
Isolation of a Virulent Clostridium perfringens Strain from Elaphurus davidianus and Characterization by Whole-Genome Sequence Analysis

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Bacteria and Resuscitating Bacteria
2.2. Assay of Phospholipase C Clostridium (PLC) Activity
2.3. Hemolytic Activity Assay
2.4. Mouse LD50 Test
2.5. Whole-Genome Sequencing Analysis of the Most Virulent Isolates
2.6. Comparative Genomic Analysis of the Most Virulent Strains
2.7. Statistical Analysis
3. Results
3.1. Phospholipase C Activity Assay
3.2. Determination of Hemolytic Activity
3.3. Pathogenicity Test in Mice
3.4. Whole-Genome Sequencing Analysis of Virulent Strain CX1-4
3.5. Generic Database Annotation Results
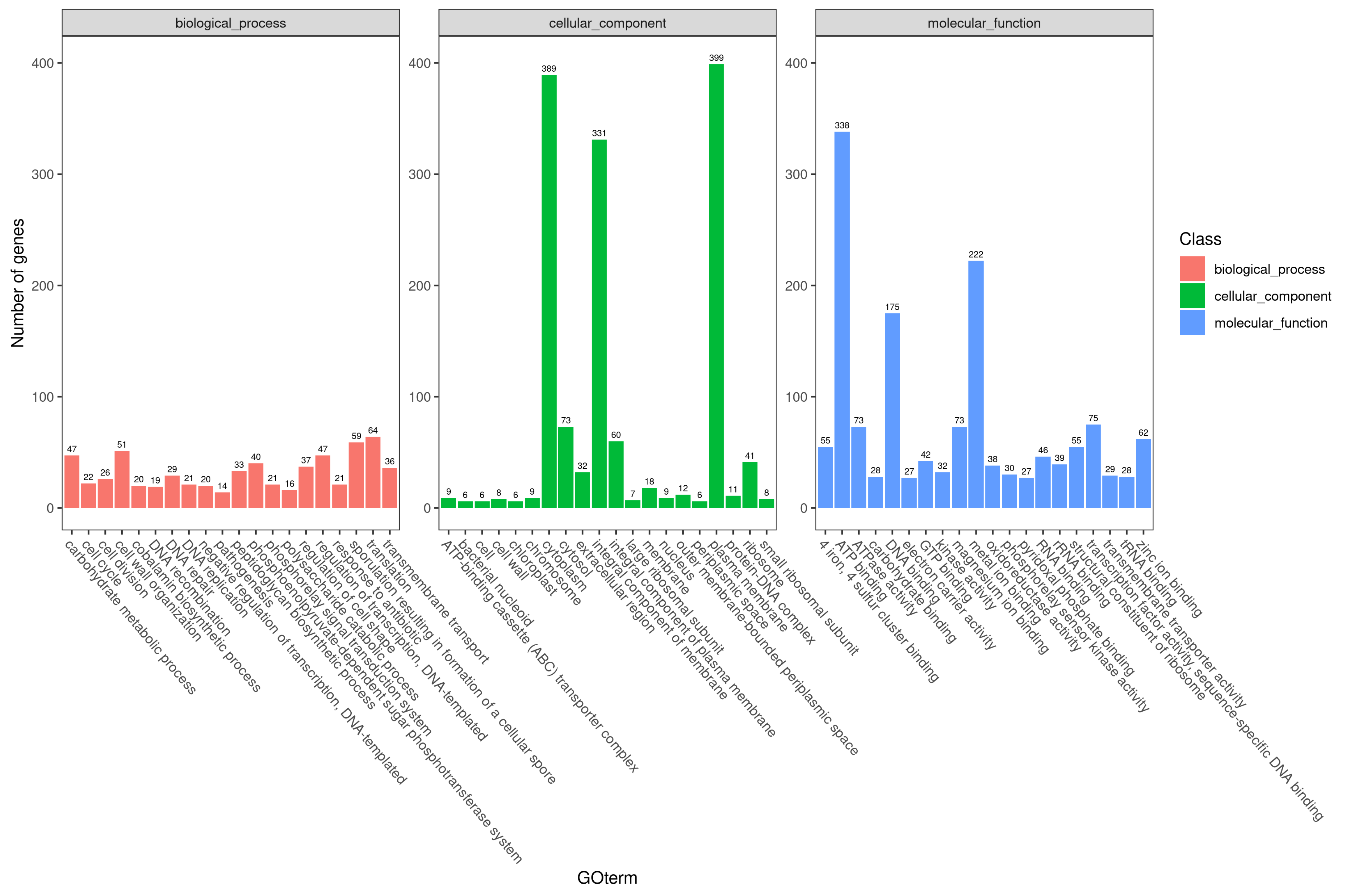
3.5.1. Gene Ontology Classification Results
3.5.2. KEGG Pathway Classification
3.5.3. COG Classification Results
3.5.4. VFDB Annotation Results
3.6. Results of Comparative Genomic Analysis of Virulent CX1-4 Strain
3.6.1. Comparison of Basic Genomic Features
3.6.2. Genomic ANI Analysis and Analysis of Genomic Collinearity
3.6.3. Comparison of Virulence Factors
4. Discussion
4.1. Selection of the Most Virulent Strains
4.2. Whole-Genome Sequencing Analysis of Virulent Strain CX1-4
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANI | Average nucleotide identity |
| CDS | Coding DNA sequences |
| COG | Cluster of orthologous group |
| CPE | C. perfringens enterotoxin |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LD50 | Median lethal dose |
| NCBI | National Center for Biotechnology Information |
| NetB | Necrotising B-like |
| OD | Optical density |
| PLC | Phospholipase C |
| VFDB | Virulence Factor Database |
References
- Zhou, Y.; Elk, G. Return and come back, ever living and thriving. J. Hunan 2024. [Google Scholar] [CrossRef]
- Zhang, F.X.; Meng, Q.H.; Jiang, Z.G.; Cheng, Z.B.; Song, Y.; Bai, J.D.; Ding, Y.H.; Wen, H.J. The vicissitudes of the elk’s fate and prospects for future protection. Acta Ecol. Sin. 2022, 42, 1–10. [Google Scholar]
- Ohtani, K.; Shimizu, T. Regulation of toxin production in Clostridium perfringens. Toxins 2016, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- McClane, B.A.; Robertson, S.L.; Li, J. Clostridium perfringens. In Food Microbiology: Fundamentals and Frontiers, 4th ed.; ASM Press: Washington, DC, USA, 2012; pp. 465–489. [Google Scholar]
- Mehdizadeh Gohari, I.; Navarro, M.A.; Li, J.; Shrestha, A.; Uzal, F.; McClane, A.B. Pathogenicity and virulence of Clostridium perfringens. Virulence 2021, 12, 723–753. [Google Scholar]
- Dumas, G.; Tankovic, J.; Bigé, N.; Maury, E. Clostridium perfringens related spleen gangrene. Intensive Care Med. 2017, 43, 1730–1731. [Google Scholar] [CrossRef] [PubMed]
- Freedman, J.C.; Shrestha, A.; McClane, B.A. Clostridium perfringens enterotoxin: Action, genetics, and translational applications. Toxins 2016, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Revitt-Mills, S.A.; I Rood, J.; Adams, V. Clostridium perfringens extracellular toxins and enzymes: 20 and counting. Microbiol. Aust. 2015, 36, 114–117. [Google Scholar] [CrossRef]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.A.; McClane, B.A.; Uzal, F.A. Mechanisms of action and cell death associated with Clostridium perfringens toxins. Toxins 2018, 10, 212. [Google Scholar] [CrossRef]
- Moustafa, S.; Zakaria, I.; Moustafa, A.; AboSakaya, R.; Selim, A. Bacteriological and serological investigation of Clostridium perfringens in lambs. Sci. Rep. 2022, 12, 19715. [Google Scholar] [CrossRef]
- Kiu, R.; Sim, K.; Shaw, A.; Cornwell, E.; Pickard, D.; Kroll, J.S.; Hall, L.J. Genomic analysis of Clostridium perfringens BEC/CPILE-positive, toxinotype D and E strains isolated from healthy children. Toxins 2019, 11, 543. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Adams, V.; Bannam, T.L.; Miyamoto, K.; Garcia, J.P.; Uzal, F.A.; Rood, J.I.; McClane, B.A. Toxin plasmids of Clostridium perfringens. Microbiol. Mol. Biol. Rev. 2013, 77, 208–233. [Google Scholar] [CrossRef] [PubMed]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Shimizu, T. Regulation of toxin gene expression in Clostridium perfringens. Res. Microbiol. 2015, 166, 280–289. [Google Scholar] [CrossRef]
- Wash, P.; Yasmin, H.; Ullah, H.; Haider, W.; Khan, N.; Ahmad, A.; Mumtaz, S.; Hassan, M.N. Deciphering the genetics of antagonism and antimicrobial resistance in Bacillus velezensis HU-91 by whole genome analysis. J. King Saud. Univ. -Sci. 2023, 35, 102954. [Google Scholar] [CrossRef]
- Xing, J.; Ji, X.; Sun, Y.; Zhu, L.; Jiang, Q.; Guo, X.; Liu, J. Preparation and immunological characterization of an inactivated canine Clostridium perfringens type A vaccine. Lett. Appl. Microbiol. 2019, 69, 385–390. [Google Scholar] [CrossRef]
- Karaseva, O.; Ozhegov, G.; Khusnutdinova, D.; Siniagina, M.; Anisimova, E.; Akhatova, F.; Fakhrullin, R.; Yarullina, D. Whole Genome Sequencing of the Novel Probiotic Strain Lactiplantibacillus plantarum FCa3L. Microorganisms 2023, 11, 1234. [Google Scholar] [CrossRef]
- Boucard, A.-S.; Florent, I.; Polack, B.; Langella, P.; Bermúdez-Humarán, L.G. Genome sequence and assessment of safety and potential probiotic traits of lactobacillus johnsonii CNCM I-4884. Microorganisms 2022, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Tregaskis, P.L.; Devaney, R.; Smyth, V.J. The first whole genome sequence and characterisation of avian nephritis virus genotype 3. Viruses 2021, 13, 235. [Google Scholar] [CrossRef]
- Kiu, R.; Brown, J.; Bedwell, H.; Leclaire, C.; Caim, S.; Pickard, D.; Dougan, G.; Dixon, R.A.; Hall, L.J. Genomic analysis on broiler-associated Clostridium perfringens strains and exploratory caecal microbiome investigation reveals key factors linked to poultry necrotic enteritis. Anim. Microbiome 2019, 1, 12. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Y.; Yang, D.; Zhang, S.; Sun, Z.; Wang, Y.; Wang, S.; Wu, C. Prevalence and antimicrobial susceptibility of Clostridium perfringens in chickens and pigs from Beijing and Shanxi, China. Vet. Microbiol. 2021, 252, 108932. [Google Scholar] [CrossRef] [PubMed]
- Issimov, A.; Baibatyrov, T.; Tayeva, A.; Kenenbay, S.; Abzhanova, S.; Shambulova, G.; Kuzembayeva, G.; Kozhakhiyeva, M.; Brel-Kisseleva, I.; Safronova, O.; et al. Prevalence of Clostridium perfringens and Detection of Its Toxins in Meat Products in Selected Areas of West Kazakhstan. Agriculture 2022, 12, 1357. [Google Scholar] [CrossRef]
- Milton, A.A.P.; Agarwal, R.K.; Priya, G.B.; Saminathan, M.; Aravind, M.; Reddy, A.; Athira, C.; Ramees, T.; Sharma, A.K.; Kumar, A. Prevalence and molecular typing of Clostridium perfringens in captive wildlife in India. Anaerobe 2017, 44, 55–57. [Google Scholar] [CrossRef]
- Hoffmann, S.; Batz, M.B.; Morris, J.G., Jr. Annual cost of illness and quality-adjusted life year losses in the United States due to 14 foodborne pathogens. J. Food Prot. 2012, 75, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.M.; Callan, R.J.; Van Metre, D.C. Clostridial abomasitis and enteritis in ruminants. Vet. Clin. Food Anim. Pract. 2018, 34, 155–184. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.; Smithee, L.; McClane, B.; Distefano, R.F.; Uzal, F.; Songer, J.G.; Mallonee, S.; Crutcher, J.M. Fatal necrotizing colitis following a foodborne outbreak of enterotoxigenic Clostridium perfringens type A infection. Clin. Infect. Dis. 2005, 40, e78–e83. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Freedman, J.C.; Shrestha, A.; Theoret, J.R.; Garcia, J.; Awad, M.M.; Adams, V.; Moore, R.J.; I Rood, J.; A McClane, B. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Future Microbiol. 2014, 9, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Vidal, J.E.; McClane, B.A.; Gurjar, A.A. Clostridium perfringens toxins involved in mammalian veterinary diseases. Open Toxinology J. 2010, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Babajimopoulus, M.; Mikolajcik, E.M. Purification and activity of Clostridium perfringens alpha toxin. J. Food Prot. 1977, 40, 831–834. [Google Scholar] [CrossRef]
- Urbina, P.; Collado, M.I.; Alonso, A.; Goñi, F.M.; Flores-Díaz, M.; Alape-Girón, A.; Ruysschaert, J.-M.; Lensink, M.F. Unexpected wide substrate specificity of C. perfringens α-toxin phospholipase C. Biochim. Biophys. Acta 2011, 1808, 2618–2627. [Google Scholar] [CrossRef]
- Oda, M.; Terao, Y.; Sakurai, J.; Nagahama, M. Membrane-Binding Mechanism of Clostridium perfringens Alpha-Toxin. Toxins 2015, 7, 5268–5275. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.; Zhao, H.; Ye, C.; Fu, S.; Liu, Y.; Liu, T.; Qiu, Y. Baicalin-aluminum alleviates necrotic enteritis in broiler chickens by inhibiting virulence factors expression of Clostridium perfringens. Front. Cell Infect. Microbiol. 2023, 13, 1243819. [Google Scholar] [CrossRef] [PubMed]
- Hacker, J.; Blum-Oehler, G.; Mühldorfer, I.; Tschäpe, H. Pathogenicity islands of virulent bacteria: Structure, function and impact on microbial evolution. Mol. Microbiol. 1997, 23, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Hsiao, W.W.L.; Brinkman, F.S.L. Detecting genomic islands using bioinformatics approaches. Nat. Rev. Microbiol. 2010, 8, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Hudson, C.M.; Bent, Z.W.; Meagher, R.J.; Williams, K.P. Resistance determinants and mobile genetic elements of an NDM-1-encoding Klebsiella pneumoniae strain. PLoS ONE 2014, 9, e99209. [Google Scholar] [CrossRef] [PubMed]
- Blum, G.; Ott, M.; Lischewski, A.; Ritter, A.; Imrich, H.; Tschäpe, H.; Hacker, J. Excision of large DNA regions termed pathogenicity islands from tRNA-specific loci in the chromosome of an Escherichia coli wild-type pathogen. Infect. Immun. 1994, 62, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Seth-Smith, H.M.B.; Fookes, M.C.; Okoro, C.K.; Baker, S.; Harris, S.R.; Scott, P.; Pickard, D.; Quail, M.A.; Churcher, C.; Sanders, M.; et al. Structure, diversity, and mobility of the Salmonella pathogenicity island 7 family of integrative and conjugative elements within Enterobacteriaceae. J. Bacteriol. 2012, 194, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Cain, J.A.; Montibus, B.; Oakey, R.J. Intragenic CpG islands and their impact on gene regulation. Front. Cell Dev. Biol. 2022, 10, 832348. [Google Scholar] [CrossRef]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef]
- Dhavan, G.M.; Crothers, D.M.; Chance, M.R.; Brenowitz, M. Concerted binding and bending of DNA by Escherichia coli integration host factor. J. Mol. Biol. 2002, 315, 1027–1037. [Google Scholar] [CrossRef]
- Perez-Howard, G.M.; Weil, P.A.; Beechem, J.M. Yeast TATA binding protein interaction with DNA: Fluorescence determination of oligomeric state, equilibrium binding, on-rate, and dissociation kinetics. Biochemistry 1995, 34, 8005–8017. [Google Scholar] [CrossRef]
- Vargas, R., Jr.; Luna, M.J.; Freschi, L.; Marin, M.; Froom, R.; Murphy, K.C.; Campbell, E.A.; Ioerger, T.R.; Sassetti, C.M.; Farhat, M.R. Phase variation as a major mechanism of adaptation in Mycobacterium tuberculosis complex. Proc. Natl. Acad. Sci. USA 2023, 120, e2301394120. [Google Scholar] [CrossRef] [PubMed]
- Seth, W.C.; Geoffrey, J.F.; Marcel, E.D. Overcoming challenges and dogmas to understand the functions of pseudogenes. Nat. Rev. Genet. 2020, 21, 191–201. [Google Scholar]
- Nakamura-García, A.K.; Espinal-Enríquez, J. Pseudogenes in Cancer: State of the Art. Cancers 2023, 15, 4024. [Google Scholar] [CrossRef] [PubMed]
- Di Sanzo, M.; Quaresima, B.; Biamonte, F.; Palmieri, C.; Faniello, M.C. FTH1 Pseudogenes in Cancer and Cell Metabolism. Cells. 2020, 9, 2554. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.I.; Jabbar, A.; Imran, M.; Ahmed, S.; Majeed, T.; Tolba, M.M.; Mahnoor, M.; Nasser, H.A.; Khan, M.A.; Simirgiotis, M.J. Behind the Scene: Surprises and Snags of Pseudogenes. Crit. Rev. Eukaryot. Gene Expr. 2021, 31, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, T.F.; Patrushev, L.I. Pseudogenes as Functionally Significant Elements of the Genome. Biochemistry 2018, 83, 1332–1349. [Google Scholar] [CrossRef]
- Chen, X.; Wan, L.; Wang, W.; Xi, W.J.; Yang, A.G.; Wang, T. Re-recognition of pseudogenes: From molecular to clinical applications. Theranostics 2020, 10, 1479–1499. [Google Scholar] [CrossRef] [PubMed]
- Ten Berk de Boer, E.; Bilgrav Saether, K.; Eisfeldt, J. Discovery of non-reference processed pseudogenes in the Swedish population. Front. Genet. 2023, 14, 1176626. [Google Scholar] [CrossRef]
- Emadi-Baygi, M.; Sedighi, R.; Nourbakhsh, N.; Nikpour, P. Pseudogenes in gastric cancer pathogenesis: A review article. Brief. Funct. Genom. 2017, 16, 348–360. [Google Scholar] [CrossRef]
- Paredes-Sabja, D.; Sarker, N.; Sarker, M.R. Clostridium perfringens tpeL is expressed during sporulation. Microb. Pathog. 2011, 51, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Edwards, A.N.; Sarker, M.R.; Paredes-Sabja, D. Sporulation and germination in clostridial pathogens. Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Alnoman, M.; Udompijitkul, P.; Banawas, S.; Sarker, M.R. Bicarbonate and amino acids are co-germinants for spores of Clostridium perfringens type A isolates carrying plasmid-borne enterotoxin gene. Food Microbiol. 2018, 69, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Sabja, D.; Raju, D.; Torres, J.A.; Sarker, M.R. Role of small, acid-soluble spore proteins in the resistance of Clostridium perfringens spores to chemicals. Int. J. Food Microbiol. 2008, 122, 333–335. [Google Scholar] [CrossRef]
- Paredes-Sabja, D.; Sarker, N.; Setlow, B.; Setlow, P.; Sarker, M.R. Roles of DacB and Spm proteins in Clostridium perfringens spore resistance to moist heat, chemicals, and UV radiation. Appl. Environ. Microbiol. 2008, 74, 3730–3738. [Google Scholar] [CrossRef]
- Paredes-Sabja, D.; Setlow, B.; Setlow, P.; Sarker, M.R. Characterization of Clostridium perfringens spores that lack SpoVA proteins and dipicolinic acid. J. Bacteriol. 2008, 190, 4648–4659. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Sabja, D.; Torres, J.A.; Setlow, P.; Sarker, M.R. Clostridium perfringens spore germination: Characterization of germinants and their receptors. J. Bacteriol. 2008, 190, 1190–1201. [Google Scholar] [CrossRef] [PubMed]
- Sakanoue, H.; Nakano, T.; Sano, K.; Yasugi, M.; Monma, C.; Miyake, M. Adherence of Clostridium perfringens spores to human intestinal epithelial Caco-2 cells. FEMS Microbiol. Lett. 2018, 365, fny016. [Google Scholar] [CrossRef]
- Fourie, J.C.J.; Bezuidenhout, C.C.; Sanko, T.J.; Mienie, C.; Adeleke, R. Inside environmental Clostridium perfringens genomes: Antibiotic resistance genes, virulence factors and genomic features. J. Water Health 2020, 18, 477–493. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.J. A genomic perspective on protein families. Science 1997, 278, 631–637. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Z.; Wang, Y. Contents, construction methods, data resources, and functions comparative analysis of bacteria databases. Int. J. Biol. Sci. 2020, 16, 838. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef]
- Yang, J.; Chen, L.; Sun, L.; Yu, J.; Jin, Q. VFDB 2008 release: An enhanced web-based resource for comparative pathogenomics. Nucleic Acids Res. 2008, 36, D539–D542. [Google Scholar] [CrossRef]
- Chen, L.; Xiong, Z.; Sun, L.; Yang, J.; Jin, Q. VFDB 2012 update: Toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Res. 2012, 40, D641–D645. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, H.; Hazarika, R.; Tamuly, S.; Deka, P.; Manoharan, S.; Sharma, R.K. Evaluation of different antigenic preparations against necrotic enteritis in broiler birds using a novel Clostridium perfringens type G strain. Anaer-Obe 2021, 70, 102377. [Google Scholar] [CrossRef]
- Navarro, M.A.; Li, J.; Beingesser, J.; McClane, B.A.; Uzal, F.A. NanI sialidase enhances the action of Clostridium perfringens enterotoxin in the presence ofmucus. mSphere 2021, 6, e0084821. [Google Scholar] [CrossRef]
- Zeng, X.; Liu, B.; Zhou, J.; Dai, Y.; Han, C.; Wang, L.; Wu, Y.; Zhang, J. Complete genomic sequence and analysis of β2 toxin gene mapping of Clostridium perfringens JXJA17 isolated from piglets in China. Sci. Rep. 2021, 11, 475. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.T.; Hartigan, J.A. Reconstruction of evolutionary trees from pairwise distributions on current species. In Computing Science and Statistics: Proceedings of the 23rd Symposium on the Interface; Interface Foundation: Fairfax Station, VA, USA, 1991; Volume 254, p. 257. [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Read, D.A.; Slippers, B.; Steenkamp, E. Genomic characterization of a novel potyvirus infecting Barleria repens in South Africa. Arch. Virol. 2022, 168, 10. [Google Scholar] [CrossRef] [PubMed]
- Nowell, V.J.; Kropinski, A.M.; Songer, J.G.; MacInnes, J.I.; Parreira, V.R.; Prescott, J.F. Genome sequencing and analysis of a type A Clostridium perfringens isolate from a case of bovine clostridial abomasitis. PLoS ONE 2012, 7, e32271. [Google Scholar] [CrossRef]
- Abdel-Glil, M.Y.; Thomas, P.; Linde, J.; Busch, A.; Wieler, L.H.; Neubauer, H.; Seyboldt, C. Comparative in silico genome analysis of Clostridium perfringens unravels stable phylogroups with different genome characteristics and pathogenic potential. Sci. Rep. 2021, 11, 6756. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Strain | ATCC13124 | Str.13 | CPN 17a | JP838 | SM101 | FORC_003 | F262 | EHE-NE18 |
|---|---|---|---|---|---|---|---|---|
| SOURCE | Unknown | Japan | Finland | America | America | Korea | Canada | Australia |
| HOSTS | Human | NA | Cattle | Dog | NA | NA | Cattle | Chicken |
| TYPE OF SAMPLE | Gas gangrene tissue | Soil | Feces | Feces | Food | Aquarium water source | Contents of the abomasum | Intestinal contents |
| GENOTYPE | A | A | NA | A | A | NA | A | NA |
| GI_ID | seq_ID | Start | End | GI_Length |
|---|---|---|---|---|
| GI1 | assembly_1 | 1,628,448 | 1,637,836 | 9389 |
| GI2 | assembly_1 | 2,129,345 | 2,140,974 | 11,630 |
| GI3 | assembly_1 | 2,854,714 | 2,861,402 | 6689 |
| GI4 | assembly_1 | 3,283,647 | 3,295,496 | 11,850 |
| Sequence | Pseudogene Size (bp) | Pseudogene Number | Average Length (bp) | Percentage |
|---|---|---|---|---|
| ctg-1 | 36,695 | 72 | 509.65 | 1.09% |
| VFclass | Virulence Factors | Related Genes | Results |
|---|---|---|---|
| Adherence | Cna | cna | + |
| Fibronectin-binding protein | fbpA/fbp68 | + | |
| GroEL | groEL | + | |
| Toxin | Alpha-clostripain | cloSI | + |
| Alpha-toxin | plc | + | |
| Beta2 toxin | cpb2 | + | |
| C. perfringens enterotoxin (CPE) | cpe | − | |
| Hemolysin | Hemolysin | + | |
| Kappa-toxin (collagenase) | colA | + | |
| Mu-toxin (neuraminidase) | nagH | + | |
| nagI | + | ||
| nagJ | + | ||
| nagK | − | ||
| nagL | + | ||
| Perfringolysin O (theta-toxin/PFO)/botulinolysin/tetanolysin O | pfoA | + | |
| Sialidase | nanH | + | |
| nanI | + | ||
| nanJ | + | ||
| Regulation | VirR/VirS two-component system | virR | + |
| virS | + |
| Bacterial Strain | Genome Size | GC Ratio (%) | Number of Genes | CDS Region | tRNA | rRNA | Pseudogenes | Crispr Sequence |
|---|---|---|---|---|---|---|---|---|
| CX1-4 | 3,355,389 | 28.36 | 3193 | 3002 | 94 | 30 | 72 | 0 |
| ATCC13124 | 3,256,683 | 28.38 | 2976 | 2855 | 93 | 24 | 27 | 0 |
| Str.13 | 3,085,740 | 28.51 | 2813 | 2683 | 96 | 30 | 68 | 0 |
| SM101 | 2,960,088 | 28.23 | 2694 | 2566 | 94 | 30 | 116 | 1 |
| JP838 | 4,070,811 | 27.94 | 3202 | 3014 | 92 | 30 | 65 | 0 |
| CPN 17a | 3,266,195 | 28.39 | 2976 | 2848 | 94 | 30 | 22 | 2 |
| FORC_003 | 3,338,532 | 28.38 | 3093 | 2962 | 97 | 30 | 36 | 1 |
| F262 | 3,333,039 | 28.04 | 3224 | 3163 | 76 | 30 | NA | NA |
| EHE-NE18 | 3,463,721 | 28.27 | 3409 | 3278 | 97 | 30 | 60 | 0 |
| Virulence Factors | Related Genes | CX1-4 | ATCC 13124 | Str.13 | SM101 | |
|---|---|---|---|---|---|---|
| Adherence | Cna | cna | + | − | − | − |
| Fibronectin-binding protein | fbpA/fbp68 | + | + | + | + | |
| GroEL | groEL | + | + | + | − | |
| Toxin | Alpha-clostripain | cloSI | + | + | + | + |
| Alpha-toxin | plc | + | + | + | + | |
| Beta2 toxin | cpb2 | + | − | + | − | |
| C.perfringens enterotoxin (CPE) | cpe | − | − | − | + | |
| Hemolysin | Hemolysin | + | + | + | + | |
| Kappa-toxin (collagenase) | colA | + | + | + | + | |
| Mu-toxin (neuraminidase) | nagH | + | + | + | − | |
| nagI | + | + | + | − | ||
| nagJ | + | + | + | − | ||
| nagK | − | + | + | − | ||
| nagL | + | − | + | − | ||
| Perfringolysin O (theta-toxin/PFO)/botulinolysin/tetanolysin O | pfoA | + | + | + | − | |
| Sialidase | nanH | + | + | − | + | |
| nanI | + | + | + | − | ||
| nanJ | + | + | + | − | ||
| Regulation | VirR/VirS two-component system | virR | + | + | + | + |
| virS | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wang, X.; Li, S.; Fu, Y.; Li, Y.; Nawaz, S.; Chen, J.; Yang, G.; Li, J.; Shi, D. Isolation of a Virulent Clostridium perfringens Strain from Elaphurus davidianus and Characterization by Whole-Genome Sequence Analysis. Curr. Issues Mol. Biol. 2024, 46, 7169-7186. https://doi.org/10.3390/cimb46070427
Zhang Z, Wang X, Li S, Fu Y, Li Y, Nawaz S, Chen J, Yang G, Li J, Shi D. Isolation of a Virulent Clostridium perfringens Strain from Elaphurus davidianus and Characterization by Whole-Genome Sequence Analysis. Current Issues in Molecular Biology. 2024; 46(7):7169-7186. https://doi.org/10.3390/cimb46070427
Chicago/Turabian StyleZhang, Zhao, Xiao Wang, Siyuan Li, Yuhang Fu, Yan Li, Shah Nawaz, Jing Chen, Guoxiang Yang, Jiakui Li, and Daoliang Shi. 2024. "Isolation of a Virulent Clostridium perfringens Strain from Elaphurus davidianus and Characterization by Whole-Genome Sequence Analysis" Current Issues in Molecular Biology 46, no. 7: 7169-7186. https://doi.org/10.3390/cimb46070427
APA StyleZhang, Z., Wang, X., Li, S., Fu, Y., Li, Y., Nawaz, S., Chen, J., Yang, G., Li, J., & Shi, D. (2024). Isolation of a Virulent Clostridium perfringens Strain from Elaphurus davidianus and Characterization by Whole-Genome Sequence Analysis. Current Issues in Molecular Biology, 46(7), 7169-7186. https://doi.org/10.3390/cimb46070427