
Exploring Genetic and Epigenetic Changes in Lingonberry Using Molecular Markers: Implications for Clonal Propagation




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions and Morphological Data
2.2. DNA Isolation
2.3. Clonal Fidelity Experiment
2.4. Methylation-Sensitive Amplification Polymorphism (MSAP) Assay
2.4.1. Digestion
2.4.2. Ligation
2.4.3. Preamplification
2.4.4. Selective Amplification
2.4.5. Polyacrylamide Gel Electrophoresis (PAGE)
2.4.6. Profiling Scoring and Data Analysis
2.4.7. Statistical Analysis
3. Results
3.1. Morphological Characteristics
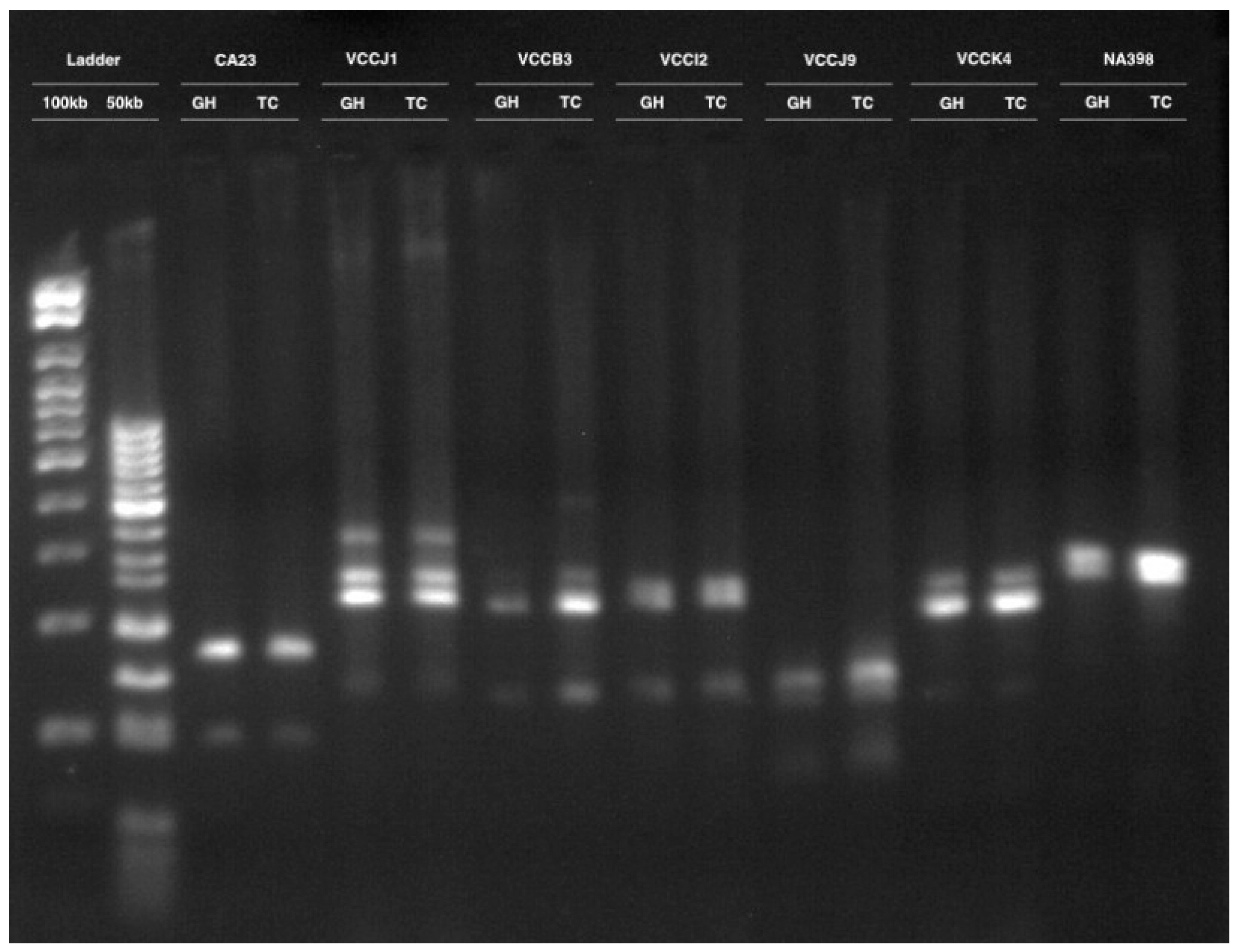
3.2. Clonal Fidelity
3.3. DNA Methylation Pattern
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rohde, A.; Junttila, O. Remembrances of an Embryo: Long-Term Effects on Phenology Traits in Spruce. New Phytol. 2008, 177, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Igamberdiev, A.U.; Debnath, S.C. Tissue Culture-Induced DNA Methylation in Crop Plants: A Review. Mol. Biol. Rep. 2021, 48, 823–841. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.K. Epigenetic Regulation of Stress Responses in Plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujor, O.C.; Ginies, C.; Popa, V.I.; Dufour, C. Phenolic Compounds and Antioxidant Activity of Lingonberry (Vaccinium Vitis-Idaea L.) Leaf, Stem and Fruit at Different Harvest Periods. Food Chem. 2018, 252, 356–365. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, Z.; Liu, D.; Shi, G.; Liu, D.; Yang, Y.; Gu, H.; Yang, L.; Zhou, Z. Natural Antioxidant of Rosemary Extract Used as an Additive in the Ultrasound-Assisted Extraction of Anthocyanins from Lingonberry (Vaccinium Vitis-Idaea L.) Pomace. Ind. Crops Prod. 2019, 138, 111425. [Google Scholar] [CrossRef]
- Debnath, S.C.; Arigundam, U. In Vitro Propagation Strategies of Medicinally Important Berry Crop, Lingonberry (Vaccinium Vitis-Idaea L.). Agronomy 2020, 10, 744. [Google Scholar] [CrossRef]
- Kelly, E.; Vyas, P.; Weber, J.T. Biochemical Properties and Neuroprotective Effects of Compounds in Various Species of Berries. Molecules 2018, 23, 26. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Park, S.W. Edible Berries: Bioactive Components and Their Effect on Human Health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Holloway, P.S. Rooting of Lingonberry, Vaccinium Vitis-Idaea, Stem Cuttings. Plant Prop. 1985, 31, 7–9. [Google Scholar]
- Arigundam, U.; Variyath, A.M.; Siow, Y.L.; Marshall, D.; Debnath, S.C. Liquid Culture for Efficient In Vitro Propagation of Adventitious Shoots in Wild Vaccinium Vitis-Idaea Ssp. Minus (Lingonberry) Using Temporary Immersion and Stationary Bioreactors. Sci. Hortic. 2020, 264, 109199. [Google Scholar] [CrossRef]
- Larkin, P.J.; Scowcroft, W.R. Somaclonal Variation—A Novel Source of Variability from Cell Cultures for Plant Improvement. Theor. Appl. Gen. 1981, 60, 197–214. [Google Scholar] [CrossRef]
- Azman, A.S.; Mhiri, C.; Tam, S.M. Transposable Elements and the Detection of Somaclonal Variation in Plant Tissue Culture: A Review. Malaysian App. Biol. 2014, 43, 1–12. [Google Scholar]
- Hirsch, S.; Baumberger, R.; Grossniklaus, U. Epigenetic Variation, Inheritance, and Selection in Plant Populations. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Murthy, H.N.; Chakrabarthy, D.; Paek, K.Y. Detection of Epigenetic Variation in Tissue-Culture-Derived Plants of Doritaenopsis by Methylation-Sensitive Amplification Polymorphism (MSAP) Analysis. In Vitro Cell. Dev. Biol.-Plant 2009, 45, 104–108. [Google Scholar] [CrossRef]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.Á.; Abarca, D.; et al. Epigenetic Regulation of Adaptive Responses of Forest Tree Species to the Environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Hatorangan, M.R.; Laenen, B.; Steige, K.A.; Slotte, T.; Köhler, C. Rapid Evolution of Genomic Imprinting in Two Species of the Brassicaceae. Plant Cell 2016, 28, 1815–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, I.R.; Jacobsen, S.E. Epigenetic Inheritance in Plants. Nature 2007, 447, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.; O’Malley, R.C.; Tonti-Filippini, J.; Gregory, B.D.; Berry, C.C.; Millar, A.H.; Ecker, J.R. Highly Integrated Single-Base Resolution Maps of the Epigenome in Arabidopsis. Cell 2008, 133, 523–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maunakea, A.K.; Chepelev, I.; Cui, K.; Zhao, K. Intragenic DNA Methylation Modulates Alternative Splicing by Recruiting MeCP2 to Promote Exon Recognition. Cell Res. 2013, 23, 1256–1269. [Google Scholar] [CrossRef] [Green Version]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for Plant Improvement: Current Knowledge and Modeling Avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Reyna-López, G.E.; Simpson, J.; Ruiz-Herrera, J. Differences in DNA Methylation Patterns Are Detectable during the Dimorphic Transition of Fungi by Amplification of Restriction Polymorphisms. Mol. Gen. Genet. 1997, 253, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Pérez, R.; Bazaga, P.; Medrano, M.; Herrera, C.M. MSAP Markers and Global Cytosine Methylation in Plants: A Literature Survey and Comparative Analysis for a Wild-Growing Species. Mol. Ecol. Resour. 2016, 16, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boches, P.S.; Bassil, N.V.; Rowland, L.J. Microsatellite Markers for Vaccinium from EST and Genomic Libraries. Mol. Ecol. Notes 2005, 5, 657–660. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A New Technique for DNA Fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulneček, J.; Kovařík, A. How to Interpret Methylation Sensitive Amplified Polymorphism (MSAP) Profiles? BMC Genet. 2014, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Şahin, M.; Aybek, E. Jamovi: An easy to use statistical software for the social scientists. Int. J. Assess. Tools Educ. 2019, 6, 670–692. [Google Scholar] [CrossRef] [Green Version]
- Mohan Jain, S. Tissue Culture-Derived Variation in Crop Improvement. Euphytica 2001, 118, 153–166. [Google Scholar] [CrossRef]
- Machczyńska, J.; Zimny, J.; Bednarek, P.T. Tissue Culture-Induced Genetic and Epigenetic Variation in Triticale (× Triticosecale Spp. Wittmack Ex A. Camus 1927) Regenerants. Plant Mol. Biol. 2015, 89, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Jaligot, E.; Beulé, T.; Rival, A. Methylation-Sensitive RFLPs: Characterisation of Two Oil Palm Markers Showing Somaclonal Variation-Associated Polymorphism. Theor. Appl. Genet. 2002, 104, 1263–1269. [Google Scholar] [CrossRef]
- Miguel, C.; Marum, L. An Epigenetic View of Plant Cells Cultured In Vitro: Somaclonal Variation and Beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef] [Green Version]
- Peraza-Echeverria, S.; Herrera-Valencia, V.A.; Kay, A.J. Detection of DNA Methylation Changes in Micropropagated Banana Plants Using Methylation-Sensitive Amplification Polymorphism (MSAP). Plant Sci. 2001, 161, 359–367. [Google Scholar] [CrossRef]
- Baránek, M.; Čechová, J.; Raddová, J.; Holleinová, V.; Ondrušíková, E.; Pidra, M. Dynamics and Reversibility of the DNA Methylation Landscape of Grapevine Plants (Vitis Vinifera) Stressed by In Vitro Cultivation and Thermotherapy. PLoS ONE 2015, 10, e0126638. [Google Scholar] [CrossRef]
- Sikdar, A.; Sharma, U.; Barua, R.; Igamberdiev, A.U.; Debnath, S.C. Epigenomic Insight of Lingonberry and Health-Promoting Traits during Micropropagation. Sci. Rep. 2022, 12, 12487. [Google Scholar] [CrossRef] [PubMed]
- Goyali, J.C.; Igamberdiev, A.U.; Debnath, S.C. Propagation Methods Affect Fruit Morphology and Antioxidant Properties but Maintain Clonal Fidelity in Lowbush Blueberry. HortSci 2015, 50, 888–896. [Google Scholar] [CrossRef] [Green Version]
- Rathore, M.S.; Jha, B. DNA Methylation and Methylation Polymorphism in Genetically Stable In Vitro Regenerates of Jatropha Curcas L. Using Methylation-Sensitive AFLP Markers. Appl. Biochem. Biotechnol. 2016, 178, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.; Singh, K.P.; Jhang, T.; Sharma, T.R. Assessment of Clonal Fidelity of Micropropagated Gerbera Plants by ISSR Markers. Sci. Hortic. 2009, 119, 208–211. [Google Scholar] [CrossRef]
- Tiwari, J.; Saurabh, S.; Chandel, P.; Singh, B.; Bhardwaj, V. Analysis of Genetic and Epigenetic Variation in in Vitro Propagated Potato Somatic Hybrid by AFLP and MSAP Markers. Electr. J. Biotechnol. 2013, 16, 5. [Google Scholar] [CrossRef]
- Xu, X.; Li, T.; Li, Y.; Wang, H. Variation of DNA Cytosine Methylation Patterns Among Parent Lines and Reciprocal Hybrids in Hot Pepper. Chem. Eng. Trans. 2015, 46, 1345–1350. [Google Scholar] [CrossRef]
- Richards, E.J. Inherited Epigenetic Variation—Revisiting Soft Inheritance. Nat. Rev. Genet. 2006, 7, 395–401. [Google Scholar] [CrossRef]
- Bond, D.M.; Finnegan, E.J. Passing the Message on: Inheritance of Epigenetic Traits. Trends Plant Sci. 2007, 12, 211–216. [Google Scholar] [CrossRef]
- Mathieu, O.; Reinders, J.; Čaikovski, M.; Smathajitt, C.; Paszkowski, J. Transgenerational Stability of the Arabidopsis Epigenome Is Coordinated by CG Methylation. Cell 2007, 130, 851–862. [Google Scholar] [CrossRef]
- Kalisz, S.; Purugganan, M.D. Epialleles via DNA Methylation: Consequences for Plant Evolution. Trends Ecol. Evol. 2004, 19, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Mirouze, M.; Paszkowski, J. Epigenetic Contribution to Stress Adaptation in Plants. Curr. Opin. Plant Biol. 2011, 14, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, K.; Katakami, H.; Kim, H.-J.; Ogawa, E.; Sano, C.M.; Wada, Y.; Sano, H. Epigenetic Inheritance in Rice Plants. Ann. Bot. 2007, 100, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, M.; Li, Y.; Zhang, Q.; Lindsey, K.; Daniell, H.; Jin, S.; Zhang, X. Multi-Omics Analyses Reveal Epigenomics Basis for Cotton Somatic Embryogenesis through Successive Regeneration Acclimation Process. Plant Biotechnol. J. 2019, 17, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant Regeneration: Cellular Origins and Molecular Mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Liu, H.; Cheng, Z.J.; Su, Y.H.; Han, H.N.; Zhang, Y.; Zhang, X.S. DNA Methylation and Histone Modifications Regulate de Novo Shoot Regeneration in Arabidopsis by Modulating WUSCHEL Expression and Auxin Signaling. PLoS Genet. 2011, 7, e1002243. [Google Scholar] [CrossRef] [Green Version]
- Law, J.A.; Jacobsen, S.E. Establishing, Maintaining and Modifying DNA Methylation Patterns in Plants and Animals. Nat Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef]
- Yang, D.; Huang, Z.; Jin, W.; Xia, P.; Jia, Q.; Yang, Z.; Hou, Z.; Zhang, H.; Ji, W.; Han, R. DNA Methylation: A New Regulator of Phenolic Acids Biosynthesis in Salvia Miltiorrhiza. Ind. Crops Prod. 2018, 124, 402–411. [Google Scholar] [CrossRef]
- Hu, Y.; Morota, G.; Rosa, G.J.; Gianola, D. Prediction of Plant Height in Arabidopsis thaliana Using DNA Methylation Data. Genetics 2015, 201, 779–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grzybkowska, D.; Morończyk, J.; Wójcikowska, B.; Gaj, M.D. Azacitidine (5-AzaC)-Treatment and Mutations in DNA Methylase Genes Affect Embryogenic Response and Expression of the Genes That Are Involved in Somatic Embryogenesis in Arabidopsis. Plant Growth Regul. 2018, 85, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Rival, A.; Jaligot, E.; Beule, T.; Finnegan, E.J. Isolation and Expression Analysis of Genes Encoding MET, CMT, and DRM Methyltransferases in Oil Palm (Elaeis Guineensis Jacq.) in Relation to the ‘Mantled’ Somaclonal Variation. J. Exp. Bot. 2008, 59, 3271–3281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De-la-Peña, C.; Nic-Can, G.I.; Galaz-Ávalos, R.M.; Avilez-Montalvo, R.; Loyola-Vargas, V.M. The Role of Chromatin Modifications in Somatic Embryogenesis in Plants. Front. Plant Sci. 2015, 6, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo Name | Function | Nucleotide Sequences |
|---|---|---|
| Ad. EcoRI | Forward adaptor | 5′-CTG TAG ACT GCG TAC C-3′ |
| Ad. EcoRI | Reverse adaptor | 3′-CAT CTG ACG CAT GGT TAA-5′ |
| Ad. MspI/HpaII | Forward adaptor | 5′-GAT CAT GAG TCC TGC T-3′ |
| Ad. MspI/HpaII | Reverse adaptor | 3′-AGT ACT CAG GAC GAG C-5′ |
| EcoRI (E) | Preselective amplification primer | 5′-GAC TGC GTA CCA ATT CA-3′ |
| MspI/HpaII (MH) | Preselective amplification primer | 5′-ATC ATG AGT CCT GCT CGG-3′ |
| E-TT | Selective amplification primer | 5′-GAC TGC GTA CCA ATT CAT T-3′ |
| E-TG | Selective amplification primer | 5′-GAC TGC GTA CCA ATT CAT G-3′ |
| MH-ATG | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG ATG-3′ |
| MH-AAC | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG AAC-3′ |
| MH-AAG | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG AAG-3′ |
| MH-ACA | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG ACA-3′ |
| MH-ATT | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG ATT-3′ |
| MH-TCC | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG TCC-3′ |
| MH-AAT | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG AAT-3′ |
| MH-TCG | Selective amplification primer | 5′-ATC ATG AGT CCT GCT CGG TCG-3′ |
| Primer Type | Primer Name | Primer Sequence | Annealing Temperature | Bands Present (No.) | Size of Amplified Alleles (bp) |
|---|---|---|---|---|---|
| EST PCR | CA21 | F:TCCGATAACCGTTACCAAGC R:TATACAGCGACACGCCAAAA | 54 | 2 | 110, 230 |
| EST SSR | CA23 | F:GAGAGGGTTTCGAGGAGGAG R:GTTTAGAAACGGGACTGTGAGACG | 60 | 2 | 100, 175 |
| EST SSR | CA169 | F:TAGTGGAGGGTTTTGCTTGG R:GTTTATCGAAGCGAAGGTCAAAGA | 54 | 2 | 260, 350 |
| EST SSR | CA421 | F:TCAAATTCAAAGCTCAAAATCAA R:GTTTAAGGATGATCCCGAAGCTCT | 60 | 2 | 175, 250 |
| EST SSR | NA398 | F:TCCTTGCTCCAGTCCTATGC R:GTTTCCTTCCACTCCAAGATGC | 60 | 2 | 145, 200 |
| EST SSR | NA1040 | F:GCAACTCCCAGACTTTCTCC R:GTTTAGTCAGCAGGGTGCACAA | 56 | 3 | 150, 210, 350 |
| GSSR | VCCB3 | F:CCTTCGATCTTGTTCCTTGC R:GTTTGATGCAATTGAGGTGGAGA | 62 | 3 | 125, 270, 300 |
| GSSR | VCCI2 | F:AGGCGTTTTTGAGGCTAACA R:TAAAAGTTCGGCTCGTTTGC | 62 | 3 | 130, 300, 325 |
| GSSR | VCCJ9 | F:GCGAAGAACTTCCGTCAAAA R:GTGAGGGCACAAAGCTCTC | 60 | 3 | 75, 120, 135 |
| GSSR | VCCJ1 | F:CTCATGGGTTCCCATAGACAA R:TGCAGTGAGGCAAAAGATTG | 62 | 3 | 275, 300, 350 |
| GSSR | VCCK4 | F:CCTCCACCCCACTTTCATTA R:GCACACAGGTCCAGTTTTTG | 62 | 3 | 100, 140, 150 |
| GSSR | VCCS10 | F:ATTTGGTGTGAAACCCCTGA R:GTTTGCGGCTATATCCGTGTTTGT | 60 | 3 | 150, 175, 215 |
| DNA Bands | Erntedank | Hybrid (H1 = HY GH, HY GR) | ||
|---|---|---|---|---|
| Greenhouse | Tissue Culture | Greenhouse | Tissue Culture | |
| Type 1 | 228 | 189 | 240 | 204 |
| Type 2 | 54 | 80 | 48 | 86 |
| Type 3 | 46 | 48 | 46 | 50 |
| Type 4 | 16 | 12 | 19 | 24 |
| Total analyzed bands | 344 | 329 | 353 | 364 |
| Total methylated bands | 116 | 140 | 113 | 160 |
| Fully methylated bands | 62 | 60 | 65 | 74 |
| Fully methylated percentage | 53.45% | 42.86% | 57.52% | 46.25% |
| Hemimethylated bands | 54 | 80 | 48 | 86 |
| Hemimethylated percentage | 46.55% | 57.14% | 42.48% | 53.75% |
| MSAP percentage | 33.72% | 42.55% | 32.01% | 43.96% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, U.; Sikdar, A.; Igamberdiev, A.U.; Debnath, S.C. Exploring Genetic and Epigenetic Changes in Lingonberry Using Molecular Markers: Implications for Clonal Propagation. Curr. Issues Mol. Biol. 2023, 45, 6296-6310. https://doi.org/10.3390/cimb45080397
Sharma U, Sikdar A, Igamberdiev AU, Debnath SC. Exploring Genetic and Epigenetic Changes in Lingonberry Using Molecular Markers: Implications for Clonal Propagation. Current Issues in Molecular Biology. 2023; 45(8):6296-6310. https://doi.org/10.3390/cimb45080397
Chicago/Turabian StyleSharma, Umanath, Arindam Sikdar, Abir U. Igamberdiev, and Samir C. Debnath. 2023. "Exploring Genetic and Epigenetic Changes in Lingonberry Using Molecular Markers: Implications for Clonal Propagation" Current Issues in Molecular Biology 45, no. 8: 6296-6310. https://doi.org/10.3390/cimb45080397
APA StyleSharma, U., Sikdar, A., Igamberdiev, A. U., & Debnath, S. C. (2023). Exploring Genetic and Epigenetic Changes in Lingonberry Using Molecular Markers: Implications for Clonal Propagation. Current Issues in Molecular Biology, 45(8), 6296-6310. https://doi.org/10.3390/cimb45080397