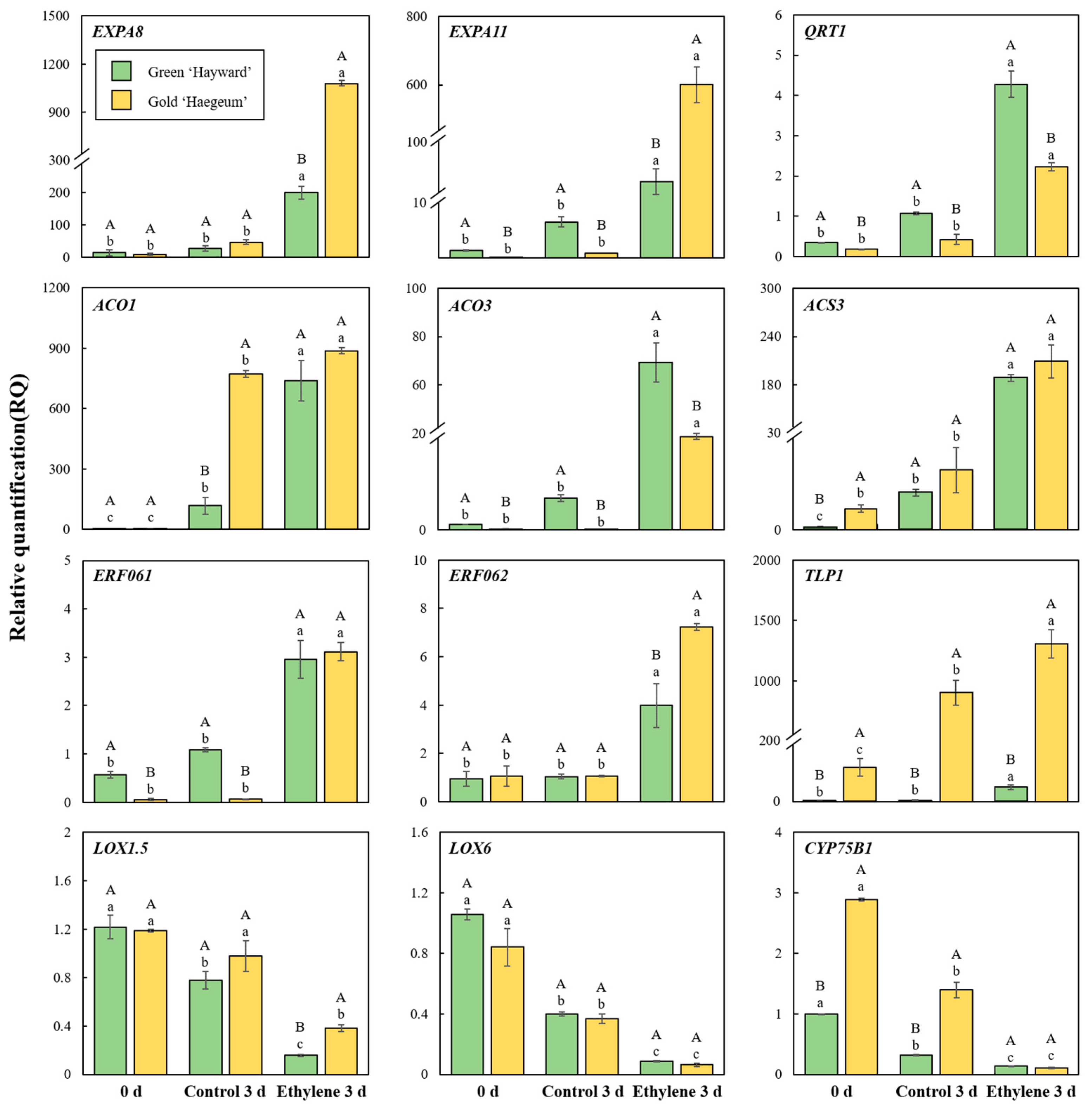
2.4. Verification of DEGs by qRT-PCR
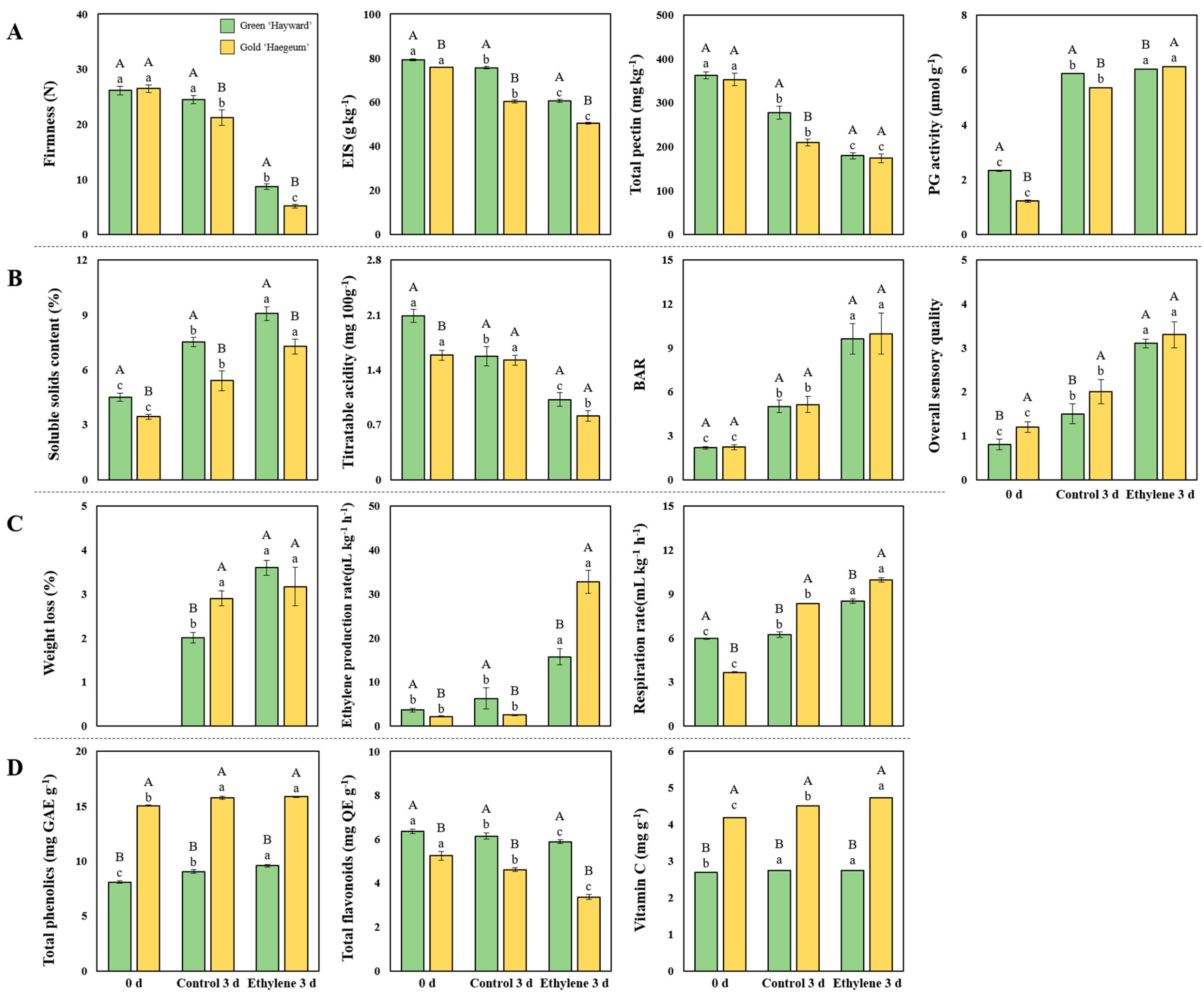
2.5. Measurement of Firmness, Pectin Content, EIS and Polygalacturonase(PG) Activity
2.6. Measurement of Soluble Solids Content (SSC), Titratable Acidity (TA), Brix-Acid Ratio (BAR) and Overall Acceptability
2.7. Weight Loss, Ethylene Production and Respiration Rates
2.8. Total Phenolics, Total Flavonoids, and Vitamin C
2.9. Statistical Analysis
3. Results and Discussion
3.1. Assembly and Annotation
3.2. Firmness, Total Pectin, EIS, PG Activity, and Related Genes
3.3. SSC, TA, BAR and Sensory Evaluation and the Related Genes
3.4. Ethylene Production and Respiration Rates and Related Genes
3.5. Stress-Related Genes Due to Ethylene Treatment
3.6. Verification of DEGs by qRT-PCR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, H.R.; Baek, M.W.; Cheol, L.H.; Jeong, C.S.; Tilahun, S. Changes in metabolites and antioxidant activities of green ‘Hayward’and gold ‘Haegeum’ kiwifruits during ripening with ethylene treatment. Food Chem. 2022, 384, 132490. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.R.; Baek, M.W.; Tilahun, S.; Jeong, C.S. Long-term cold storage affects metabolites, antioxidant activities, and ripening and stress-related genes of kiwifruit cultivars. Postharvest Biol. Technol. 2022, 189, 111912. [Google Scholar] [CrossRef]
- Tilahun, S.; Choi, H.R.; Lee, Y.M.; Choi, J.H.; Baek, M.W.; Hyok, K.; Park, S.M.; Jeong, C.S. Ripening quality of kiwifruit cultivars is affected by harvest time. Sci. Hortic. 2020, 261, 108936. [Google Scholar] [CrossRef]
- Lin, X.; Yang, R.; Dou, Y.; Zhang, W.; Du, H.; Zhu, L.; Chen, J. Transcriptome analysis reveals delaying of the ripening and cell-wall degradation of kiwifruit by hydrogen sulfide. J. Sci. Food Agric. 2020, 100, 2280–2287. [Google Scholar] [CrossRef]
- Hunter, D.C.; Skinner, M.A.; Ferguson, A.R. Kiwifruit and health. In Fruits, Vegetables, and Herbs; Academic Press: Cambridge, MA, USA, 2016; pp. 239–269. [Google Scholar] [CrossRef]
- Meena, N.K.; Baghel, M.; Jain, S.K.; Asrey, R. Postharvest biology and technology of kiwifruit. In Postharvest Biology and Technology of Temperate Fruits; Springer: Cham, Switzerland, 2018; pp. 299–329. [Google Scholar] [CrossRef]
- Chiaramonti, N.; Barboni, T. Relationship between the physicochemical parameters and the ethylene emission during cold storage of kiwifruits. Int. J. Food Sci. Technol. 2010, 45, 1513–1516. [Google Scholar] [CrossRef]
- Schroder, R.; Atkinson, R.G. Kiwifruit cell walls: Towards an understanding of softening? N. Z. J. For. Sci. 2006, 36, 112. [Google Scholar]
- Lim, S.; Lee, J.G.; Lee, E.J. Comparison of fruit quality and GC–MS-based metabolite profiling of kiwifruit ‘Jecy green’: Natural and exogenous ethylene-induced ripening. Food Chem. 2017, 234, 81–92. [Google Scholar] [CrossRef]
- Tilahun, S.; Park, K.C.; Choi, I.Y.; Jeong, C.S. Transcriptome analysis of astringent ‘Cheongdo-Bansi’persimmon fruit treated with ethylene for removal of astringency. Postharvest Biol. Technol. 2019, 150, 52–59. [Google Scholar] [CrossRef]
- Asif, M.H.; Lakhwani, D.; Pathak, S.; Gupta, P.; Bag, S.K.; Nath, P.; Trivedi, P.K. Transcriptome analysis of ripe and unripe fruit tissue of banana identifies major metabolic networks involved in fruit ripening process. BMC Plant Biol. 2014, 14, 316. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Xu, Q.; Da, X.; Guo, F.; Ding, Y.; Deng, X. Transcriptome changes during fruit development and ripening of sweet orange (Citrus sinensis). BMC Genom. 2012, 13, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Gao, P.; Liu, S.; Zhu, Z.; Amanullah, S.; Davis, A.R.; Luan, F. Comparative transcriptome analysis of two contrasting watermelon genotypes during fruit development and ripening. BMC Genom. 2017, 18, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.H.; Zhao, C.; Zhang, M.; Li, Y.Z.; Shen, Y.Y.; Guo, J.X. Transcriptome analysis around the onset of strawberry fruit ripening uncovers an important role of oxidative phosphorylation in ripening. Sci. Rep. 2017, 7, 41477. [Google Scholar] [CrossRef] [Green Version]
- Tilahun, S.; Choi, H.R.; Kwon, H.; Park, S.M.; Park, D.S.; Jeong, C.S. Transcriptome analysis of ‘Haegeum’ gold kiwifruit following ethylene treatment to improve postharvest ripening quality. Agronomy 2020, 10, 487. [Google Scholar] [CrossRef] [Green Version]
- Tilahun, S.; Heo, J.Y.; Jeong, C.S. Quality and expression of ethylene response genes of ‘Daebong’ persimmon fruit during ripening at different temperatures. Postharvest Biol. Technol. 2017, 133, 57–63. [Google Scholar] [CrossRef]
- Choi, H.R.; Tilahun, S.; Lee, Y.M.; Choi, J.H.; Baek, M.W.; Jeong, C.S. Harvest time affects quality and storability of kiwifruit (Actinidia spp.): Cultivars during long-term cool storage. Sci. Hortic. 2019, 256, 108523. [Google Scholar] [CrossRef]
- Baek, M.W.; Choi, H.R.; Solomon, T.; Jeong, C.S.; Lee, O.H.; Tilahun, S. Preharvest methyl jasmonate treatment increased the antioxidant activity and glucosinolate contents of hydroponically grown pak choi. Antioxidants 2021, 10, 131. [Google Scholar] [CrossRef]
- Krupa, T.; Latocha, P.; Liwińska, A. Changes of physicochemical quality, phenolics and vitamin C content in hardy kiwifruit (Actinidia arguta and its hybrid) during storage. Sci. Hortic. 2011, 130, 410–417. [Google Scholar] [CrossRef]
- Tilahun, S.; Choi, H.R.; Baek, M.W.; Cheol, L.H.; Kwak, K.W.; Park, D.S.; Solomon, T.; Jeong, C.S. Antioxidant Properties, γ-Aminobutyric Acid (GABA) Content, and Physicochemical Characteristics of Tomato Cultivars. Agronomy 2021, 11, 1204. [Google Scholar] [CrossRef]
- Fabi, J.P.; Broetto, S.G.; Silva, S.L.G.L.D.; Zhong, S.; Lajolo, F.M.; do Nascimento, J.R.O. Analysis of papaya cell wall-related genes during fruit ripening indicates a central role of polygalacturonases during pulp softening. PLoS ONE 2014, 9, e105685. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Li, T.; Jiang, G.; Jiang, Y.; Duan, X. Cell wall proteome analysis of banana fruit softening using iTRAQ technology. J. Proteom. 2019, 209, 103506. [Google Scholar] [CrossRef]
- Guo, D.L.; Xi, F.F.; Yu, Y.H.; Zhang, X.Y.; Zhang, G.H.; Zhong, G.Y. Comparative RNA-Seq profiling of berry development between table grape ‘Kyoho’and its early-ripening mutant ‘Fengzao’. BMC Genom. 2016, 17, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Cao, J. Differential gene expression of tropical pumpkin (Cucurbita moschata Duchesne) bush mutant during internode development. Sci. Hortic. 2008, 117, 219–224. [Google Scholar] [CrossRef]
- Jara, K.; Castro, R.I.; Ramos, P.; Parra-Palma, C.; Valenzuela-Riffo, F.; Morales-Quintana, L. Molecular insights into FaEG1, a strawberry endoglucanase enzyme expressed during strawberry fruit ripening. Plants 2019, 8, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sane, V.A.; Chourasia, A.; Nath, P. Softening in mango (Mangifera indica cv. Dashehari) is correlated with the expression of an early ethylene responsive, ripening related expansin gene, MiExpA1. Postharvest Biol. Technol. 2005, 38, 223–230. [Google Scholar] [CrossRef]
- Palapol, Y.; Kunyamee, S.; Thongkhum, M.; Ketsa, S.; Ferguson, I.B.; van Doorn, W.G. Expression of expansin genes in the pulp and the dehiscence zone of ripening durian (Durio zibethinus) fruit. J. Plant Physiol. 2015, 182, 33–39. [Google Scholar] [CrossRef]
- Kotake, T.; Aohara, T.; Hirano, K.; Sato, A.; Kaneko, Y.; Tsumuraya, Y.; Takatsuji, H.; Kawasaki, S. Rice Brittle culm 6 encodes a dominant-negative form of CesA protein that perturbs cellulose synthesis in secondary cell walls. J. Exp. Bot. 2011, 62, 2053–2062. [Google Scholar] [CrossRef] [Green Version]
- Malladi, A. Molecular physiology of fruit growth in apple. Hortic. Rev. 2020, 47, 1–42. [Google Scholar] [CrossRef]
- Cao, Y.; Tang, X.; Giovannoni, J.; Xiao, F.; Liu, Y. Functional characterization of a tomato COBRA-likegene functioning in fruit development and ripening. BMC Plant Biol. 2012, 12, 211. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Zhang, H.; Li, J.; Tao, S.; Qiao, X.; Korban, S.S.; Zhang, S.; Wu, J. Genome-wide analysis and characterization of molecular evolution of the HCT gene family in pear (Pyrus bretschneideri). Plant Syst. Evol. 2017, 303, 71–90. [Google Scholar] [CrossRef]
- Setiabudi, E.; Meitha, K.; Dwivany, F.M. In silico characterization and comparison of the fruit ripening related beta-amylase (BAM) gene family in banana genome A and B. Indones. J. Biotechnol. 2021, 26, 175–182. [Google Scholar] [CrossRef]
- Chen, J.; Duan, Y.; Hu, Y.; Li, W.; Sun, D.; Hu, H.; Xie, J. Transcriptome analysis of atemoya pericarp elucidates the role of polysaccharide metabolism in fruit ripening and cracking after harvest. BMC Plant Biol. 2019, 19, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Yuan, Y.; Zhang, C.; Li, H.; Ma, F.; Li, M. Sucrose phloem unloading follows an apoplastic pathway with high sucrose synthase in Actinidia fruit. Plant Sci. 2017, 255, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Moing, A.; Rothan, C.; Svanella, L.; Just, D.; Diakou, P.; Raymond, P.; Gaudillère, J.P.; Monet, R. Role of phosphoenolpyruvate carboxylase in organic acid accumulation during peach fruit development. Physiol. Plant. 2000, 108, 1–10. [Google Scholar] [CrossRef]
- Zhang, B.; Yin, X.R.; Li, X.; Yang, S.L.; Ferguson, I.B.; Chen, K.S. Lipoxygenase gene expression in ripening kiwifruit in relation to ethylene and aroma production. J. Agric. Food Chem. 2009, 57, 2875–2881. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Luca, V.D. The biosynthesis and regulation of biosynthesis of Concord grape fruit esters, including ‘foxy’ methylanthranilate. Plant J. 2005, 44, 606–619. [Google Scholar] [CrossRef]
- Zhang, R.G.; Andersson, C.E.; Savchenko, A.; Skarina, T.; Evdokimova, E.; Beasley, S.; Arrowsmith, C.H.; Edwards, A.M.; Joachimiak, A.; Mowbray, S.L. Structure of Escherichia coli ribose-5-phosphate isomerase: A ubiquitous enzyme of the pentose phosphate pathway and the Calvin cycle. Structure 2003, 11, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Pattyn, J.; Vaughan-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: A complex multilevel control circuitry. New Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef]
- Kende, H. Enzymes of ethylene biosynthesis. Plant Physiol. 1989, 91, 1–4. [Google Scholar] [CrossRef] [Green Version]
- John, P. How plant molecular biologists revealed a surprising relationship between two enzymes, which took an enzyme out of a membrane where it was not located, and put it into the soluble phase where it could be studied. Plant Mol. Biol. Report. 1991, 9, 192–194. [Google Scholar] [CrossRef]
- Choi, H.R.; Jeong, M.J.; Baek, M.W.; Choi, J.H.; Lee, H.C.; Jeong, C.S.; Tilahun, S. Transcriptome Analysis of Pre-Storage 1-MCP and High CO2-Treated ‘Madoka’ Peach Fruit Explains the Reduction in Chilling Injury and Improvement of Storage Period by Delaying Ripening. Int. J. Mol. Sci. 2021, 22, 4437. [Google Scholar] [CrossRef]
- Leitzmann, C. Characteristics and health benefits of phytochemicals. Complementary Med. Res. 2016, 23, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Balestrini, R.; Ghignone, S.; Quiroga, G.; Fiorilli, V.; Romano, I.; Gambino, G. Long-term impact of chemical and alternative fungicides applied to Grapevine cv Nebbiolo on Berry Transcriptome. Int. J. Mol. Sci. 2020, 21, 6067. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, S.; Sato, F. Plant pathogenesis-related proteins: Molecular mechanisms of gene expression and protein function. J. Biochem. 1999, 125, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Selitrennikoff, C.P. Antifungal proteins. Appl. Environ. Microbiol. 2001, 67, 2883–2894. [Google Scholar] [CrossRef] [Green Version]
- Wurms, K.; Cui, W.; Ah-Chee, A.; Rees-George, J.; Bublin, M.; Breiteneder, H. Down Regulation of Putative Defence-associated Transcripts Correlates with Ripe Rot Symptoms on Kiwifruit (Actinidia chinensis). J. Phytopathol. 2011, 159, 435–442. [Google Scholar] [CrossRef]
- Bai, S.; Dong, C.; Li, B.; Dai, H. A PR-4 gene identified from Malus domestica is involved in the defense responses against Botryosphaeria dothidea. Plant Physiol. Biochem. 2013, 62, 23–32. [Google Scholar] [CrossRef]
- Cao, J.; Lv, Y.; Hou, Z.; Li, X.; Ding, L. Expansion and evolution of thaumatin-like protein (TLP) gene family in six plants. Plant Growth Regul. 2016, 79, 299–307. [Google Scholar] [CrossRef]
- Li, H.; Han, X.; Qiu, W.; Xu, D.; Wang, Y.; Yu, M.; Wang, Y.; Zhuo, R. Identification and expression analysis of the GDSL esterase/lipase family genes, and the characterization of SaGLIP8 in Sedum alfredii Hance under cadmium stress. PeerJ 2019, 7, e6741. [Google Scholar] [CrossRef] [Green Version]
- Ni, P.Y.; Ji, X.R.; Guo, D.L. Genome-wide identification, characterization, and expression analysis of GDSL-type esterases/lipases gene family in relation to grape berry ripening. Sci. Hortic. 2020, 264, 109162. [Google Scholar] [CrossRef]
- Singh, A.; Singh, S.; Singh, R.; Kumar, S.; Singh, S.K.; Singh, I.K. Dynamics of Zea mays transcriptome in response to a polyphagous herbivore, Spodoptera litura. Funct. Integr. Genom. 2021, 21, 571–592. [Google Scholar] [CrossRef]
- Giordano, D.; Provenzano, S.; Ferrandino, A.; Vitali, M.; Pagliarani, C.; Roman, F.; Cardinale, F.; Castellarin, S.D.; Schubert, A. Characterization of a multifunctional caffeoyl-CoA O-methyltransferase activated in grape berries upon drought stress. Plant Physiol. Biochem. 2016, 101, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.; Moon, S.J.; Han, S.; Kim, B.G.; Park, S.R.; Lee, S.K.; Yoon, H.J.; Lee, H.E.; Kwon, H.B.; Baek, D.W.; et al. Expression of StMYB1R-1, a novel potato single MYB-like domain transcription factor, increases drought tolerance. Plant Physiol. 2011, 155, 421–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2012, 1819, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Ni, L.; Carballar-Lejarazú, R.; Zou, X.; Sun, W.; Wu, L.; Yuan, X.; Mao, Y.; Huang, W.; Zou, S. Comparative transcriptome among Euscaphis konishii Hayata tissues and analysis of genes involved in flavonoid biosynthesis and accumulation. BMC Genom. 2019, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Ishimaru, M.; Ding, C.K.; Yakushiji, H.; Goto, N. Comparison of UDP-glucose: Flavonoid 3-O-glucosyltransferase (UFGT) gene sequences between white grapes (Vitis vinifera) and their sports with red skin. Plant Sci. 2001, 160, 543–550. [Google Scholar] [CrossRef]
- Oh, T.G.; Jo, J.A.; Lee, S.J. Evaluation of time–temperature integrator for indicating the ripeness of kiwifruit in plastic container at home. J. Food Sci. 2021, 86, 2872–2885. [Google Scholar] [CrossRef]
- Yihui, G.; Song, J.; Du, L.; Vinqvist, M.; Palmer, L.C.; Fillmore, S.; Pang, X.; Zhang, Z. Characterization of laccase from apple fruit during postharvest storage and its response to diphenylamine and 1-methylcyclopropene treatments. Food Chem. 2018, 253, 314–321. [Google Scholar] [CrossRef]
- Wu, F.; Li, Q.; Yan, H.; Zhang, D.; Jiang, G.; Jiang, Y.; Duan, X. Characteristics of three thioredoxin genes and their role in chilling tolerance of harvested banana fruit. Int. J. Mol. Sci. 2016, 17, 1526. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | |||||
| Sample | Raw Data | Trimmed Data | |||
| Total reads | Total read bases (bp) | Total reads | Total read bases (bp) | % | |
| Green_Con | 85,610,346 | 13,520,193,606 | 85,610,346 | 12,683,777,637 | 93.8% |
| Green_Eth | 75,655,556 | 11,974,790,146 | 75,655,556 | 11,152,550,066 | 93.1% |
| Gold_Con | 61,430,214 | 6,204,451,614 | 58,940,256 | 5,834,133,639 | 94.0% |
| Gold_Eth | 61,777,836 | 6,239,561,436 | 59,494,040 | 5,873,712,144 | 94.1% |
| (B) | |||||
| Sample | Progressed read | Mapped reads | Properly paired reads | Mapping rate | |
| Green_Con | 85,610,346 | 72,135,718 | 65,600,622 | 84.3% | |
| Green_Eth | 75,665,556 | 64,094,382 | 59,061,836 | 84.7% | |
| Gold_Con | 58,940,256 | 51,437,355 | 48,447,084 | 87.3% | |
| Gold_Eth | 59,494,040 | 52,721,368 | 49,888,316 | 88.6% | |
| (C) | |||||
| Total transcripts | p < 0.05, ǀlog2 fold changeǀ ≥ 1 | ||||
| UP | DOWN | TOTAL | |||
| Green Con vs. Eth | 26,130 | 594 | 693 | 1287 | |
| Gold Con vs. Eth | 28,605 | 906 | 818 | 1724 | |
| Gene_id | Gene Descriptions | Log2 Fold Change | p Value | ||
|---|---|---|---|---|---|
| Green | Gold | Green | Gold | ||
| Upregulated | |||||
| Actinidia04880.t1 | expansin-A1 | 8.41 | 6.65 | 0.000 | 0.024 |
| Actinidia15395.t1 | aldehyde oxidase GLOX-like | 7.59 | 7.52 | 0.003 | 0.016 |
| Actinidia22915.t1 | chitinase 1-like | 7.50 | 5.58 | 0.000 | 0.000 |
| Actinidia31692.t1 | pathogenesis-related protein 4 | 7.44 | 8.77 | 0.005 | 0.000 |
| Actinidia02029.t1 | thaumatin-like protein | 7.24 | 4.74 | 0.000 | 0.002 |
| Actinidia22914.t1 | basic endochitinase | 6.30 | 5.83 | 0.000 | 0.002 |
| Actinidia01237.t1 | methanol O-anthraniloyltransferase-like | 5.23 | 6.43 | 0.000 | 0.002 |
| Actinidia35948.t1 | probable pectate lyase 8 | 5.02 | 6.13 | 0.000 | 0.002 |
| Actinidia29762.t1 | expansin-A11 | 4.33 | 6.18 | 0.000 | 0.002 |
| Actinidia09749.t1 | 1-aminocyclopropane-1-carboxylate oxidase 3 | 4.31 | 3.93 | 0.000 | 0.011 |
| Actinidia33383.t1 | laccase-15-like | 3.98 | 5.91 | 0.039 | 0.020 |
| Actinidia35390.t1 | pectinesterase QRT1-like | 3.97 | 6.68 | 0.000 | 0.000 |
| Actinidia22149.t1 | ethylene-responsive transcription factor ERF061-like | 3.95 | 6.41 | 0.000 | 0.000 |
| Actinidia16205.t1 | probable pectate lyase 18 | 3.89 | 4.61 | 0.008 | 0.022 |
| Actinidia10870.t1 | beta-amylase 3, chloroplastic | 3.89 | 6.42 | 0.000 | 0.003 |
| Actinidia33182.t1 | ethylene-responsive transcription factor ERF062 | 3.83 | 5.51 | 0.014 | 0.005 |
| Actinidia00998.t1 | expansin-A8-like | 3.78 | 5.50 | 0.000 | 0.002 |
| Actinidia04756.t1 | cellulose synthase A catalytic subunit 2 | 3.71 | 2.80 | 0.000 | 0.034 |
| Actinidia28962.t1 | polygalacturonase-like | 2.84 | 6.10 | 0.027 | 0.006 |
| Actinidia25260.t1 | 1-aminocyclopropane-1-carboxylate oxidase 1 | 2.84 | 4.52 | 0.029 | 0.004 |
| Actinidia11956.t1 | 1-aminocyclopropane-1-carboxylate synthase 3 | 2.65 | 6.17 | 0.009 | 0.000 |
| Actinidia23856.t1 | GDSL esterase/lipase | 1.93 | 8.06 | 0.045 | 0.000 |
| Downregulated | |||||
| Actinidia04831.t1 | flavonoid 3′-monooxygenase-like isoform X2 | −2.66 | −3.54 | 0.035 | 0.013 |
| Actinidia25766.t1 | lipoxygenase 6, chloroplastic | −2.69 | −3.63 | 0.016 | 0.012 |
| Actinidia18864.t1 | putative UDP-glucose flavonoid 3-O-glucosyltransferase 3 | −2.46 | −3.90 | 0.034 | 0.004 |
| Actinidia05691.t1 | probable linoleate 9S-lipoxygenase 5 isoform X2 | −3.29 | −4.35 | 0.001 | 0.002 |
| Actinidia18971.t1 | LOW QUALITY PROTEIN: flavonol sulfotransferase-like | −4.29 | −5.46 | 0.003 | 0.000 |
| Actinidia20469.t1 | endoglucanase 24 | −4.05 | −6.74 | 0.022 | 0.022 |
| Actinidia28570.t1 | probable WRKY transcription factor 70 | −6.74 | −7.44 | 0.023 | 0.031 |
| Actinidia12711.t1 | transcription factor MYB41 | −2.42 | −11.42 | 0.024 | 0.000 |
| Gene_id | Gene Descriptions | Log2 Fold Change | p Value |
|---|---|---|---|
| Upregulated | |||
| Actinidia04140.t1 | xyloglucan endotransglucosylase/hydrolase protein 22-like | 8.68 | 0.000 |
| Actinidia21371.t1 | probable pectinesterase 53 | 8.16 | 0.001 |
| Actinidia24466.t1 | flavonoid 3′,5′-hydroxylase 2 | 7.40 | 0.006 |
| Actinidia17388.t1 | endochitinase-like | 6.97 | 0.011 |
| Actinidia33393.t1 | endochitinase EP3 | 6.96 | 0.000 |
| Actinidia17733.t1 | pectin acetylesterase 12 | 6.77 | 0.021 |
| Actinidia05128.t2 | thioredoxin-like protein CXXS1 | 6.64 | 0.009 |
| Actinidia15218.t1 | 1-aminocyclopropane-1-carboxylate oxidase 5 | 6.52 | 0.000 |
| Actinidia39196.t1 | glucan endo-1,3-beta-glucosidase, acidic-like | 6.22 | 0.038 |
| Actinidia38435.t2 | probable S-adenosylmethionine-dependent methyltransferase At5g38100 | 6.03 | 0.032 |
| Actinidia22508.t1 | beta-galactosidase BG1-like precursor | 4.69 | 0.034 |
| Actinidia38371.t1 | alpha-xylosidase 1-like | 4.64 | 0.009 |
| Actinidia05812.t1 | probable xyloglucan endotransglucosylase/hydrolase protein 30 | 4.22 | 0.000 |
| Actinidia08156.t1 | polygalacturonase At1g48100-like isoform X1 | 3.65 | 0.030 |
| Actinidia25891.t1 | ethylene-responsive transcription factor 2-like | 3.42 | 0.006 |
| Actinidia18363.t1 | pathogenesis-related protein PR-4-like | 3.18 | 0.002 |
| Actinidia40364.t1 | glucan endo-1,3-beta-glucosidase, basic vacuolar isoform-like | 2.73 | 0.028 |
| Downregulated | |||
| Actinidia27081.t1 | electron transfer flavoprotein-ubiquinone oxidoreductase, mitochondrial isoform X1 | −2.03 | 0.036 |
| Actinidia17597.t1 | protein COBRA-like | −2.20 | 0.023 |
| Actinidia19061.t1 | flavonol synthase/flavanone 3-hydroxylase-like | −3.78 | 0.016 |
| Actinidia15644.t1 | sucrose synthase-like | −5.89 | 0.034 |
| Actinidia25276.t1 | probable caffeoyl-CoA O-methyltransferase At4g26220 isoform X3 | −5.95 | 0.015 |
| Actinidia39608.t2 | probable pectinesterase/pectinesterase inhibitor 12 | −6.39 | 0.000 |
| Gene_id | Gene Descriptions | Log2 Fold Change | p Value |
|---|---|---|---|
| Upregulated | |||
| Actinidia27791.t1 | polygalacturonase At1g48100 | 7.99 | 0.019 |
| Actinidia12748.t1 | thioredoxin-like protein CITRX1, chloroplastic | 7.29 | 0.041 |
| Actinidia26056.t1 | beta-galactosidase 17-like | 7.08 | 0.021 |
| Actinidia39719.t1 | beta-galactosidase-like isoform X1 | 5.64 | 0.000 |
| Actinidia39827.t1 | ethylene-responsive transcription factor ERF118 | 5.05 | 0.010 |
| Actinidia18256.t1 | shikimate O-hydroxycinnamoyltransferase-like | 4.81 | 0.001 |
| Actinidia37578.t1 | probable pectate lyase 8 isoform X1 | 4.77 | 0.007 |
| Actinidia23167.t1 | expansin-A4-like | 4.68 | 0.004 |
| Actinidia20414.t1 | ethylene-responsive transcription factor TINY-like | 4.63 | 0.032 |
| Actinidia28999.t1 | probable glucan endo-1,3-beta-glucosidase A6 | 4.56 | 0.019 |
| Actinidia28348.t1 | 1-aminocyclopropane-1-carboxylate synthase-like | 4.55 | 0.016 |
| Actinidia29796.t1 | 1-aminocyclopropane-1-carboxylate oxidase 1 isoform X1 | 4.44 | 0.005 |
| Actinidia32144.t1 | beta-galactosidase-like | 4.39 | 0.002 |
| Actinidia21025.t1 | ethylene-responsive transcription factor ERF073 | 3.79 | 0.006 |
| Actinidia07250.t1 | glucan endo-1,3-beta-glucosidase 14 | 3.78 | 0.010 |
| Actinidia35006.t1 | thioredoxin-like 1–2, chloroplastic | 3.47 | 0.010 |
| Actinidia36687.t1 | probable ribose-5-phosphate isomerase 2 | 3.27 | 0.019 |
| Actinidia26661.t1 | S-adenosylmethionine synthase 1 | 3.05 | 0.022 |
| Actinidia18544.t1 | probable pectate lyase 18 isoform X1 | 2.91 | 0.041 |
| Actinidia28909.t1 | S-adenosylmethionine synthase 3 | 2.82 | 0.035 |
| Actinidia14729.t2 | pectin acetylesterase 8-like | 2.73 | 0.042 |
| Downregulated | |||
| Actinidia33920.t1 | leucoanthocyanidin dioxygenase-like | −3.07 | 0.044 |
| Actinidia20664.t1 | cellulose synthase A catalytic subunit 3 | −4.71 | 0.001 |
| Actinidia10640.t1 | cellulose synthase A catalytic subunit 2 | −5.30 | 0.017 |
| Actinidia14835.t1 | phosphoenolpyruvate carboxylase 4 isoform X2 | −5.75 | 0.000 |
| Actinidia34216.t1 | probable pectinesterase/pectinesterase inhibitor 34 | −6.32 | 0.000 |
| Actinidia12573.t1 | phosphoenolpyruvate carboxylase, housekeeping isozyme | −6.32 | 0.001 |
| Actinidia19849.t1 | glutathione S-transferase U17 | −6.79 | 0.028 |
| Actinidia03363.t1 | cinnamoyl-CoA reductase-like SNL6 | −7.03 | 0.031 |
| Actinidia11714.t1 | endoglucanase 25-like | −7.75 | 0.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.R.; Baek, M.W.; Jeong, C.S.; Tilahun, S. Comparative Transcriptome Analysis of Softening and Ripening-Related Genes in Kiwifruit Cultivars Treated with Ethylene. Curr. Issues Mol. Biol. 2022, 44, 2593-2613. https://doi.org/10.3390/cimb44060177
Choi HR, Baek MW, Jeong CS, Tilahun S. Comparative Transcriptome Analysis of Softening and Ripening-Related Genes in Kiwifruit Cultivars Treated with Ethylene. Current Issues in Molecular Biology. 2022; 44(6):2593-2613. https://doi.org/10.3390/cimb44060177
Chicago/Turabian StyleChoi, Han Ryul, Min Woo Baek, Cheon Soon Jeong, and Shimeles Tilahun. 2022. "Comparative Transcriptome Analysis of Softening and Ripening-Related Genes in Kiwifruit Cultivars Treated with Ethylene" Current Issues in Molecular Biology 44, no. 6: 2593-2613. https://doi.org/10.3390/cimb44060177
APA StyleChoi, H. R., Baek, M. W., Jeong, C. S., & Tilahun, S. (2022). Comparative Transcriptome Analysis of Softening and Ripening-Related Genes in Kiwifruit Cultivars Treated with Ethylene. Current Issues in Molecular Biology, 44(6), 2593-2613. https://doi.org/10.3390/cimb44060177