
Association between Donor Age and Osteogenic Potential of Human Adipose Stem Cells in Bone Tissue Engineering

 , ,
, ,  and
and
Abstract
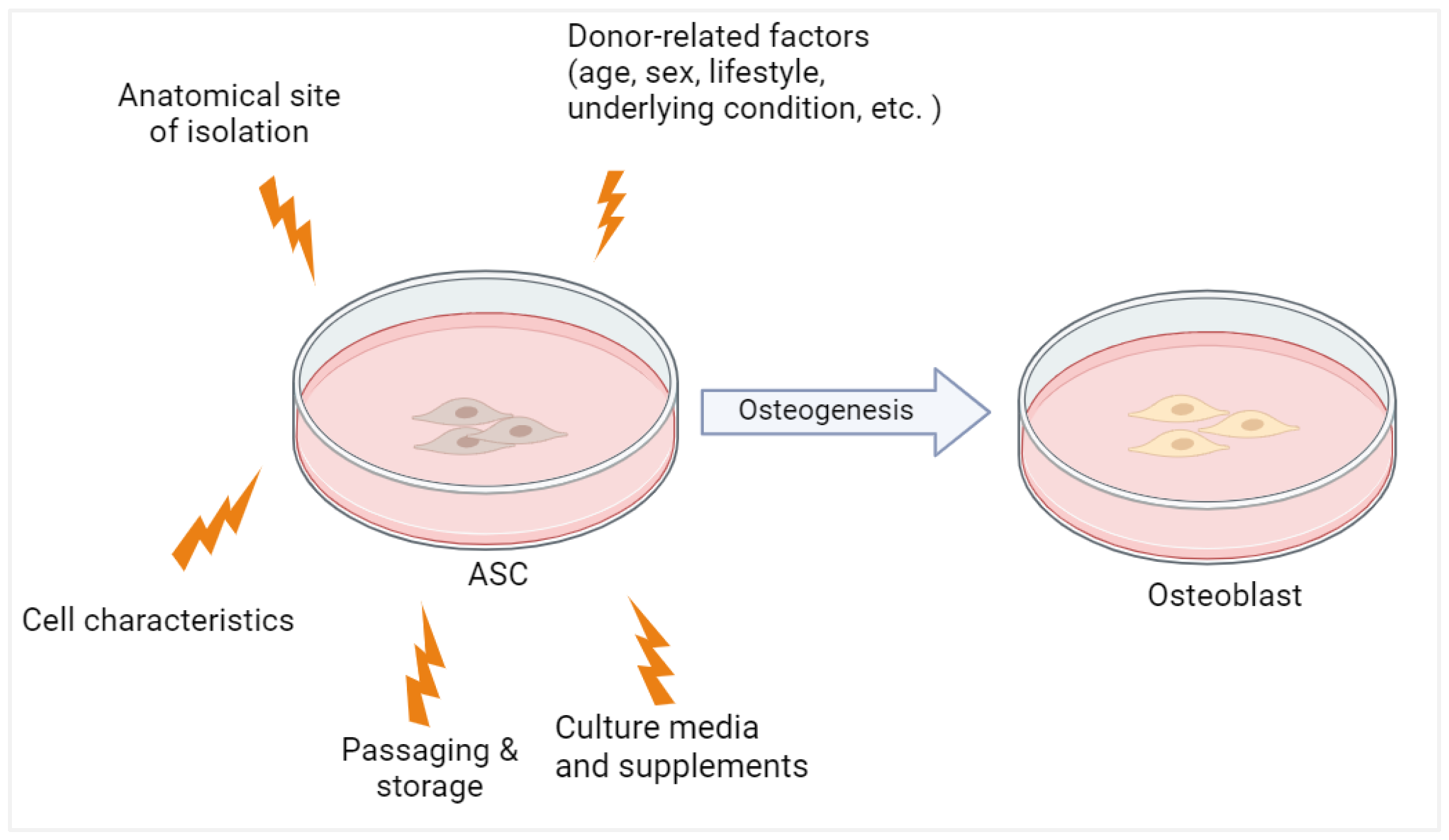
1. Introduction
2. Review of Previous Studies
3. Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pfeiffenberger, M.; Damerau, A.; Lang, A.; Buttgereit, F.; Hoff, P.; Gaber, T. Fracture Healing Research-Shift towards In Vitro Modeling? Biomedicines 2021, 9, 748. [Google Scholar] [CrossRef]
- Mende, W.; Götzl, R.; Kubo, Y.; Pufe, T.; Ruhl, T.; Beier, J.P. The Role of Adipose Stem Cells in Bone Regeneration and Bone Tissue Engineering. Cells 2021, 10, 975. [Google Scholar] [CrossRef]
- Bostrom, M.P. Expression of bone morphogenetic proteins in fracture healing. Clin. Orthop. Relat. Res. 1998, 355, S116–S123. [Google Scholar] [CrossRef]
- Cho, T.J.; Gerstenfeld, L.C.; Einhorn, T.A. Differential temporal expression of members of the transforming growth factor beta superfamily during murine fracture healing. J. Bone Miner. Res. 2002, 17, 513–520. [Google Scholar] [CrossRef]
- Carter, D.R.; Beaupr, G.S.; Giori, N.J.; Helms, J.A. Mechanobiology of skeletal regeneration. Clin. Orthop. Relat. Res. 1998, 355, S41–S55. [Google Scholar] [CrossRef]
- Ghimire, S.; Miramini, S.; Edwards, G.; Rotne, R.; Xu, J.; Ebeling, P.; Zhang, L. The investigation of bone fracture healing under intramembranous and endochondral ossification. Bone Rep. 2021, 14, 100740. [Google Scholar] [CrossRef]
- Roberts, T.T.; Rosenbaum, A.J. Bone grafts, bone substitutes and orthobiologics: The bridge between basic science and clinical advancements in fracture healing. Organogenesis 2012, 8, 114–124. [Google Scholar] [CrossRef]
- Henkel, J.; Woodruff, M.; Epari, D.; Steck, R.; Glatt, V.; Dickinson, I.C.; Choong, P.; Schuetz, M.A.; Hutmacher, D.W. Bone Regeneration Based on Tissue Engineering Conceptions—A 21st Century Perspective. Bone Res. 2013, 1, 216–248. [Google Scholar] [CrossRef]
- Sui, B.; Hu, C.; Zheng, C.; Jin, Y. Microenvironmental Views on Mesenchymal Stem Cell Differentiation in Aging. J. Dent. Res. 2016, 95, 1333–1340. [Google Scholar] [CrossRef]
- Alt, E.U.; Senst, C.; Murthy, S.N.; Slakey, D.P.; Dupin, C.L.; Chaffin, A.E.; Kadowitz, P.J.; Izadpanah, R. Aging alters tissue resident mesenchymal stem cell properties. Stem Cell Res. 2012, 8, 215–225. [Google Scholar] [CrossRef]
- Rosenbaum, A.J.; Grande, D.A.; Dines, J.S. The use of mesenchymal stem cells in tissue engineering: A global assessment. Organogenesis 2008, 4, 23–27. [Google Scholar] [CrossRef]
- Oryan, A.; Kamali, A.; Moshiri, A.; Baghaban Eslaminejad, M. Role of Mesenchymal Stem Cells in Bone Regenerative Medicine: What Is the Evidence? Cells Tissues Organs 2017, 204, 59–83. [Google Scholar] [CrossRef]
- Sethe, S.; Scutt, A.; Stolzing, A. Aging of mesenchymal stem cells. Ageing Res. Rev. 2006, 5, 91–116. [Google Scholar] [CrossRef]
- Bellantuono, I.; Aldahmash, A.; Kassem, M. Aging of marrow stromal (skeletal) stem cells and their contribution to age-related bone loss. Biochim. Biophys. Acta 2009, 1792, 364–370. [Google Scholar] [CrossRef]
- Asumda, F.Z. Age-associated changes in the ecological niche: Implications for mesenchymal stem cell aging. Stem Cell Res. Ther. 2013, 4, 47. [Google Scholar] [CrossRef]
- Baker, N.; Boyette, L.B.; Tuan, R.S. Characterization of bone marrow-derived mesenchymal stem cells in aging. Bone 2015, 70, 37–47. [Google Scholar] [CrossRef]
- Infante, A.; Rodriguez, C.I. Osteogenesis and aging: Lessons from mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 244. [Google Scholar] [CrossRef]
- Prall, W.C.; Saller, M.M.; Scheumaier, A.; Tucholski, T.; Taha, S.; Böcker, W.; Polzer, H. Proliferative and osteogenic differentiation capacity of mesenchymal stromal cells: Influence of harvesting site and donor age. Injury 2018, 49, 1504–1512. [Google Scholar] [CrossRef]
- Majors, A.K.; Boehm, C.A.; Nitto, H.; Midura, R.J.; Muschler, G.F. Characterization of human bone marrow stromal cells with respect to osteoblastic differentiation. J. Orthop. Res. 1997, 15, 546–557. [Google Scholar] [CrossRef]
- Stolzing, A.; Jones, E.; McGonagle, D.; Scutt, A. Age-related changes in human bone marrow-derived mesenchymal stem cells: Consequences for cell therapies. Mech. Ageing Dev. 2008, 129, 163–173. [Google Scholar] [CrossRef]
- Stenderup, K.; Justesen, J.; Clausen, C.; Kassem, M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone 2003, 33, 919–926. [Google Scholar] [CrossRef]
- Zhou, S.; Greenberger, J.S.; Epperly, M.W.; Goff, J.P.; Adler, C.; LeBoff, M.S.; Glowacki, J. Age-related intrinsic changes in human bone-marrow-derived mesenchymal stem cells and their differentiation to osteoblasts. Aging Cell 2008, 7, 335–343. [Google Scholar] [CrossRef]
- Alonso-Goulart, V.; Ferreira, L.B.; Duarte, C.A.; de Lima, I.L.; Ferreira, E.R.; de Oliveira, B.C.; Vargas, L.N.; de Moraes, D.D.; Silva, I.B.B.; Faria, R.d.O.; et al. Mesenchymal stem cells from human adipose tissue and bone repair: A literature review. Biotechnol. Res. Innov. 2018, 2, 74–80. [Google Scholar] [CrossRef]
- Meyer, J.; Salamon, A.; Herzmann, N.; Adam, S.; Kleine, H.-D.; Matthiesen, I.; Ueberreiter, K.; Peters, K. Isolation and differentiation potential of human mesenchymal stem cells from adipose tissue harvested by water jet-assisted liposuction. Aesthet. Surg. J. 2015, 35, 1030–1039. [Google Scholar] [CrossRef]
- Al-Ghadban, S.; Artiles, M.; Bunnell, B.A. Adipose Stem Cells in Regenerative Medicine: Looking Forward. Front. Bioeng. Biotechnol. 2022, 9, 837464. [Google Scholar] [CrossRef]
- Diri, D.; Alasaad, H.; Muhammed, H.; Ibrahim, J. Case report: Adipose-derived mesenchymal stem cells combined with core decompression in the treatment of early-stage avascular necrosis of the femoral head. Int. J. Surg. Case Rep. 2023, 102, 107861. [Google Scholar]
- Li, C.; Mills, Z.; Zheng, Z. Novel cell sources for bone regeneration. MedComm 2021, 2, 145–174. [Google Scholar] [CrossRef]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar]
- Setiawan, A.M.; Kamarudin, T.A.; Abd Ghafar, N. The role of BMP4 in adipose-derived stem cell differentiation: A minireview. Front. Cell Dev. Biol. 2022, 10, 1045103. [Google Scholar] [CrossRef]
- Li, X.; Yao, J.; Wu, L.; Jing, W.; Tang, W.; Lin, Y.; Tian, W.; Liu, L. Osteogenic induction of adipose-derived stromal cells: Not a requirement for bone formation in vivo. Artif. Organs 2010, 34, 46–54. [Google Scholar] [CrossRef]
- Yoshida, Y.; Matsubara, H.; Fang, X.; Hayashi, K.; Nomura, I.; Ugaji, S.; Hamada, T.; Tsuchiya, H. Adipose-derived stem cell sheets accelerate bone healing in rat femoral defects. PLoS ONE 2019, 14, e0214488. [Google Scholar] [CrossRef] [PubMed]
- Dufrane, D. Impact of Age on Human Adipose Stem Cells for Bone Tissue Engineering. Cell Transpl. 2017, 26, 1496–1504. [Google Scholar] [CrossRef]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic review of patient factors affecting adipose stem cell viability and function: Implications for regenerative therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef]
- Holtze, S.; Gorshkova, E.; Braude, S.; Cellerino, A.; Dammann, P.; Hildebrandt, T.B.; Hoeflich, A.; Hoffmann, S.; Koch, P.; Terzibasi Tozzini, E.; et al. Alternative Animal Models of Aging Research. Front. Mol. Biosci. 2021, 8, 660959. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Kohan, E.; Bradley, J.; Hedrick, M.; Benhaim, P.; Zuk, P. The effect of age on osteogenic, adipogenic and proliferative potential of female adipose-derived stem cells. J. Tissue Eng. Regen. Med. 2009, 3, 290–301. [Google Scholar] [CrossRef]
- Girolamo, L.; Lopa, S.; Arrigoni, E.; Sartori, M.; Preis, F.W.B.; Brini, A. Human adipose-derived stem cells isolated from young and elderly women: Their differentiation potential and scaffold interaction during in vitro osteoblastic differentiation. Cytotherapy 2009, 11, 793–803. [Google Scholar] [CrossRef]
- Chen, H.; Lee, M.; Chen, C.; Chuang, S.; Chang, L.; Ho, M.; Hung, S.; Fu, Y.; Wang, Y.; Wang, H.; et al. Proliferation and differentiation potential of human adipose-derived mesenchymal stem cells isolated from elderly patients with osteoporotic fractures. J. Cell. Mol. Med. 2012, 16, 582–592. [Google Scholar] [CrossRef]
- Wu, W.; Niklason, L.; Steinbacher, D.M. The effect of age on human adipose-derived stem cells. Plast. Reconstr. Surg. 2013, 131, 27–37. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef]
- Bodle, J.C.; Teeter, S.D.; Hluck, B.H.; Hardin, J.W.; Bernacki, S.H.; Loboa, E.G. Age-Related Effects on the Potency of Human Adipose-Derived Stem Cells: Creation and Evaluation of Superlots and Implications for Musculoskeletal Tissue Engineering Applications. Tissue Eng. Part C Methods 2014, 20, 972–983. [Google Scholar] [CrossRef]
- Ye, X.; Liao, C.; Liu, G.; Xu, Y.; Tan, J.; Song, Z. Age-Related Changes in the Regenerative Potential of Adipose-Derived Stem Cells Isolated from the Prominent Fat Pads in Human Lower Eyelids. PLoS ONE 2016, 11, e0166590. [Google Scholar] [CrossRef]
- Marędziak, M.; Marycz, K.; Tomaszewski, K.A.; Kornicka, K.; Henry, B.M. The Influence of Aging on the Regenerative Potential of Human Adipose Derived Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 2152435. [Google Scholar] [CrossRef]
- Liu, M.; Lei, H.; Dong, P.; Fu, X.; Yang, Z.; Yang, Y.; Ma, J.; Liu, X.; Cao, Y.; Xiao, R. Adipose-Derived Mesenchymal Stem Cells from the Elderly Exhibit Decreased Migration and Differentiation Abilities with Senescent Properties. Cell Transpl. 2017, 26, 1505–1519. [Google Scholar] [CrossRef]
- Kawagishi-Hotta, M.; Hasegawa, S.; Igarashi, T.; Yamada, T.; Takahashi, M.; Numata, S.; Kobayashi, T.; Iwata, Y.; Arima, M.; Yamamoto, N.; et al. Enhancement of individual differences in proliferation and differentiation potentials of aged human adipose-derived stem cells. Regen. Ther. 2017, 6, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, R.C.; Zhang, J.; Griffith, E.H.; Frank, M.W.; Starly, B.; Loboa, E.G. Electrical Cell-Substrate Impedance Spectroscopy Can Monitor Age-Grouped Human Adipose Stem Cell Variability During Osteogenic Differentiation. Stem Cells Transl. Med. 2017, 6, 502–511. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Z.; Zhao, Y.; Zhang, L.; Xu, L.; Cao, L.; He, W. The Effect of Age on the Regenerative Potential of Human Eyelid Adipose-Derived Stem Cells. Stem Cells Int. 2018, 2018, 5654917. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, C.D.; Barisón, M.J.; Robert, A.W.; Kuligovski, C.; Aguiar, A.M.; Dallagiovanna, B. Influence of donor age on the differentiation and division capacity of human adipose-derived stem cells. World J. Stem Cells 2020, 12, 1640–1651. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Park, G.; Hong, H.S. Age affects the paracrine activity and differentiation potential of human adipose-derived stem cells. Mol. Med. Rep. 2021, 23, 160. [Google Scholar] [CrossRef] [PubMed]
- Kornicka, K.; Marycz, K.; Tomaszewski, K.A.; Marędziak, M.; Śmieszek, A. The Effect of Age on Osteogenic and Adipogenic Differentiation Potential of Human Adipose Derived Stromal Stem Cells (hASCs) and the Impact of Stress Factors in the Course of the Differentiation Process. Oxidat. Med. Cell. Longev. 2015, 2015, 309169. [Google Scholar] [CrossRef]
- Li, K.; Shi, G.; Lei, X.; Huang, Y.; Li, X.; Bai, L.; Qin, C. Age-related alteration in characteristics, function, and transcription features of ADSCs. Stem Cell Res. Ther. 2021, 12, 473. [Google Scholar] [CrossRef]
- Doshida, Y.; Sano, H.; Iwabuchi, S.; Aigaki, T.; Yoshida, M.; Hashimoto, S.; Ishigami, A. Age-associated changes in the transcriptomes of non-cultured adipose-derived stem cells from young and old mice assessed via single-cell transcriptome analysis. PLoS ONE 2020, 15, e0242171. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.-Y.; Nacamuli, R.P.; Salim, A.; Longaker, M.T. The osteogenic potential of adipose-derived mesenchymal cells is maintained with aging. Plast. Reconstr. Surg. 2005, 116, 1686–1696. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, A.S. Demographic and Cultural Differences in the Acceptance and Pursuit of Cosmetic Surgery: A Systematic Literature Review. Plast. Reconstr. Surg. Glob. Open 2021, 9, e3501. [Google Scholar] [CrossRef] [PubMed]
- Schipper, B.M.; Marra, K.G.; Zhang, W.B.; Donnenberg, A.D.; Rubin, J.P. Regional anatomic and age effects on cell function of human adipose-derived stem cells. Ann. Plast. Surg. 2008, 60, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Piao, J.; Park, G.; Yoo, K.S.; Hong, H.S. Osteoporotic Conditions Influence the Activity of Adipose-Derived Stem Cells. Tissue Eng. Regen. Med. 2020, 17, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Prieto González, E.A. Heterogeneity in Adipose Stem Cells. Adv. Exp. Med. Biol. 2019, 1123, 119–150. [Google Scholar] [PubMed]
- Frazier, T.P.; Gimble, J.M.; Devay, J.W.; Tucker, H.A.; Chiu, E.S.; Rowan, B.G. Body mass index affects proliferation and osteogenic differentiation of human subcutaneous adipose tissue-derived stem cells. BMC Cell Biol. 2013, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Prunet-Marcassus, B.; Cousin, B.; Caton, D.; André, M.; Pénicaud, L.; Casteilla, L. From heterogeneity to plasticity in adipose tissues: Site-specific differences. Exp. Cell Res. 2006, 312, 727–736. [Google Scholar] [CrossRef]
- Requicha, J.F.; Viegas, C.A.; Albuquerque, C.M.; Azevedo, J.M.; Reis, R.L.; Gomes, M.E. Effect of Anatomical Origin and Cell Passage Number on the Stemness and Osteogenic Differentiation Potential of Canine Adipose-Derived Stem Cells. Stem Cell Rev. Rep. 2012, 8, 1211–1222. [Google Scholar] [CrossRef]
- Bogdanova, A.; Berzins, U.; Nikulshin, S.; Skrastina, D.; Ezerta, A.; Legzdina, D.; Kozlovska, T. Characterization of human adipose-derived stem cells cultured in autologous serum after subsequent passaging and long term cryopreservation. J. Stem Cells 2014, 9, 135–148. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Year | Aim | Study Design | Methods | Results | Conclusions | Other Variables | Ref. |
|---|---|---|---|---|---|---|---|
| 2009 | Effect of donor age on differentiation potential of ASCs. | ASCs from females aged 20–58 years; N = 27, n = 9, 7, 11. | ALP activity after 7 days Ca2+ deposition by von Kossa after 4 weeks | ↓ ALP Activity with age ↓ Ca2+ deposition with age | Osteogenic differentiation decreases with age | SC: female, non-obese AO: u.s (liposuction) Passage: 1 OI day: up to 28 days | [35] |
| 2009 | In vitro differentiation potential of ASCs from young and elderly females. | Young (<35) and older (>45) females N = 26. | ALP activity Assay after 14 days Ca2+ deposition by Alizarin Red assay after 21 days | ↓ ALP activity with age No significant difference in Ca2+ deposition | Donor age mildly affects the potential of ASCs for osteogenic differentiation in vitro | SC: non-obese female, BMI < 30 AO: subcutaneous (lipo-asp.) Passage: 1 OI day: 21 days | [36] |
| 2012 | Age-associated changes in molecular characteristics of ASCs. | ASCs from healthy young (<20), middle-aged (30–40), and elderly (>50) donors. N = 40; n = 15, 17, 8. | Ca2+ deposition by Alizarin Red S ALP activity assay Osteogenic gene (BMP-6, COL2A, COL10A) expression by RT-PCR | ↓ Ca2+ deposition ↓ ALP activity ↓ Expression of osteogenic genes | Aging processes significantly attenuate the osteogenic differentiation potential of ASCs | SC: healthy male/female, BMI < 29 AO: abdominal Passage: u.s OI day: u.s | [10] |
| 2012 | Effect of age on ASCs and bmMSCs from elderly patients with osteoporosis. | ASCs from young (<36) and elderly (>67) individuals N = 22, n = 14, 8. | Ca2+ deposition by Alizarin Red S | No significant difference in Ca2+ deposition | The osteogenic differentiation of ASCs is not impacted by age | SC: male/female, osteoporotic, BMI < 26 AO: gluteal subcutaneous Passage: 5 OI day: up to 14 days | [37] |
| Osteogenic genes (OCA, BMP2, RUNX2, and ALP) by RT-PCR | No significant difference in gene expression. | ||||||
| 2012 | Effect of aging on senescence, osteogenic factors, and osteogenesis of ASCs. | ASCs from infants (<1), adults (20–54), and elderly individuals (>55), N = 13; n = 4, 6, 3. | Senescence (TL) by RT-PCR RUNX2, osteocalcin by RT-PCR ALP activity assay Ca2+ deposition by Alizarin Red S | ↑ Senescence with age ↓ Osteogenic gene expression compared to infant ↓ ALP activity and Ca2+ deposition compared to infant. | Biological properties are conserved during the adult to the elderly period (but not compared to infants) | SC: male/female. AO: abdominal (liposuction) Passage: 1 OI day: up to 21 days | [38] |
| 2014 | Impact of age on the quality of human adipose tissue-derived MSCs. | ASCs of young (<30), adult (35–50), and elderly (>60) donors, N = 29; n = 10, 8, 11. | Ca2+ deposition by von Kossa staining Senescence by β-galactosidase Staining Osteogenic genes (osteocalcin and ALP) by RT-PCR | ↓ Ca2+ deposition with age ↑ Senescence with age ↓ Expression of osteocalcin and ALP with age. | Age negatively impacts stem cell osteogenic differentiation | SC: male/female AO: (liposuction) Passage: 2–3 OI day: 21 days | [39] |
| 2014 | Effect of age on osteogenesis of female ASCs: superlot approach. | ASCs from female patients (24–81), superlot biobanking. N = 14; n = 5, 4, 5. | Ca2+ deposition by Alizarin Red S | ↑ Ca2+ deposition with older (postmenopausal) female. | Existence of a high degree of donor-to-donor variations which is independent of age | SC: female AO: u.s Passage: 1 OI day: 14 days | [40] |
| 2016 | Effects of donor age on the biological properties of human OASC. | OASCs from young (20–38, normal) and old donors (50–67, fat pad in lower eyelid) N = 20; n = 10, 10. | Ca2+ deposition by Alizarin Red S Ca2+ deposition by Von Kossa staining | ↓ Ca2+ deposition with age | The benefit of autologous OASCs from elderly patients for osteogenic therapeutic purposes may be limited | SC: female, non-obese AO: lower eyelid fat pad Passage: 3 OI day: 14 days | [41] |
| 2016 | Effect of age on the osteogenic potential of ASCs. | ASCs from different age groups: >20 y, >50 y, >60, >70 N = 32; n = 8. | Ca2+ deposition by Alizarin Red ALP activity assay Osteogenic markers (OPN, Col-I, OCL, and BMP-2) by PCR | ↓ Ca2+ deposition with age No significant difference in ALP activity ↓ Expression of osteogenic markers with age | Age negatively influences the osteogenic potential of ASCs | SC: healthy male/female AO: subcutaneous Passage: 1 OI day: 21 days | [42] |
| 2017 | Systematical analysis of the effects of age on the quantity and quality of ASCs. | ASCs were isolated from children (6–12), young individuals (22–27), adults (60–73), and the elderly, N = 24; n = 10, 8, 6. | Cellular senescence assay Ca2+ deposition by Alizarin Red S Osteogenic genes (RUNX2, ALP, OCN, and OPN) by RT-PCR | ASCs from elderly donors exhibit senescent properties. ASCs from aged patients exhibit impaired osteogenic potential | While ASCs from different age populations are phenotypically similar, they present major differences at the functional level | SC: male/female BMI < 22 AO: chest subcutaneous Passage: 3 OI day: up to 21 days | [43] |
| 2017 | Effect of donor age on differentiation potential of ASCs. | ASCs of 260 donors (ages 5–97 years) N = 260. | Ca2+ deposition by Alizarin Red S | The osteogenic potential (marked by Ca2+ deposition) of ASCs does not correlate with donor age | The chondrogenic and osteogenic potential of ASCs were not affected by age | SC: male/female, median BMI = 22.7 AO: subcutaneous Passage: 5 OI day: 21 days | [44] |
| 2017 | Cell–substrate impedance spectroscopy (ECIS) to track complex bioimpedance pattern of ASC osteogenesis. | ASC superlot from young (24–36), middle-aged (48–55), and elderly (60–81) adults. | ECIS measurement throughout the osteogenic differentiation phases | ASCs from younger donors require a longer time to differentiate than ASCs from older donors. | Donor age may temporally control the onset of osteogenesis | SC: female AO: u.s (liposuction) Passage: u.s CS: u.s OI day: u.s | [45] |
| 2018 | Effect of donor age on the regenerative potential of HEASCs. | HEASCs from <20 y, >20 y, <45 y, >55 y N = 13; n = 4, 5, 4. | Ca2+ deposition by Alizarin Red S RUNX2 by RT-PCR | ↓ Ca2+ deposition with age No difference in gene expression. | Donor age has a negative influence on the osteogenic differentiation of HEASCs | SC: healthy donor AO: eyelid Passage: 5 OI day: 21 days | [46] |
| 2020 | Differentiation potential of ASCs isolated from the lipoaspirates of elderly and young donors. | ASCs from young (<34) and old (>54) female donors, N = 18; n = 9, 9. | Cell mineralization assay RUNX2 by RT-PCR | No significant difference No significant difference | Age does not significantly impact the osteogenesis of ASCs | SC: female, BMI < 30; AO: u.s (liposuction) Passage: 4–7 OI day: up to 28 days | [47] |
| 2021 | Association between age and ASC differentiation potential. | ASCs from young (<30) and elderly (>70), N = 8. | Ca2+ deposition by Alizarin Red S BMP-2 by ELISA BMP-2 receptor by WB | ↓ Ca2+ deposition with age No significant difference ↓ BMP-2 with age | Age may affect the cellular function and differentiation of ASCs | SC: healthy male/female, AO: u.s Passage: 3–5 OI day: 20 days | [48] |
| Method | Positive Markers | Negative Markers | References |
|---|---|---|---|
| Flow Cytometry | CD44, CD90, CD105, CD146 | CD3, CD4, CD11b, CD34, CD45 | [10] |
| Flow Cytometry | CD44, CD73, CD90,CD105 | - | [38] |
| Flow Cytometry | CD44, CD73, CD90, CD105 | CD3, CD14, CD19, CD34, CD45 | [39] |
| Flow Cytometry | CD73, CD90, CD105 | CD14, CD19, CD34, CD45 | [41] |
| Flow Cytometry | CD44, CD73, CD90, CD105 | CD34, CD45 | [42] |
| Flow Cytometry | CD44, CD73, CD90, CD105 | CD34, CD11b, CD19, CD45, HLA-DR | [43] |
| RT-qPCR | CD44, CD73, CD90, CD105, CD271, NANOG | - | [44] |
| Flow Cytometry | CD44, CD73, CD90, CD105 | CD31, CD34, CD45 | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sattar, M.A.; Lingens, L.F.; Guillaume, V.G.J.; Goetzl, R.; Beier, J.P.; Ruhl, T. Association between Donor Age and Osteogenic Potential of Human Adipose Stem Cells in Bone Tissue Engineering. Curr. Issues Mol. Biol. 2024, 46, 1424-1436. https://doi.org/10.3390/cimb46020092
Sattar MA, Lingens LF, Guillaume VGJ, Goetzl R, Beier JP, Ruhl T. Association between Donor Age and Osteogenic Potential of Human Adipose Stem Cells in Bone Tissue Engineering. Current Issues in Molecular Biology. 2024; 46(2):1424-1436. https://doi.org/10.3390/cimb46020092
Chicago/Turabian StyleSattar, Md Abdus, Lara F. Lingens, Vincent G. J. Guillaume, Rebekka Goetzl, Justus P. Beier, and Tim Ruhl. 2024. "Association between Donor Age and Osteogenic Potential of Human Adipose Stem Cells in Bone Tissue Engineering" Current Issues in Molecular Biology 46, no. 2: 1424-1436. https://doi.org/10.3390/cimb46020092
APA StyleSattar, M. A., Lingens, L. F., Guillaume, V. G. J., Goetzl, R., Beier, J. P., & Ruhl, T. (2024). Association between Donor Age and Osteogenic Potential of Human Adipose Stem Cells in Bone Tissue Engineering. Current Issues in Molecular Biology, 46(2), 1424-1436. https://doi.org/10.3390/cimb46020092