
Implications of Activating the ANT2/mTOR/PGC-1α Feedback Loop: Insights into Mitochondria-Mediated Injury in Hypoxic Myocardial Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Integration for Differentially Expressed miRNAs (DEMs)
2.2. Cells Culture
2.3. Oxygen and Glucose Deprivation (OGD) Cells Models
2.4. Cell Transfection
2.5. Luciferase Reporter Assays
2.6. Measurement of Mitochondrial Membrane Potential (MMP)
2.7. Intracellular ROS Levels
2.8. Flow Cytometry
2.9. Animal AMI Model
2.10. Hematoxylin–Eosin (HE) Staining
2.11. Electrocardiography
2.12. Total RNA Extraction and Real-Time Quantitative PCR (RT-qPCR)
2.13. Western Blot
2.14. Prediction of miRNAs Targeted ANT2
2.15. Statistical Analysis
3. Results
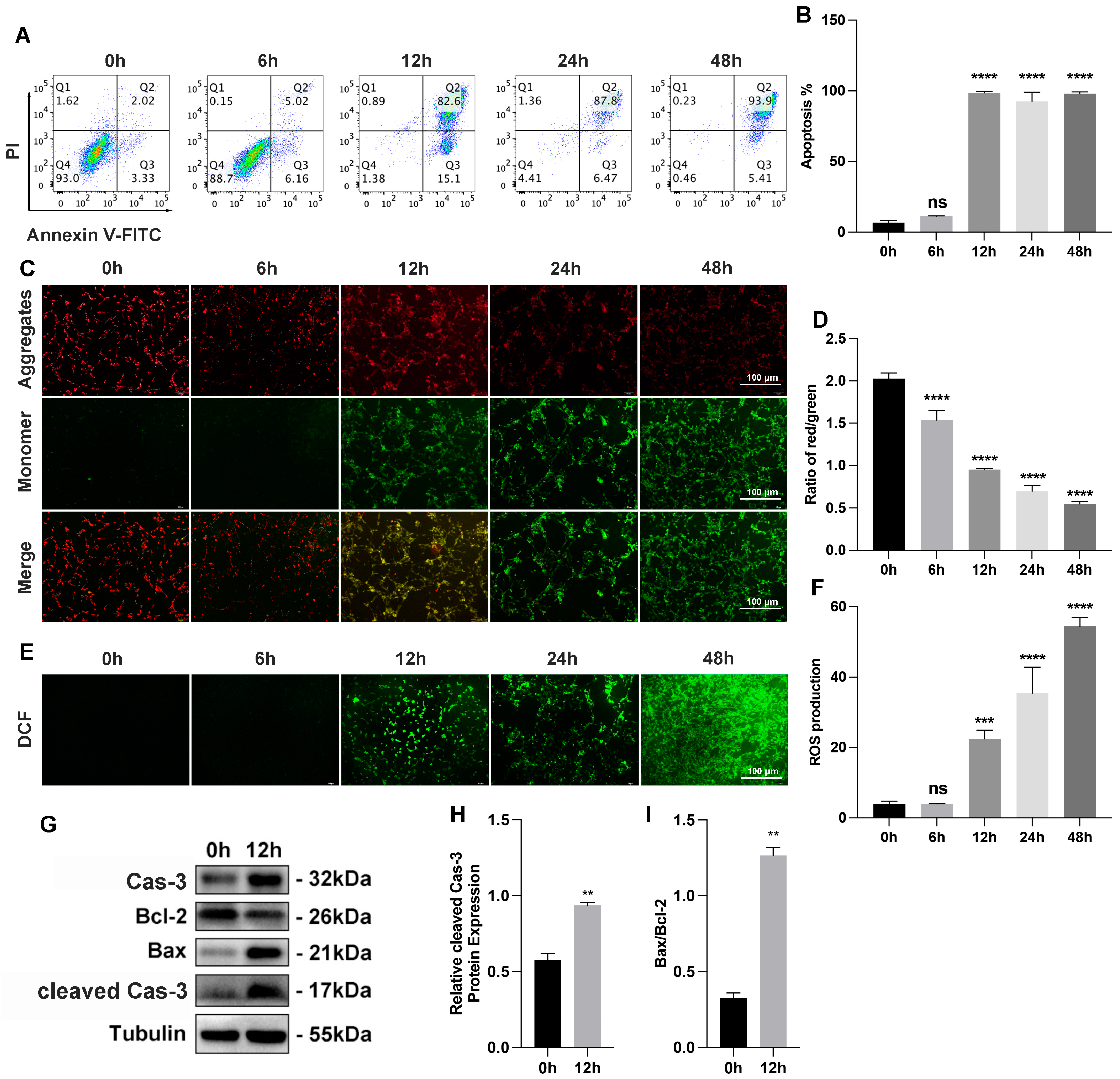
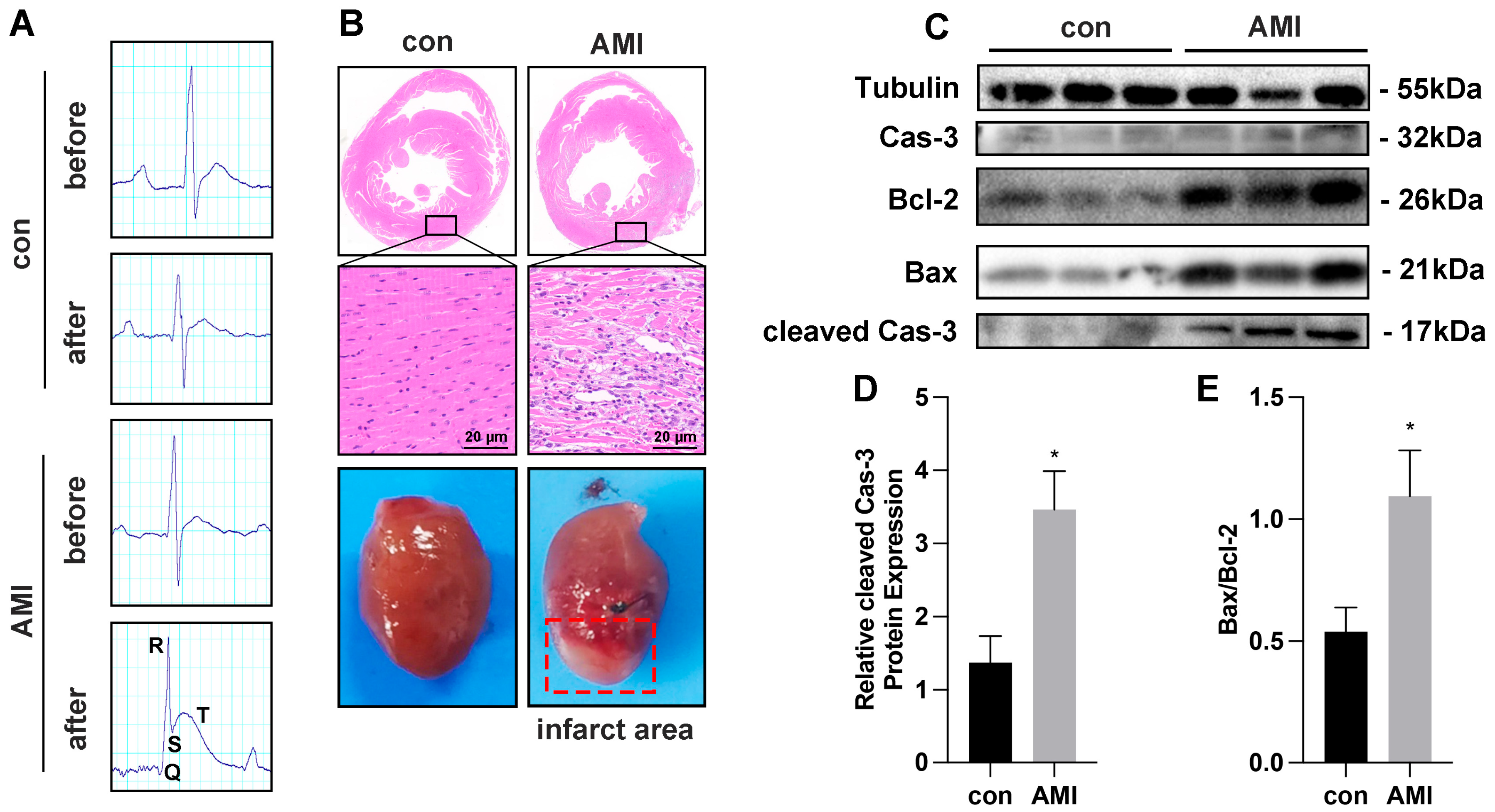
3.1. Myocardial Cells Injury Induced by OGD In Vitro and by Ischemia–Hypoxia In Vivo
3.2. ANT2 siRNA Shows a Protection for AC16 Cells from Injury Caused by OGD
3.3. ANT2 Was Targeted and Regulated by miR-1203 in Human AC16 Cardiocytes
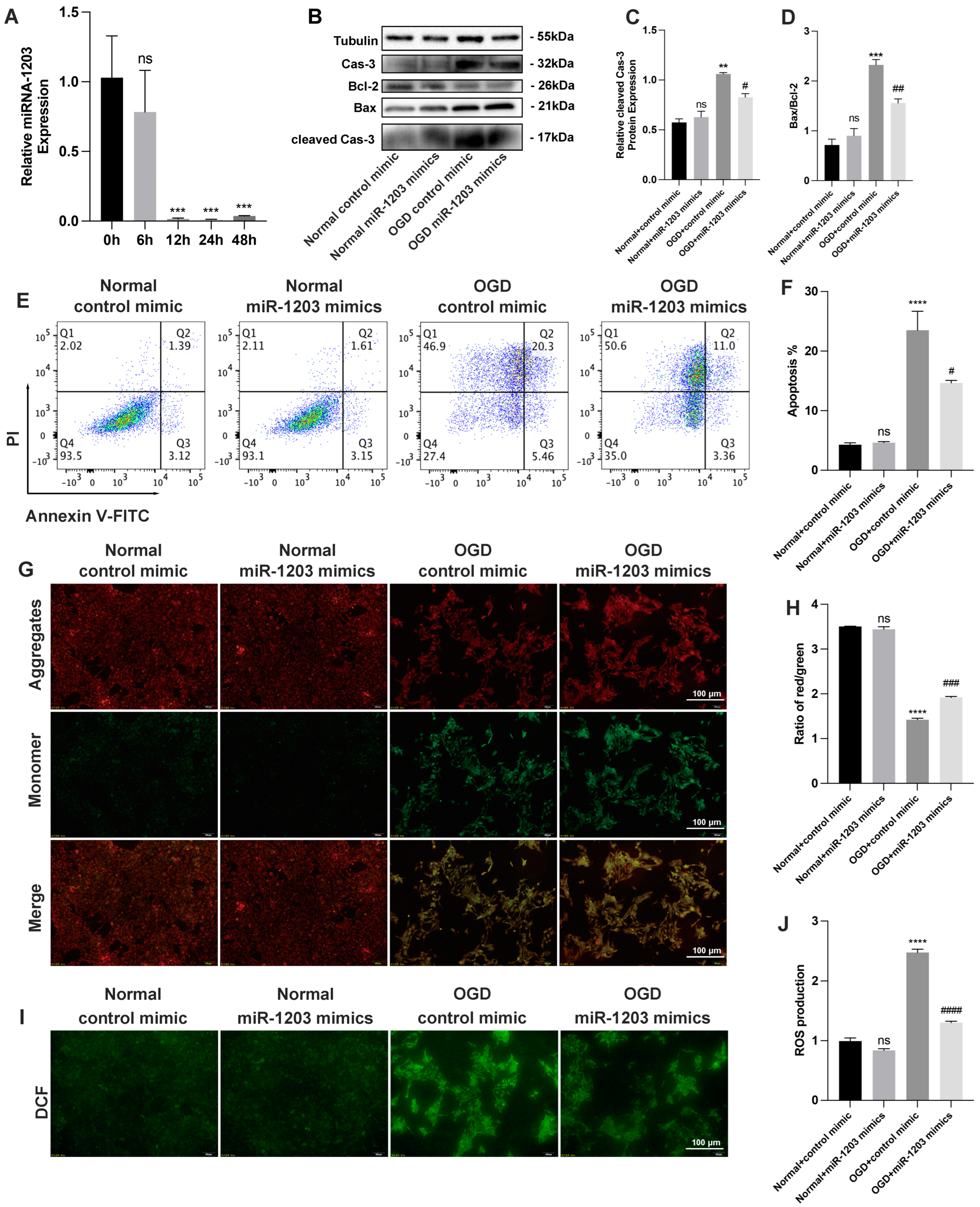
3.4. miR-1203 Mimics Show Potential as Protective Agents against OGD-Induced Injury in AC16 Cells
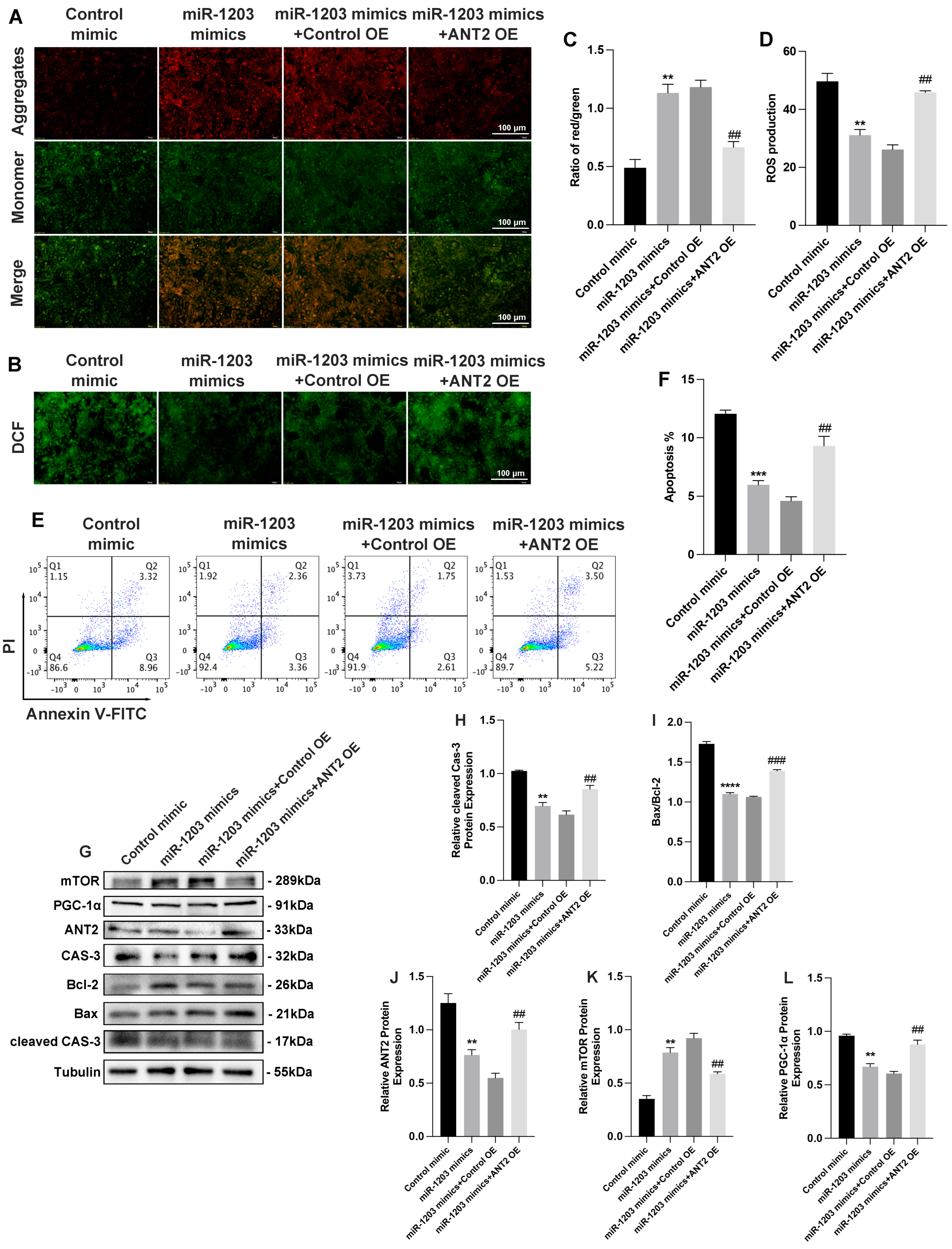
3.5. ANT2 Overexpression Counteracted the Effect of miR-1203
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reed, G.W.; Rossi, J.E.; Cannon, C.P. Acute myocardial infarction. Lancet 2017, 389, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.A.; Alam Khan, S.; Safdar, S.; Taseer, I.-U.-H. Chest Pain as a presenting complaint in patients with acute myocardial infarction (AMI). Pak. J. Med. Sci. 2013, 29, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Sossalla, S.; Vollmann, D. Arrhythmia-Induced Cardiomyopathy. Dtsch. Ärzteblatt Int. 2018, 115, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, M.; Jessup, M.; Mullens, W.; Reza, N.; Shah, A.M.; Sliwa, K.; Mebazaa, A. Acute heart failure. Nat. Rev. Dis. Primers 2020, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Kapur, N.K.; Thayer, K.L.; Zweck, E. Cardiogenic Shock in the Setting of Acute Myocardial Infarction. Methodist. Debakey Cardiovasc. J. 2020, 16, 16–21. [Google Scholar] [CrossRef]
- Su, S.-A.; Zhang, Y.; Li, W.; Xi, Y.; Lu, Y.; Shen, J.; Ma, Y.; Wang, Y.; Shen, Y.; Xie, L.; et al. Cardiac Piezo1 Exacerbates Lethal Ventricular Arrhythmogenesis by Linking Mechanical Stress with Ca2+ Handling after Myocardial Infarction. Research 2023, 6, 0165. [Google Scholar] [CrossRef]
- Heusch, G. Myocardial ischaemia-reperfusion injury and cardioprotection in perspective. Nat. Rev. Cardiol. 2020, 17, 773–789. [Google Scholar] [CrossRef]
- Venugopal, H.; Hanna, A.; Humeres, C.; Frangogiannis, N.G. Properties and Functions of Fibroblasts and Myofibroblasts in Myocardial Infarction. Cells 2022, 11, 1386. [Google Scholar] [CrossRef]
- Seropian, I.M.; Toldo, S.; Van Tassell, B.W.; Abbate, A. Anti-Inflammatory Strategies for Ventricular Remodeling Following ST-Segment Elevation Acute Myocardial Infarction. J. Am. Coll. Cardiol. 2014, 63, 1593–1603. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Wang, S.; Cheng, H.; Xu, L.; Pei, G.; Wang, Y.; Fu, C.; Jiang, Y.; He, C.; et al. Signaling pathways and targeted therapy for myocardial infarction. Signal Transduct. Target. Ther. 2022, 7, 78. [Google Scholar] [CrossRef]
- Li, B.; Li, T.; Han, C.; Liu, Y.; Zhong, X.; Cao, Y.; Deng, Y. Potential of Dragon’s Blood as a Space Radiation Protectant Especially on Brain-Liver Bystander Effect. Space Sci. Technol. 2022, 2022, 9791283. [Google Scholar] [CrossRef]
- Ramachandra, C.J.A.; Hernandez-Resendiz, S.; Crespo-Avilan, G.E.; Lin, Y.-H.; Hausenloy, D.J. Mitochondria in acute myocardial infarction and cardioprotection. EBioMedicine 2020, 57, 102884. [Google Scholar] [CrossRef] [PubMed]
- Chevrollier, A.; Loiseau, D.; Reynier, P.; Stepien, G. Adenine nucleotide translocase 2 is a key mitochondrial protein in cancer metabolism. Biochim. Biophys. Acta (BBA)-Bioenerg. 2011, 1807, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Heger, J.; Abdallah, Y.; Shahzad, T.; Klumpe, I.; Piper, H.M.; Schultheiss, H.P.; Schlüter, K.D.; Schulz, R.; Euler, G.; Dörner, A. Transgenic overexpression of the adenine nucleotide translocase 1 protects cardiomyocytes against TGFβ1-induced apoptosis by stabilization of the mitochondrial permeability transition pore. J. Mol. Cell. Cardiol. 2012, 53, 73–81. [Google Scholar] [CrossRef]
- Klingenberg, M. The ADP and ATP transport in mitochondria and its carrier. Biochim. Biophys. Acta (BBA)-Bioenerg. 2008, 1778, 1978–2021. [Google Scholar] [CrossRef]
- Flierl, A.; Schriner, S.E.; Hancock, S.; Coskun, P.E.; Wallace, D.C. The mitochondrial adenine nucleotide transporters in myogenesis. Free Radic. Biol. Med. 2022, 188, 312–327. [Google Scholar] [CrossRef]
- Kokoszka, J.E.; Waymire, K.G.; Flierl, A.; Sweeney, K.M.; Angelin, A.; MacGregor, G.R.; Wallace, D.C. Deficiency in the mouse mitochondrial adenine nucleotide translocator isoform 2 gene is associated with cardiac noncompaction. Biochim. Biophys. Acta (BBA)-Bioenerg. 2016, 1857, 1203–1212. [Google Scholar] [CrossRef]
- Chevrollier, A.; Loiseau, D.; Gautier, F.; Malthièry, Y.; Stepien, G. ANT2 expression under hypoxic conditions produces opposite cell-cycle behavior in 143B and HepG2 cancer cells. Mol. Carcinog. 2005, 42, 1–8. [Google Scholar] [CrossRef]
- Lu, Y.W.; Acoba, M.G.; Selvaraju, K.; Huang, T.C.; Nirujogi, R.S.; Sathe, G.; Pandey, A.; Claypool, S.M. Human adenine nucleotide translocases physically and functionally interact with respirasomes. Mol. Biol. Cell 2017, 28, 1489–1506. [Google Scholar] [CrossRef]
- Gouriou, Y.; Alam, M.R.; Harhous, Z.; Crola Da Silva, C.; Baetz, D.B.; Badawi, S.; Lefai, E.; Rieusset, J.; Durand, A.; Harisseh, R.; et al. ANT2-Mediated ATP Import into Mitochondria Protects against Hypoxia Lethal Injury. Cells 2020, 9, 2542. [Google Scholar] [CrossRef]
- Doench, J.G.; Petersen, C.P.; Sharp, P.A. siRNAs can function as miRNAs. Genes Dev. 2003, 17, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhou, Y.; Richards, A.M. Effective tools for RNA-derived therapeutics: siRNA interference or miRNA mimicry. Theranostics 2021, 11, 8771–8796. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, M.; Greiser, U.; Wall, J.G.; O’Brien, T.; Pandit, A. Interference: An alteRNAtive therapy following acute myocardial infarction. Trends Pharmacol. Sci. 2012, 33, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Chang, Y.-H.; Xiong, Q.; Zhang, P.; Zhang, L.; Somasundaram, P.; Lepley, M.; Swingen, C.; Su, L.; Wendel, J.S.; et al. Cardiac Repair in a Porcine Model of Acute Myocardial Infarction with Human Induced Pluripotent Stem Cell-Derived Cardiovascular Cells. Cell Stem Cell 2014, 15, 750–761. [Google Scholar] [CrossRef]
- McKeever, R.G.; Vearrier, D.; Jacobs, D.; LaSala, G.; Okaneku, J.; Greenberg, M.I. K2—Not the Spice of Life; Synthetic Cannabinoids and ST Elevation Myocardial Infarction: A Case Report. J. Med. Toxicol. 2015, 11, 129–131. [Google Scholar] [CrossRef]
- Zhang, C.-X.; Cheng, Y.; Liu, D.-Z.; Liu, M.; Cui, H.; Zhang, B.-l.; Mei, Q.-B.; Zhou, S.-Y. Mitochondria-targeted cyclosporin A delivery system to treat myocardial ischemia reperfusion injury of rats. J. Nanobiotechnol. 2019, 17, 18. [Google Scholar] [CrossRef]
- Gao, S.-Y.; Wang, Q.-J.; Ji, Y.-B. Effect of solanine on the membrane potential of mitochondria in HepG2 cells and [Ca2+]i in the cells. World J. Gastroenterol. 2006, 12, 3359–3367. [Google Scholar] [CrossRef]
- Belzacq-Casagrande, A.-S.; Vieira, H.; Kroemer, G.; Brenner, C. The adenine nucleotide translocator in apoptosis. Biochimie 2002, 84, 167–176. [Google Scholar] [CrossRef]
- Peng, S.; Zhang, X.; Yu, L.; Xu, Y.; Zhou, Y.; Qian, S.; Cao, X.; Ye, X.; Yang, J.; Jia, W.; et al. NF-κB regulates brown adipocyte function through suppression of ANT2. Acta Pharm. Sin. B 2022, 12, 1186–1197. [Google Scholar] [CrossRef]
- Hang, T.; Lumpuy-Castillo, J.; Goikoetxea-Usandizaga, N.; Azkargorta, M.; Aldamiz, G.; Martinez-Milla, J.; Forteza, A.; Cortina, J.M.; Egido, J.; Elortza, F.; et al. Potential Role of the mTORC1-PGC1α-PPARα Axis under Type-II Diabetes and Hypertension in the Human Heart. Int. J. Mol. Sci. 2023, 24, 8629. [Google Scholar] [CrossRef] [PubMed]
- Gavalda-Navarro, A.; Villena, J.A.; Planavila, A.; Vinas, O.; Mampel, T. Expression of adenine nucleotide translocase (ANT) isoform genes is controlled by PGC-1α through different transcription factors. J. Cell Physiol. 2014, 229, 2126–2136. [Google Scholar] [CrossRef] [PubMed]
- Grunweller, A.; Hartmann, R.K. RNA interference as a gene-specific approach for molecular medicine. Curr. Med. Chem. 2005, 12, 3143–3161. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-K.; Zhu, J.-Q.; Zhang, J.-T.; Li, Q.; Li, Y.; He, J.; Qin, Y.-W.; Jing, Q. Circulating microRNA: A novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur. Heart J. 2010, 31, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Zhang, R.; Li, Y.; Pu, J.; Lu, Y.; Jiao, J.; Li, K.; Yu, B.; Li, Z.; Wang, R.; et al. Circulating microRNA-1 as a potential novel biomarker for acute myocardial infarction. Biochem. Biophys. Res. Commun. 2010, 391, 73–77. [Google Scholar] [CrossRef]
- Huang, W.; Tian, S.-S.; Hang, P.-Z.; Sun, C.; Guo, J.; Du, Z.-M. Combination of microRNA-21 and microRNA-146a Attenuates Cardiac Dysfunction and Apoptosis during Acute Myocardial Infarction in Mice. Mol. Ther.-Nucleic Acids 2016, 5, e296. [Google Scholar] [CrossRef]
- Yang, X.; Qin, Y.; Shao, S.; Yu, Y.; Zhang, C.; Dong, H.; Lv, G.; Dong, S. MicroRNA-214 Inhibits Left Ventricular Remodeling in an Acute Myocardial Infarction Rat Model by Suppressing Cellular Apoptosis via the Phosphatase and Tensin Homolog (PTEN). Int. Heart J. 2016, 57, 247–250. [Google Scholar] [CrossRef]
- Shyu, K.G.; Wang, B.W. MicroRNA-208a increases myocardial fibrosis via endoglin in volume overload heart. Eur. Heart J. 2013, 34, 1752. [Google Scholar] [CrossRef]
- Ma, J.L.; Guo, W.L.; Chen, X.M. Overexpressing microRNA-150 attenuates hypoxia-induced human cardiomyocyte cell apoptosis by targeting glucose-regulated protein-94. Mol. Med. Rep. 2018, 17, 4181–4186. [Google Scholar] [CrossRef]
- Zhang, J.; Qiu, W.; Ma, J.; Wang, Y.; Hu, Z.; Long, K.; Wang, X.; Jin, L.; Tang, Q.; Tang, G.; et al. miR-27a-5p Attenuates Hypoxia-induced Rat Cardiomyocyte Injury by Inhibiting Atg7. Int. J. Mol. Sci. 2019, 20, 2418. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Q.; Wang, Z.; Wang, F.; Guo, X.; Li, G. Circulating microRNA profiles and the identification of miR-593 and miR-511 which directly target the PROP1 gene in children with combined pituitary hormone deficiency. Int. J. Mol. Med. 2015, 35, 358–366. [Google Scholar] [CrossRef]
- Haldrup, C.; Kosaka, N.; Ochiya, T.; Borre, M.; Hoyer, S.; Orntoft, T.F.; Sorensen, K.D. Profiling of circulating microRNAs for prostate cancer biomarker discovery. Drug Deliv. Transl. Res. 2014, 4, 19–30. [Google Scholar] [CrossRef]
- Xu, H.B.; Zheng, Y.F.; Wu, D.; Li, Y.; Zhou, L.N.; Chen, Y.G. microRNA-1203 targets and silences cyclophilin D to protect human endometrial cells from oxygen and glucose deprivation-re-oxygenation. Aging 2020, 12, 3010–3024. [Google Scholar] [CrossRef]
- Tan, H.; Song, Y.; Chen, J.; Zhang, N.; Wang, Q.; Li, Q.; Gao, J.; Yang, H.; Dong, Z.; Weng, X.; et al. Platelet-like Fusogenic Liposome-Mediated Targeting Delivery of miR-21 Improves Myocardial Remodeling by Reprogramming Macrophages Post Myocardial Ischemia-Reperfusion Injury. Adv. Sci. 2021, 8, 2100787. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, P.; Hong, J.; Li, N.; Zhang, Y.; Deng, Y. Deep Membrane Proteome Profiling of Rat Hippocampus in Simulated Complex Space Environment by SWATH. Space Sci. Technol. 2021, 2021, 9762372. [Google Scholar] [CrossRef]
- Li, Y.; Huang, L.; Iqbal, J.; Deng, Y. Investigation on P-Glycoprotein Function and Its Interacting Proteins under Simulated Microgravity. Space Sci. Technol. 2021, 2021, 9835728. [Google Scholar] [CrossRef]
- Cook, G.J.R.; Goh, V. What can artificial intelligence teach us about the molecular mechanisms underlying disease? Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2715–2721. [Google Scholar] [CrossRef]
- Chen, A.; Xie, Y.; Wang, Y.; Li, L. Knowledge Graph-Based Image Recognition Transfer Learning Method for On-Orbit Service Manipulation. Space Sci. Technol. 2021, 2021, 9807452. [Google Scholar] [CrossRef]
- Waisberg, E.; Ong, J.; Paladugu, P.; Kamran, S.A.; Zaman, N.; Lee, A.G.; Tavakkoli, A. Challenges of Artificial Intelligence in Space Medicine. Space Sci. Technol. 2022, 2022, 9852872. [Google Scholar] [CrossRef]
- Khera, R.; Haimovich, J.; Hurley, N.C.; McNamara, R.; Spertus, J.A.; Desai, N.; Rumsfeld, J.S.; Masoudi, F.A.; Huang, C.; Normand, S.-L.; et al. Use of Machine Learning Models to Predict Death after Acute Myocardial Infarction. JAMA Cardiol. 2021, 6, 633–641. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/miRNA | Primer Sequences (5′-3′) |
|---|---|
| U6 | Forward: GCTTGCTTCGGCAGCACATATAC |
| Reverse: TGCATGTCATCCTTGCTCAGGG | |
| hsa-miR-1203 | Forward: GGAGCCAGGATGCAGCTC |
| Reverse: GCGACACAGAATTATACGACTCAC | |
| GAPDH | hsa-Forward: TCATTGACCTCAACTACATGGT |
| hsa-Reverse: CTAAGCAGTTGGTGGTGCAG | |
| mmu-Forward: TCATTGACCTCAACTACATGGT | |
| mmu-Reverse: CTAAGCAGTTGGTGGTGCAG | |
| ANT2 | hsa-Forward: CTACTTTGCAGGGAATCTGG |
| hsa-Reverse: GACACGTTAAAGCCTTGGTAC | |
| mmu-Forward: CTACTTTGCAGGGAACCTGG | |
| mmu-Reverse: GACACATTAAAGCCTTGGTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Yang, Y.; Zhu, Z.; Chen, Z.; Huang, D. Implications of Activating the ANT2/mTOR/PGC-1α Feedback Loop: Insights into Mitochondria-Mediated Injury in Hypoxic Myocardial Cells. Curr. Issues Mol. Biol. 2023, 45, 8633-8651. https://doi.org/10.3390/cimb45110543
Zhang M, Yang Y, Zhu Z, Chen Z, Huang D. Implications of Activating the ANT2/mTOR/PGC-1α Feedback Loop: Insights into Mitochondria-Mediated Injury in Hypoxic Myocardial Cells. Current Issues in Molecular Biology. 2023; 45(11):8633-8651. https://doi.org/10.3390/cimb45110543
Chicago/Turabian StyleZhang, Meng, Yuanzhan Yang, Zhu Zhu, Zixuan Chen, and Dongyang Huang. 2023. "Implications of Activating the ANT2/mTOR/PGC-1α Feedback Loop: Insights into Mitochondria-Mediated Injury in Hypoxic Myocardial Cells" Current Issues in Molecular Biology 45, no. 11: 8633-8651. https://doi.org/10.3390/cimb45110543
APA StyleZhang, M., Yang, Y., Zhu, Z., Chen, Z., & Huang, D. (2023). Implications of Activating the ANT2/mTOR/PGC-1α Feedback Loop: Insights into Mitochondria-Mediated Injury in Hypoxic Myocardial Cells. Current Issues in Molecular Biology, 45(11), 8633-8651. https://doi.org/10.3390/cimb45110543