
Tryptase-Positive Mast Cells Promote Adipose Fibrosis in Secondary Lymphedema through PDGF


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. General Histological and Immunohistological Studies
2.3. RT-PCR
2.4. Statistical Analysis
3. Results
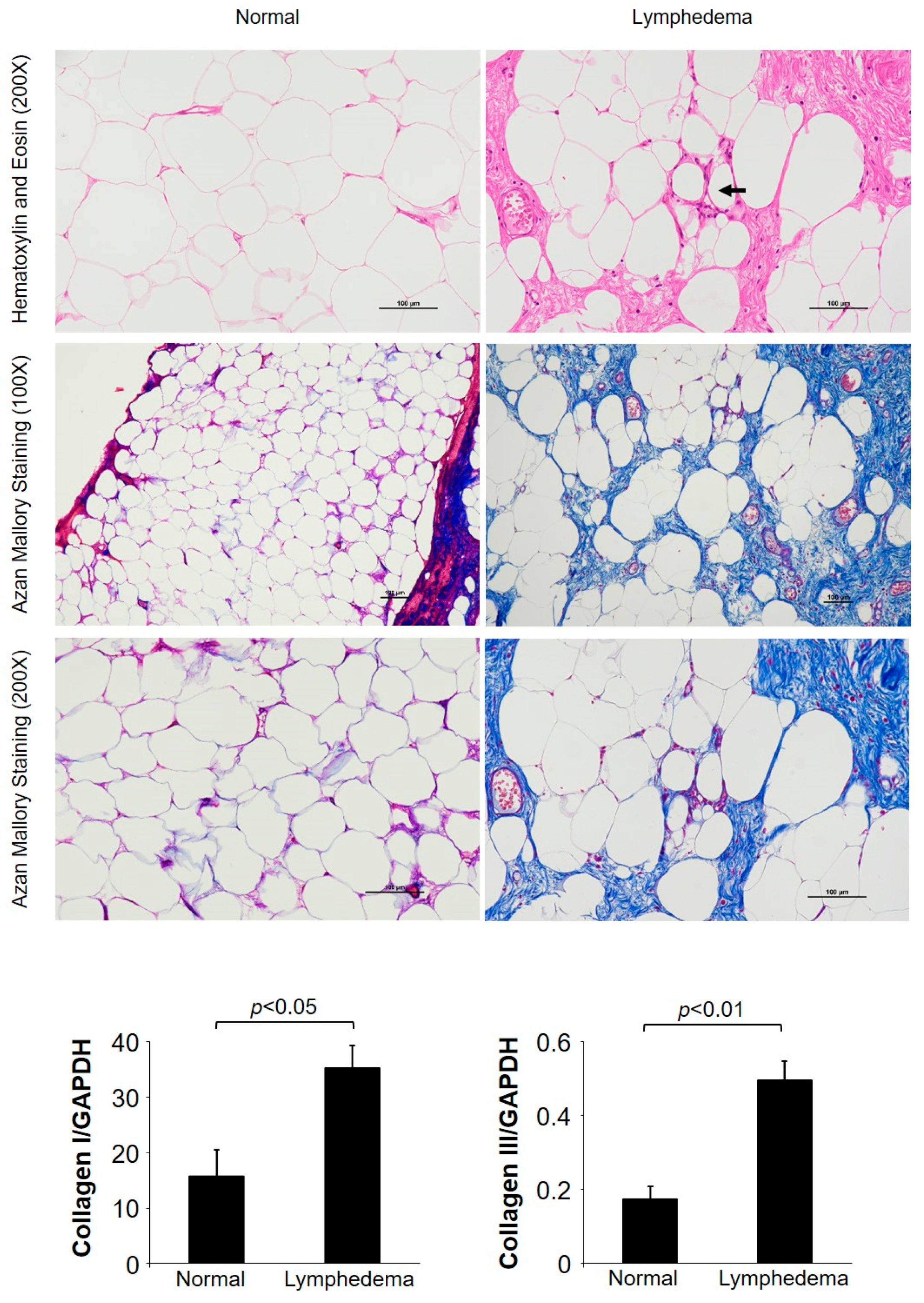
3.1. General Histological Examinations
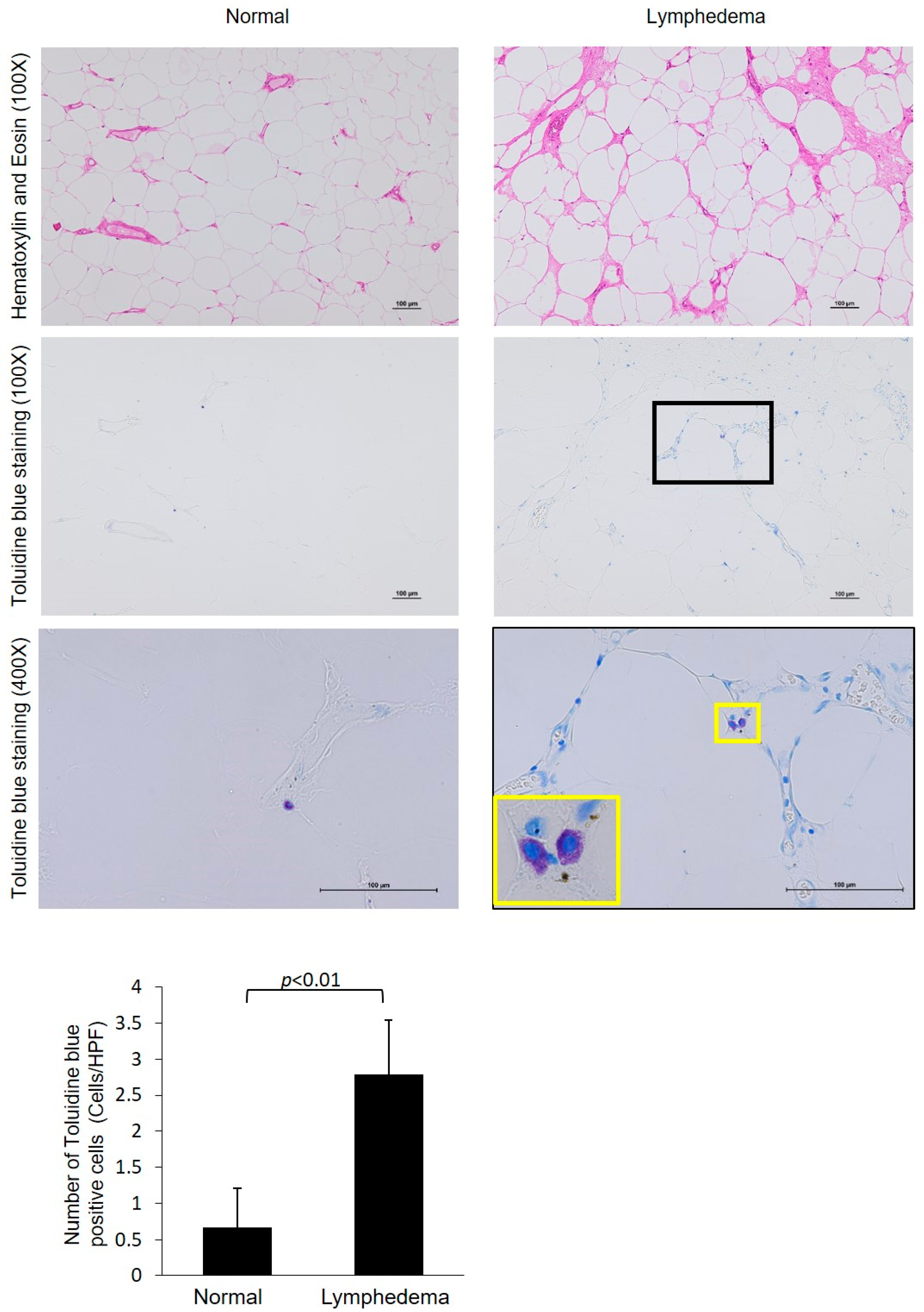
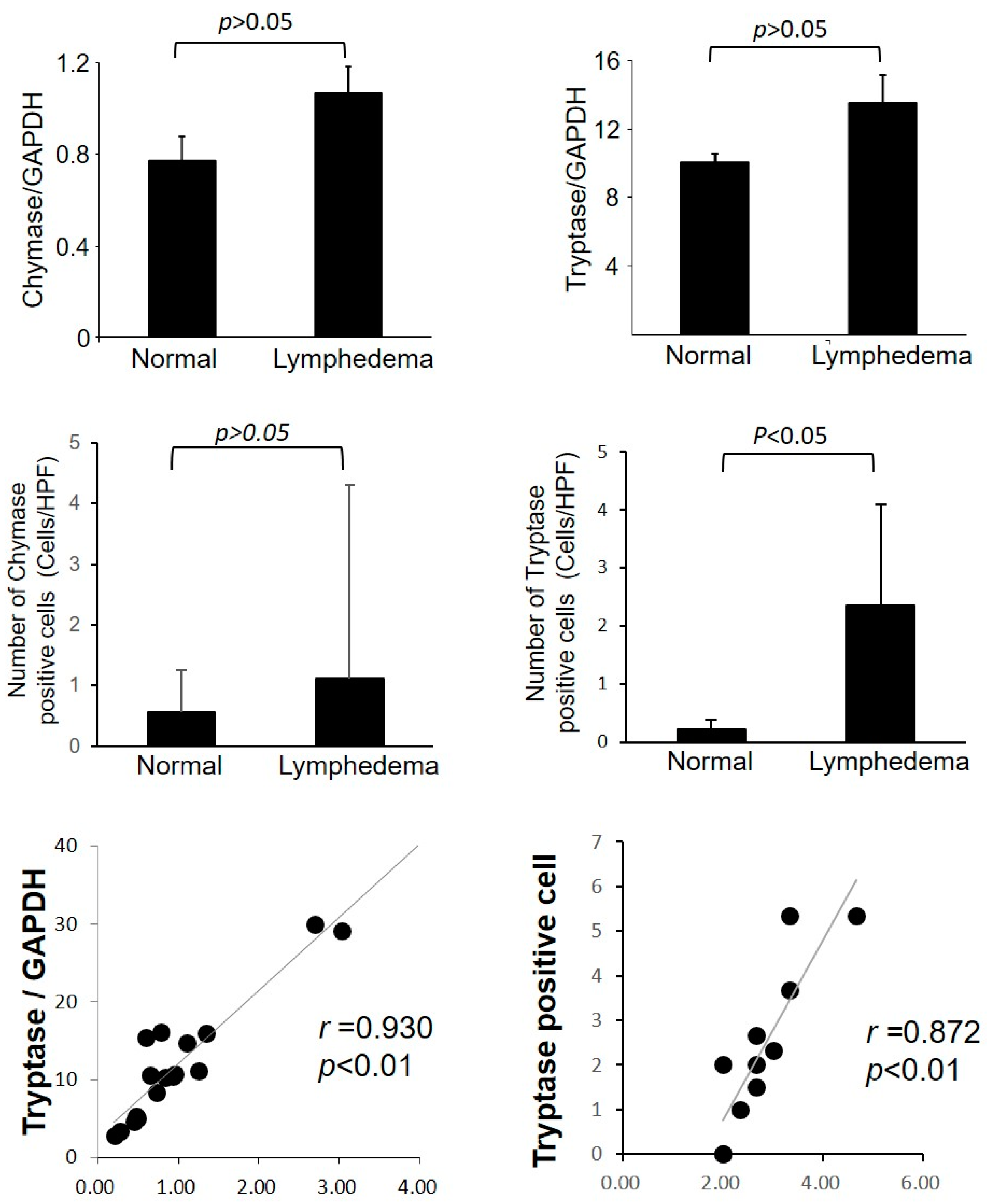
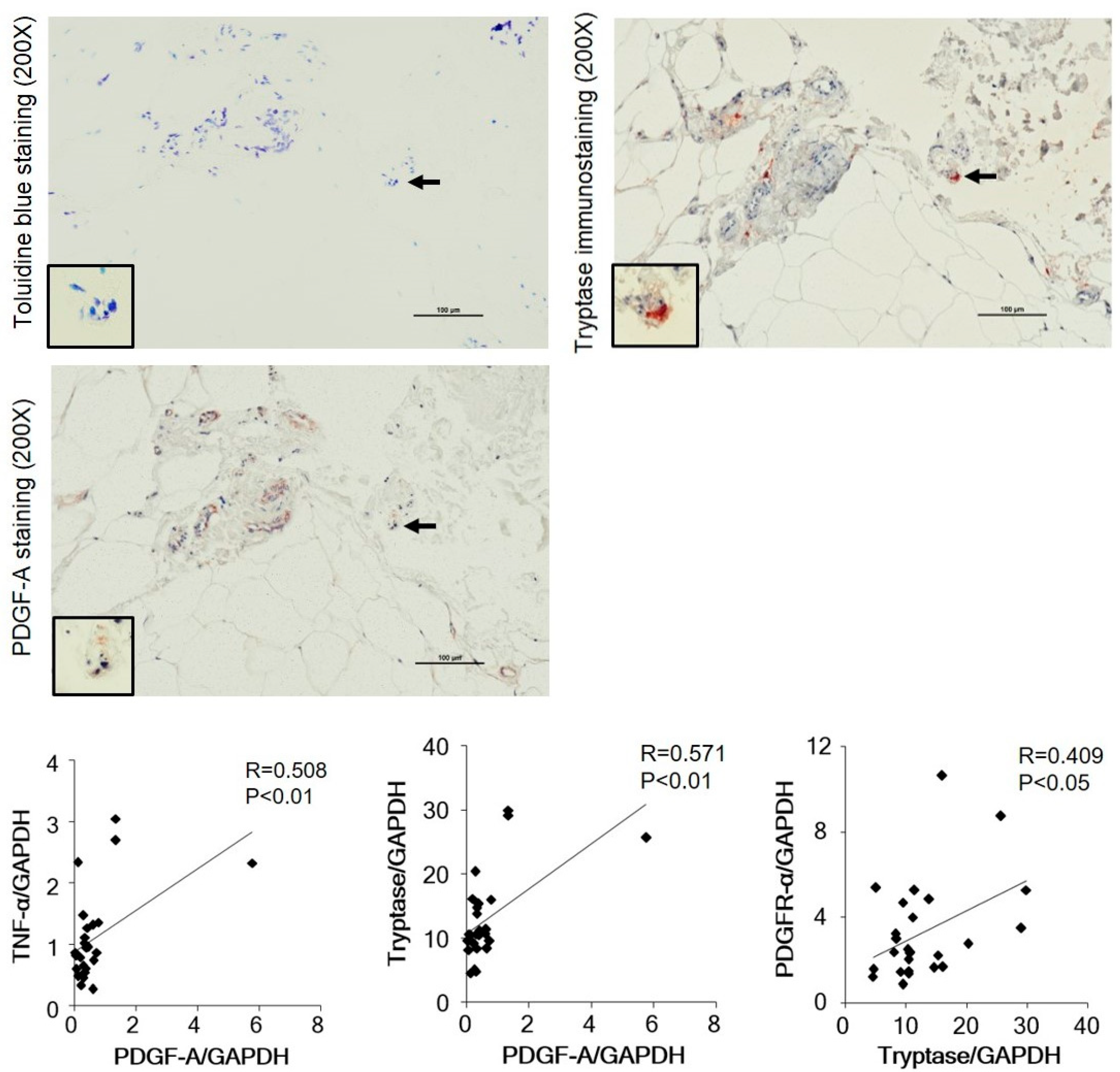
3.2. Identification of Mast Cell Type by Immunohistological Examinations
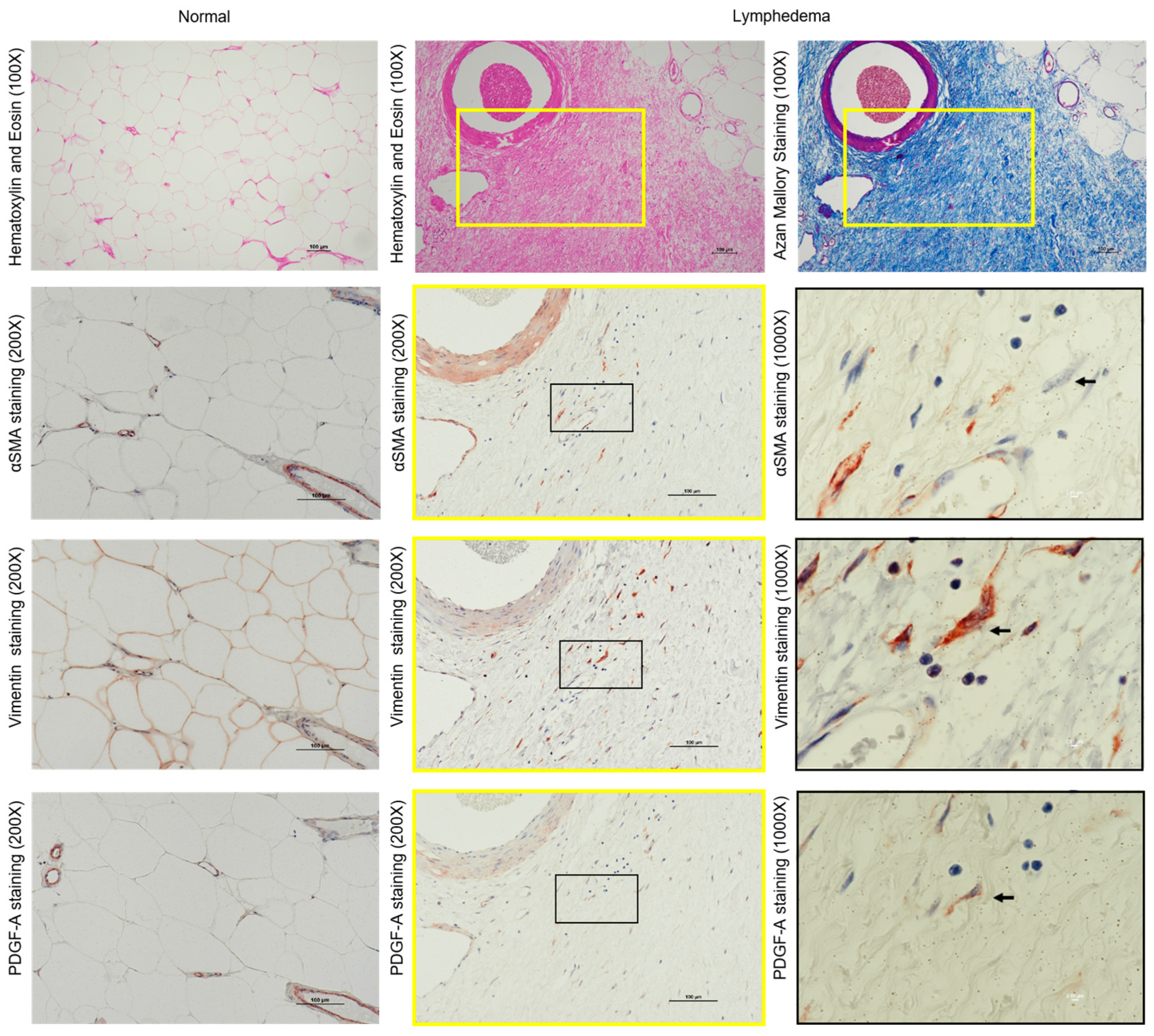
3.3. Identification of Fibroblasts and Myofibroblasts in the Fibrotic Regions of Lymphedema
4. Discussion
Limitation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hong, J.H.; Tsai, C.S.; Lai, C.H.; Chang, T.C.; Wang, C.C.; Lee, S.P.; Tseng, C.J.; Hsueh, S. Postoperative low-pelvic irradiation for stage I-IIA cervical cancer patients with risk factors other than pelvic lymph node metastasis. Int. J. Radiat. Oncol. Biol. Phys. 2002, 53, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.; Stainton, M.C.; Slaytor, E.K.; Jaconelli, C.; Watts, A.; Mackenzie, P. Aetiology and prevalence of lower limb lymphoedema following treatment for gynaecological cancer. Aust. N. Z. J. Obstet. Gynaecol. 2003, 43, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Petrek, J.A.; Heelan, M.C. Incidence of breast carcinoma-related lymphedema. Cancer 1998, 83, 2776–2781. [Google Scholar] [CrossRef]
- International Society of Lymphology Executive Committee. The Diagnosis and Treatment of Peripheral Lymphedema. Lymphology 1995, 28, 113–117. [Google Scholar]
- Mihara, M.; Hara, H.; Hayashi, Y.; Narushima, M.; Yamamoto, T.; Todokoro, T.; Iida, T.; Sawamoto, N.; Araki, J.; Kikuchi, K.; et al. Pathological steps of cancer-related lymphedema: Histological changes in the collecting lymphatic vessels after lymphadenectomy. PLoS ONE 2012, 7, e41126. [Google Scholar] [CrossRef]
- Ogata, F.; Fujiu, K.; Koshima, I.; Nagai, R.; Manabe, I. Phenotypic modulation of smooth muscle cells in lymphedema. Br. J. Dermatol. 2015, 172, 1286–1293. [Google Scholar] [CrossRef]
- DeBari, M.K.; Abbott, R.D. Adipose tissue fibrosis: Mechanism, Models, and Importance. Int. J. Mol. Sci. 2020, 21, 6030. [Google Scholar] [CrossRef]
- Mehrara, B.J.; Park, H.J.; Kataru, R.P.; Bromberg, J.; Coriddi, M.; Baik, J.E.; Shin, J.; Li, C.; Cavalli, M.R.; Encarnacion, E.M.; et al. Pilot study of anti-Th2 immunotherapy for the treatment of breast cancer-related upper extremity lymphedema. Biology 2021, 10, 934. [Google Scholar] [CrossRef]
- Avraham, T.; Zampell, J.C.; Yan, A.; Elhadad, S.; Weitman, E.S.; Rockson, S.G.; Bromberg, J.; Mehhara, B. Th2 differentiation is necessary for soft tissue fibrosis and lymphatic dysfunction resulting from lymphedema. FASEB J. 2013, 27, 1114–1126. [Google Scholar] [CrossRef]
- Nuri, T.; Iwanaga, H.; Otsuki, Y.; Ueda, K. Lymphoscintigraphy for prediction of effect of lymphaticovenular anastomosis for treatment of secondary lower limb lymphedema. J. Vasc. Surg. Venous. Lymphat. Disord. 2022, 10, 1079–1086. [Google Scholar] [CrossRef]
- Sire, A.D.; Losco, L.; Lippi, L.; Spadoni, D.; Kaciulyte, J.; Sert, G.; Ciamarra, P.; Marcasciano, M.; Cuomo, R.; Bolletta, A.; et al. Surgical Treatment and Rehabilitation Strategies for Upper and Lower Extremity Lymphedema: A Comprehensive Review. Medicina 2022, 58, 954. [Google Scholar] [CrossRef] [PubMed]
- Armbrust, T.; Batusic, D.; Ringe, B.; Ramadori, G. Mast cells distribution in human liver disease and experimental rat liver fibrosis. Indications for mast cell participation in development of liver fibrosis. J. Hepatol. 1997, 26, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.T.; Cruz, A.C.; Donnelly, S.; Dazin, P.F.; Schulman, E.S.; Jones, K.D.; Wolters, P.J.; Hoopes, C.; Dolganov, G.M.; Fang, K.C. c-Kit immunophenotyping and metalloproteinase expression profiles of mast cells in interstitial lung diseases. J. Pathol. 2005, 206, 279–290. [Google Scholar] [CrossRef]
- Zimnoch, L.; Szynaka, B.; Puchalski, Z. Mast cells and pancreatic stellate cells in chronic pancreatitis with differently intensified fibrosis. Hepatogastroenterology 2002, 49, 1135–1138. [Google Scholar] [PubMed]
- Di, S.; Ziyou, Y.; Liu, N.F. Pathological Changes of Lymphedematous Skin: Increased Mast Cells, Related Proteases, and Activated Transforming Growth Factor-β1. Lymphat. Res. Biol. 2016, 14, 162–171. [Google Scholar] [CrossRef]
- Altintas, M.M.; Azad, A.; Nayer, B.; Contreras, G.; Zaias, J.; Faul, C.; Reiser, J.; Nayer, A. Mast cells, macrophages, and crown-like structures distinguish subcutaneous from visceral fat in mice. J. Lipid. Res. 2011, 52, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Divoux, A.; Sun, J.; Zhang, J.; Clément, K.; Glickman, J.N.; Sukhova, G.K.; Wolters, P.J.; Jean, D.; Gorgun, C.Z.; et al. Genetic deficiency and pharmacological stabilization of mast cells reduce diet-induced obesity and diabetes in mice. Nat. Med. 2009, 15, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Ueda, H.; Takai, S.; Okamoto, Y.; Muramatsu, M.; Sakaguchi, M.; Shibahara, N.; Katsuoka, Y.; Miyazaki, M. Effect of chymase inhibition on the arteriovenous fistula stenosis in dogs. J. Am. Soc. Nephrol. 2005, 16, 1024–1034. [Google Scholar] [CrossRef]
- Irani, A.M.; Bradford, T.R.; Kepley, C.L.; Schechter, N.M.; Schwartz, L.B. Detection of MCT and MCTC types of human mast cells by immunohistochemistry using new monoclonal anti-tryptase and anti-chymase antibodies. Histochem. Cytochem. 1989, 37, 1509–1515. [Google Scholar] [CrossRef]
- Warren, A.G.; Brorson, H.; Borud, L.; Slavi, S.A. Lymphedem: A comprehensive review. Ann. Plast. Surg. 2007, 59, 464–472. [Google Scholar] [CrossRef]
- Reed, R.K.; Laurent, T.C.; Taylor, A.E. Hyaluronan in prenodal lymph from skin: Changes with lymph flow. Am. J. Physiol. Circ. Physiol. 1990, 259, H1097–H1100. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, W.L. The Pathophysiology of Lymphedema-2012. Handchir. Mikrochir. Plast. Chir. 2012, 44, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Rockson, S.G. The Lymphatics and the Inflammatory Response: Lessons Learned from Human Lymphedema. Lymphat. Res. Biol. 2013, 11, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Gousopoulos, E.; Proulx, S.T.; Bachmann, S.B.; Scholl, J.; Dionyssiou, D.; Demiri, E.; Halin, C.; Dieterich, L.C.; Detmar, M. Regulatory T cell transfer ameliorates lymphedema and promotes lymphatic vessel function. JCI Insight 2016, 1, e89081. [Google Scholar] [CrossRef]
- Lin, S.; Kim, J.; Lee, M.-J.; Roche, L.; Yang, N.L.; Tsao, P.S.; Rockson, S.G. Prospective Transcriptomic Pathway Analysis of Human Lymphatic Vascular Insufficiency: Identification and Validation of a Circulating Biomarker Panel. PLoS ONE 2012, 7, e52021. [Google Scholar] [CrossRef]
- Mihara, M.; Hara, H.; Kawakami, Y.; Zhou, H.P.; Tange, S.; Kikuchi, K.; Iida, T. Site specific evaluation oflymphatic vessel sclerosis in lower limb lymphedema patients. Lymphat. Res. Biol. 2018, 16, 360–367. [Google Scholar] [CrossRef]
- Kawanami, O.; Ferrans, V.J.; Fulmer, J.D.; Crystal, R.G. Ultrastructure of pulmonary mast cells in patients with fibrotic lung disorders. Lab. Investig. 1979, 40, 717–734. [Google Scholar]
- Trabucchi, E.; Radaelli, E.; Marazzi, M.; Foschi, D.; Musazzi, M.; Veronesi, A.M.; Montorsi, W. The role of mast cells in wound healing. Int. J. Tissue React. 1988, 10, 367–372. [Google Scholar]
- Leroy, E.C.; Smith, E.A.; Kahaleh, M.B.; Trojanowska, M.; Silver, R.M. A strategy for determining the pathogenesis of systemic sclerosis: Is transforming growth factor beta the answer? Arthritis Rheum. 1989, 32, 817–825. [Google Scholar] [CrossRef]
- Claman, H.N. On scleroderma: Mast cells, endothelial cells and fibroblasts. JAMA 1989, 262, 1206–1209. [Google Scholar] [CrossRef]
- Schwartz, L.B. Mediators of human mast cells and human mast cell subsets. Ann. Allergy 1987, 58, 226–235. [Google Scholar]
- Schwartz, L.B.; Lewis, R.A.; Seldin, D.; Austen, K.F. Acid hydrolases and tryptase from secretory granules of dispersed human lung mast cells. J. Immunol. 1981, 126, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Schechter, N.M.; Choi, J.K.; Slavin, D.A.; Deresienski, D.T.; Sayama, S.; Dong, G.; Lavker, R.M.; Proud, D.; Lazarus, G.S. Identification of a chymotrypsin-like proteinase in human mast cells. J. Immunol. 1986, 137, 962–970. [Google Scholar] [CrossRef]
- Caughey, G.H.; Lazarus, S.C.; Viro, N.F.; Gold, W.M.; Nabel, J.A. Tryptase and chymase: Comparison of extraction and release in two dog mastocytoma lines. Immunology 1988, 63, 339–344. [Google Scholar] [PubMed]
- Wasse, H.; Naqvi, N.; Husain, A. Impact of mast cell chymase on renal disease progression. J. Curr. Hypertens. Rev. 2012, 8, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Irani, A.M.; Craig, S.S.; DeBlois, G.; Elson, C.O.; Schechter, N.M.; Schwartz, L.B. Deficiency of the tryptase-positive, chymase-negative mast cell type in gastrointestinal mucosa of patients with defective T lymphocyte function. J. Immunol. 1987, 138, 4381–4386. [Google Scholar] [CrossRef]
- Gordon, J.R.; Galli, S.J. Promotion of mouse fibroblast collagen gene expression by mast cells stimulated via the Fc epsilon RI. Role for mast cell-derived transforming growth factor beta and tumor necrosis factor alpha. J. Exp. Med. 1994, 180, 2027–2037. [Google Scholar] [CrossRef]
- Cairns, J.A.; Wallls, A.F. Mast cell tryptase stimulates the synthesis of type I collagen in human lung fibroblasts. J. Clin. Investig. 1997, 99, 1313–1321. [Google Scholar] [CrossRef]
- Frangogiannis, N. Transforming growth factor-β in tissue fibrosis. J. Exp. Med. 2020, 217, e20160103. [Google Scholar] [CrossRef]
- Bonner, J.C. Regulation of PDGF and its receptors in fibrotic diseases. Cytokine Growth Factor Rev. 2004, 15, 255–273. [Google Scholar] [CrossRef]
- Paulsson, Y.; Austgulen, R.; Hofsli, E.; Heldin, C.H.; Westermark, B.; Nissen-Meyer, J. Tumor necrosis factor-induced expression of platelet-derived growth factor A-chain messenger RNA in fibroblasts. Exp. Cell Res. 1989, 180, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Battegay, E.J.; Raines, E.W.; Colbert, T.; Ross, R. TNF-alpha stimulation of fibroblast proliferation. Dependence on platelet-derived growth factor (PDGF) secretion and alteration of PDGF receptor expression. J. Immunol. 1995, 154, 6040–6947. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Akazawa, H.; Tamagawa, M.; Ito, K.; Yasuda, N.; Kudo, Y.; Yamamoto, R.; Ozawa, Y.; Fujimoto, M.; Wang, P.; et al. Cardiac mast cells cause atrial fibrillation through PDGF-A-mediated fibrosis in pressure-overloaded mouse hearts. J. Clin. Investig. 2010, 120, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Narari, M.; Nathan, N.C.; Lüdke, A.; Li, S.H.; Guo, J.; Weisel, R.D.; Li, R.K. Mast cells promote proliferation and migration and inhibit differentiationof mesenchymal stem cells through PDGF. J. Mol. Cell. Cardiol. 2016, 94, 32–42. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuri, T.; Jin, D.; Takai, S.; Ueda, K. Tryptase-Positive Mast Cells Promote Adipose Fibrosis in Secondary Lymphedema through PDGF. Curr. Issues Mol. Biol. 2023, 45, 8027-8039. https://doi.org/10.3390/cimb45100507
Nuri T, Jin D, Takai S, Ueda K. Tryptase-Positive Mast Cells Promote Adipose Fibrosis in Secondary Lymphedema through PDGF. Current Issues in Molecular Biology. 2023; 45(10):8027-8039. https://doi.org/10.3390/cimb45100507
Chicago/Turabian StyleNuri, Takashi, Denan Jin, Shinji Takai, and Koichi Ueda. 2023. "Tryptase-Positive Mast Cells Promote Adipose Fibrosis in Secondary Lymphedema through PDGF" Current Issues in Molecular Biology 45, no. 10: 8027-8039. https://doi.org/10.3390/cimb45100507
APA StyleNuri, T., Jin, D., Takai, S., & Ueda, K. (2023). Tryptase-Positive Mast Cells Promote Adipose Fibrosis in Secondary Lymphedema through PDGF. Current Issues in Molecular Biology, 45(10), 8027-8039. https://doi.org/10.3390/cimb45100507